
Expression and Localization of a New Parvovirus-Derived Protein in the Guinea Pig
 , ,
, , 
 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Plasmids and Cloning
2.3. Generation of Stable Cell Lines
2.4. Antibodies Generation
2.5. Western Blot Assays
2.6. Immunofluorescence
2.7. Proximity Ligation Assay
3. Results
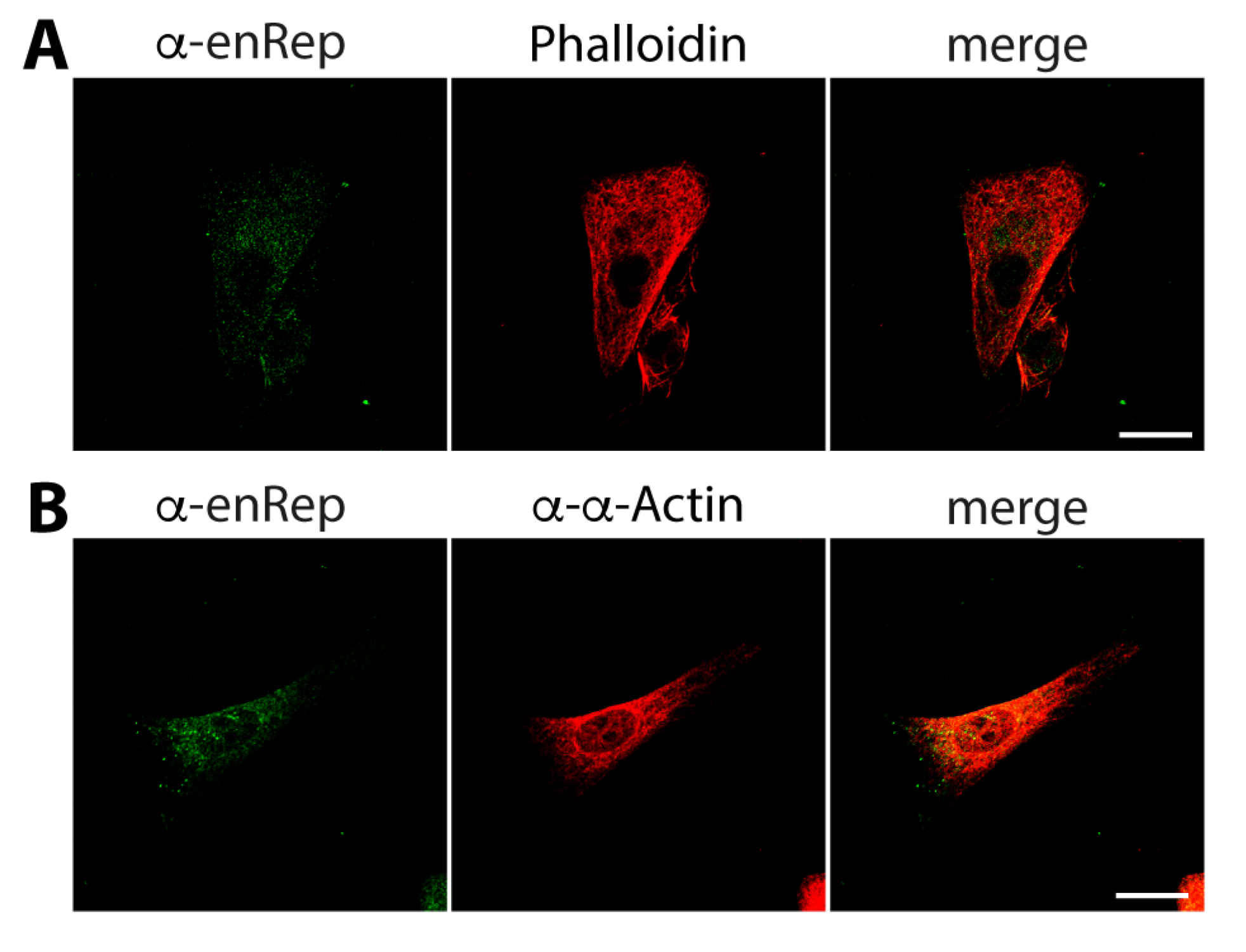
3.1. The EPV-Derived Protein enRep-M9l Is Expressed in Guinea Pig Cells and Tissues
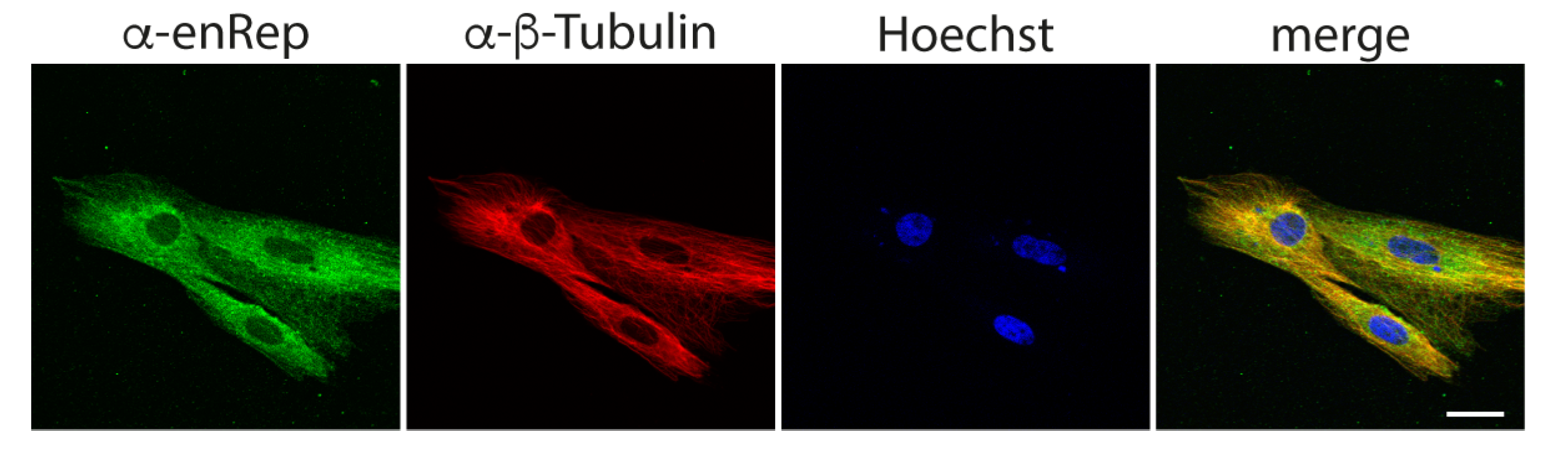
3.2. The Protein enRep-M9l Is in Close Proximity to Microtubules
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kapoor, A.; Simmonds, P.; Lipkin, W.I. Discovery and characterization of mammalian endogenous parvoviruses. J. Virol. 2010, 84, 12628–12635. [Google Scholar] [CrossRef]
- Belyi, V.A.; Levine, A.J.; Skalka, A.M. Sequences from ancestral single-stranded DNA viruses in vertebrate genomes: The parvoviridae and circoviridae are more than 40 to 50 million years old. J. Virol. 2010, 84, 12458–12462. [Google Scholar] [CrossRef] [PubMed]
- Katzourakis, A.; Gifford, R.J. Endogenous viral elements in animal genomes. PLoS Genet. 2010, 6, e1001191. [Google Scholar] [CrossRef]
- Horie, M.; Tomonaga, K. Non-retroviral fossils in vertebrate genomes. Viruses 2011, 3, 1836–1848. [Google Scholar] [CrossRef]
- Feschotte, C.; Gilbert, C. Endogenous viruses: Insights into viral evolution and impact on host biology. Nat. Rev. Genet. 2012, 13, 283–296. [Google Scholar] [CrossRef]
- Dennis, T.P.W.; Flynn, P.J.; de Souza, W.M.; Singer, J.B.; Moreau, C.S.; Wilson, S.J.; Gifford, R.J. Insights into Circovirus Host Range from the Genomic Fossil Record. J. Virol. 2018, 92, e00145-18. [Google Scholar] [CrossRef]
- Campbell, M.A.; Loncar, S.; Kotin, R.M.; Gifford, R.J. Comparative analysis reveals the long-term coevolutionary history of parvoviruses and vertebrates. PLoS Biol. 2022, 20, e3001867. [Google Scholar] [CrossRef] [PubMed]
- Holmes, E.C. The evolution of endogenous viral elements. Cell Host Microbe 2011, 10, 368–377. [Google Scholar] [CrossRef]
- Horie, M.; Honda, T.; Suzuki, Y.; Kobayashi, Y.; Daito, T.; Oshida, T.; Ikuta, K.; Jern, P.; Gojobori, T.; Coffin, J.M.; et al. Endogenous non-retroviral RNA virus elements in mammalian genomes. Nature 2010, 463, 84–87. [Google Scholar] [CrossRef]
- Shimizu, A.; Nakatani, Y.; Nakamura, T.; Jinno-Oue, A.; Ishikawa, O.; Boeke, J.D.; Takeuchi, Y.; Hoshino, H. Characterisation of cytoplasmic DNA complementary to non-retroviral RNA viruses in human cells. Sci. Rep. 2014, 4, 5074. [Google Scholar] [CrossRef]
- Dewannieux, M.; Heidmann, T. Endogenous retroviruses: Acquisition, amplification and taming of genome invaders. Curr. Opin. Virol. 2013, 3, 646–656. [Google Scholar] [CrossRef] [PubMed]
- Frank, J.A.; Feschotte, C. Co-option of endogenous viral sequences for host cell function. Curr. Opin. Virol. 2017, 25, 81–89. [Google Scholar] [CrossRef]
- Gautam, P.; Yu, T.; Loh, Y.H. Regulation of ERVs in pluripotent stem cells and reprogramming. Curr. Opin. Genet. Dev. 2017, 46, 194–201. [Google Scholar] [CrossRef]
- Fujino, K.; Horie, M.; Honda, T.; Merriman, D.K.; Tomonaga, K. Inhibition of Borna disease virus replication by an endogenous bornavirus-like element in the ground squirrel genome. Proc. Natl. Acad. Sci. USA 2014, 111, 13175–13180. [Google Scholar] [CrossRef] [PubMed]
- Horie, M. The biological significance of bornavirus-derived genes in mammals. Curr. Opin. Virol. 2017, 25, 1–6. [Google Scholar] [CrossRef]
- Fujino, K.; Horie, M.; Kojima, S.; Shimizu, S.; Nabekura, A.; Kobayashi, H.; Makino, A.; Honda, T.; Tomonaga, K. A Human Endogenous Bornavirus-Like Nucleoprotein Encodes a Mitochondrial Protein Associated with Cell Viability. J. Virol. 2021, 95, e0203020. [Google Scholar] [CrossRef]
- Bravo, A.; Fernández-García, L.; Ibarra-Karmy, R.; Mardones, G.A.; Mercado, L.; Bustos, F.J.; Gifford, R.J.; Arriagada, G. Antiviral Activity of an Endogenous Parvoviral Element. Viruses 2023, 15, 1420. [Google Scholar] [CrossRef]
- Horie, M.; Tomonaga, K. Paleovirology of bornaviruses: What can be learned from molecular fossils of bornaviruses. Virus Res. 2019, 262, 2–9. [Google Scholar] [CrossRef]
- Blanco-Melo, D.; Gifford, R.J.; Bieniasz, P.D. Co-option of an endogenous retrovirus envelope for host defense in hominid ancestors. Elife 2017, 6, e22519. [Google Scholar] [CrossRef]
- Durnaoglu, S.; Lee, S.K.; Ahnn, J. Syncytin, envelope protein of human endogenous retrovirus (HERV): No longer ‘fossil’ in human genome. Anim. Cells Syst. 2021, 25, 358–368. [Google Scholar] [CrossRef]
- Ogawa, H.; Honda, T. Viral Sequences Are Repurposed for Controlling Antiviral Responses as Non-Retroviral Endogenous Viral Elements. Acta Med. Okayama 2022, 76, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Sofuku, K.; Parrish, N.F.; Honda, T.; Tomonaga, K. Transcription Profiling Demonstrates Epigenetic Control of Non-retroviral RNA Virus-Derived Elements in the Human Genome. Cell Rep. 2015, 12, 1548–1554. [Google Scholar] [CrossRef]
- Valencia-Herrera, I.; Faunes, F.; Cena-Ahumada, E.; Ibarra-Karmy, R.; Gifford, R.J.; Arriagada, G. Molecular Properties and Evolutionary Origins of a Parvovirus-Derived Myosin Fusion Gene in Guinea Pigs. J. Virol. 2019, 93, e00404-19. [Google Scholar] [CrossRef]
- Arriagada, G.; Gifford, R.J. Parvovirus-derived endogenous viral elements in two South American rodent genomes. J. Virol. 2014, 88, 12158–12162. [Google Scholar] [CrossRef]
- Smith, R.H.; Hallwirth, C.V.; Westerman, M.; Hetherington, N.A.; Tseng, Y.S.; Cecchini, S.; Virag, T.; Ziegler, M.L.; Rogozin, I.B.; Koonin, E.V.; et al. Germline viral “fossils” guide in silico reconstruction of a mid-Cenozoic era marsupial adeno-associated virus. Sci. Rep. 2016, 6, 28965. [Google Scholar] [CrossRef] [PubMed]
- Hildebrandt, E.; Penzes, J.J.; Gifford, R.J.; Agbandje-Mckenna, M.; Kotin, R.M. Evolution of dependoparvoviruses across geological timescales-implications for design of AAV-based gene therapy vectors. Virus Evol. 2020, 6, veaa043. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Fu, Y.; Xie, J.; Cheng, J.; Ghabrial, S.A.; Li, G.; Peng, Y.; Yi, X.; Jiang, D. Widespread endogenization of densoviruses and parvoviruses in animal and human genomes. J. Virol. 2011, 85, 9863–9876. [Google Scholar] [CrossRef]
- Pénzes, J.J.; de Souza, W.M.; Agbandje-McKenna, M.; Gifford, R.J. An Ancient Lineage of Highly Divergent Parvoviruses Infects both Vertebrate and Invertebrate Hosts. Viruses 2019, 11, 525. [Google Scholar] [CrossRef]
- Cotmore, S.F.; Tattersall, P. Parvoviruses: Small Does Not Mean Simple. Annu. Rev. Virol. 2014, 1, 517–537. [Google Scholar] [CrossRef]
- Yang, C.C.; Xiao, X.; Zhu, X.; Ansardi, D.C.; Epstein, N.D.; Frey, M.R.; Matera, A.G.; Samulski, R.J. Cellular recombination pathways and viral terminal repeat hairpin structures are sufficient for adeno-associated virus integration in vivo and in vitro. J. Virol. 1997, 71, 9231–9247. [Google Scholar] [CrossRef]
- Daya, S.; Cortez, N.; Berns, K.I. Adeno-associated virus site-specific integration is mediated by proteins of the nonhomologous end-joining pathway. J. Virol. 2009, 83, 11655–11664. [Google Scholar] [CrossRef]
- Deyle, D.R.; Russell, D.W. Adeno-associated virus vector integration. Curr. Opin. Mol. Ther. 2009, 11, 442–447. [Google Scholar] [PubMed]
- Kobayashi, Y.; Shimazu, T.; Murata, K.; Itou, T.; Suzuki, Y. An endogenous adeno-associated virus element in elephants. Virus Res. 2019, 262, 10–14. [Google Scholar] [CrossRef] [PubMed]
- Callaway, H.M.; Subramanian, S.; Urbina, C.A.; Barnard, K.N.; Dick, R.A.; Bator, C.M.; Hafenstein, S.L.; Gifford, R.J.; Parrish, C.R. Examination and Reconstruction of Three Ancient Endogenous Parvovirus Capsid Protein Gene Remnants Found in Rodent Genomes. J. Virol. 2019, 93, e01542-18. [Google Scholar] [CrossRef]
- Mermall, V.; Post, P.L.; Mooseker, M.S. Unconventional myosins in cell movement, membrane traffic, and signal transduction. Science 1998, 279, 527–533. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]
- Zarate-Perez, F.; Mansilla-Soto, J.; Bardelli, M.; Burgner, J.W., 2nd; Villamil-Jarauta, M.; Kekilli, D.; Samso, M.; Linden, R.M.; Escalante, C.R. Oligomeric properties of adeno-associated virus Rep68 reflect its multifunctionality. J. Virol. 2013, 87, 1232–1241. [Google Scholar] [CrossRef]
- Bardelli, M.; Zárate-Pérez, F.; Agúndez, L.; Linden, R.M.; Escalante, C.R.; Henckaerts, E. Identification of a Functionally Relevant Adeno-Associated Virus Rep68 Oligomeric Interface. J. Virol. 2016, 90, 6612–6624. [Google Scholar] [CrossRef]
- Xie, Q.; Wang, J.; Gu, C.; Wu, J.; Liu, W. Structure and function of the parvoviral NS1 protein: A review. Virus Genes 2023, 59, 195–203. [Google Scholar] [CrossRef]
- Song, Y.; Nguyen, X.N.; Kumar, A.; da Silva, C.; Picard, L.; Etienne, L.; Cimarelli, A. Trim69 is a microtubule regulator that acts as a pantropic viral inhibitor. Proc. Natl. Acad. Sci. USA 2022, 119, e2211467119. [Google Scholar] [CrossRef]
- Vadon, C.; Magiera, M.M.; Cimarelli, A. TRIM Proteins and Antiviral Microtubule Reorganization: A Novel Component in Innate Immune Responses? Viruses 2024, 16, 1328. [Google Scholar] [CrossRef] [PubMed]
- Hanley, P.J.; Xu, Y.; Kronlage, M.; Grobe, K.; Schon, P.; Song, J.; Sorokin, L.; Schwab, A.; Bahler, M. Motorized RhoGAP myosin IXb (Myo9b) controls cell shape and motility. Proc. Natl. Acad. Sci. USA 2010, 107, 12145–12150. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Lu, Q.; Zhang, M. Structural Basis of Cargo Recognition by Unconventional Myosins in Cellular Trafficking. Traffic 2016, 17, 822–838. [Google Scholar] [CrossRef] [PubMed]
- Bernards, A.; Settleman, J. GAP control: Regulating the regulators of small GTPases. Trends Cell Biol. 2004, 14, 377–385. [Google Scholar] [CrossRef]
- Guan, G.; Cannon, R.D.; Coates, D.E.; Mei, L. Effect of the Rho-Kinase/ROCK Signaling Pathway on Cytoskeleton Components. Genes 2023, 14, 272. [Google Scholar] [CrossRef]
- Chang, Y.C.; Nalbant, P.; Birkenfeld, J.; Chang, Z.F.; Bokoch, G.M. GEF-H1 couples nocodazole-induced microtubule disassembly to cell contractility via RhoA. Mol. Biol. Cell 2008, 19, 2147–2153. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Osega, C.E.; Bustos, F.J.; Bronfman, F.C.; Gifford, R.J.; Arriagada, G. Expression and Localization of a New Parvovirus-Derived Protein in the Guinea Pig. Viruses 2025, 17, 893. https://doi.org/10.3390/v17070893
Osega CE, Bustos FJ, Bronfman FC, Gifford RJ, Arriagada G. Expression and Localization of a New Parvovirus-Derived Protein in the Guinea Pig. Viruses. 2025; 17(7):893. https://doi.org/10.3390/v17070893
Chicago/Turabian StyleOsega, Camila E., Fernando J. Bustos, Francisca C. Bronfman, Robert J. Gifford, and Gloria Arriagada. 2025. "Expression and Localization of a New Parvovirus-Derived Protein in the Guinea Pig" Viruses 17, no. 7: 893. https://doi.org/10.3390/v17070893
APA StyleOsega, C. E., Bustos, F. J., Bronfman, F. C., Gifford, R. J., & Arriagada, G. (2025). Expression and Localization of a New Parvovirus-Derived Protein in the Guinea Pig. Viruses, 17(7), 893. https://doi.org/10.3390/v17070893