

Rotavirus alphagastroenteritidis: Circulating Strains After the Introduction of the Rotavirus Vaccine (Rotarix®) in Luanda Province of Angola

 , , ,
, , ,  ,
,  , ,
, ,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Sample Collection and R. alphagastroenteritidis Detection
2.3. Genotyping of R. alphagastroenteritidis Strains
2.4. Sequencing of R. alphagastroenteritidis Strains
2.5. Phylogenetic Analysis
2.6. Statistical Analysis
2.7. Ethical Considerations
3. Results
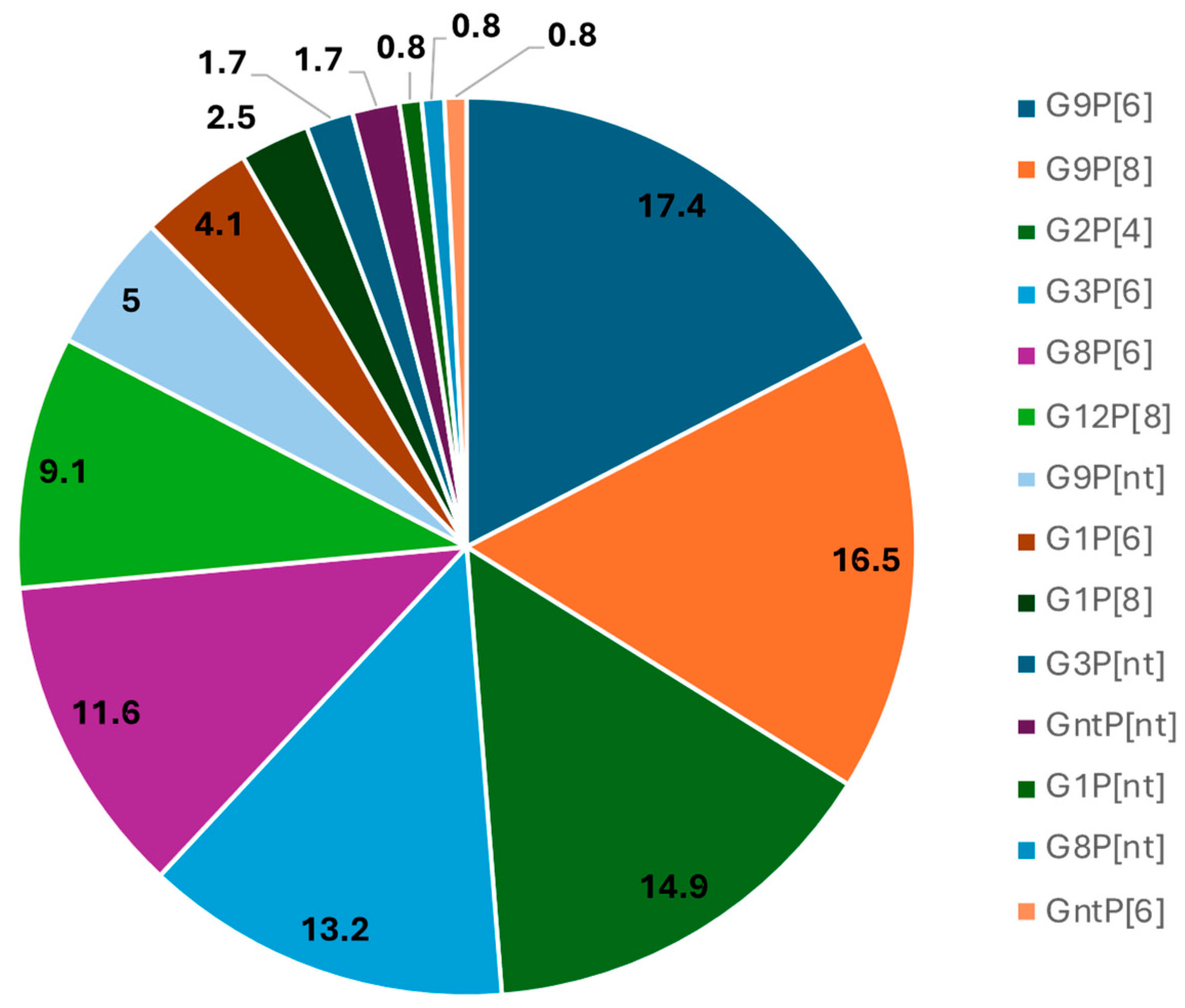
3.1. High Diversity of R. alphagastroenteritidis Strains Detected in Children Hospitalized with AGE in Luanda Province Public Hospitals
3.2. Genotype Diversity, Age, and Severity of Disease
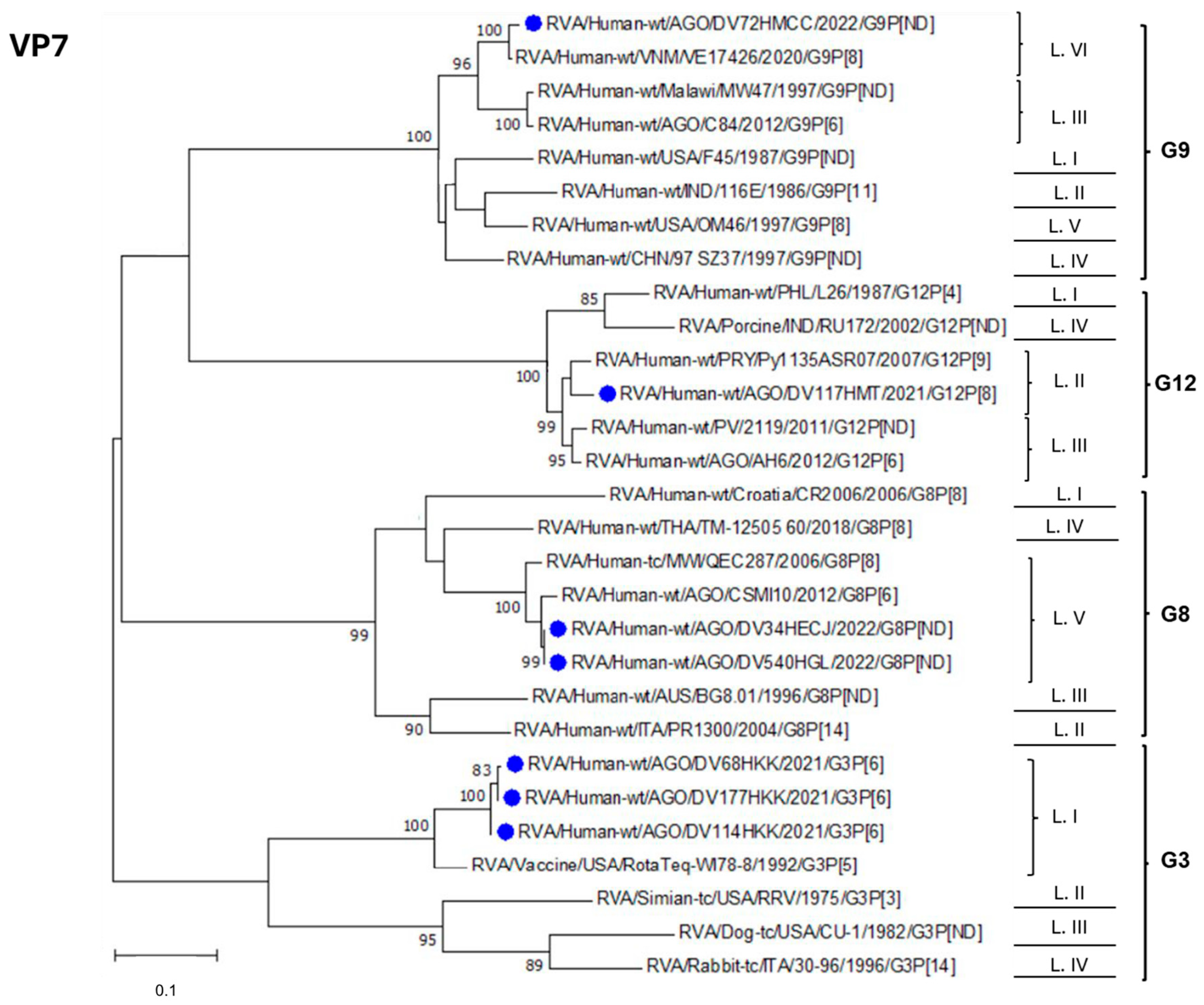
3.3. Phylogenetic Analysis of R. alphagastroenteritidis Strains Identified in Luanda Province Post-Vaccine Introduction
3.4. Comparison of the Deduced Antigenic Region of VP4 (VP8 *) and VP7
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Crawford, S.E.; Ramani, S.; Tate, J.E.; Parashar, U.D.; Svensson, L.; Hagbom, M.; Franco, M.A.; Greenberg, H.B.; O’Ryan, M.; Kang, G.; et al. Rotavirus infection. Nat. Rev. Dis. Primers 2017, 3, 17083. [Google Scholar] [CrossRef] [PubMed]
- Jampanil, N.; Kumthip, K.; Maneekarn, N.; Khamrin, P. Genetic Diversity of Rotaviruses Circulating in Pediatric Patients and Domestic Animals in Thailand. Trop. Med. Infect. Dis. 2023, 8, 347. [Google Scholar] [CrossRef]
- Troeger, C.; Khalil, I.A.; Rao, P.C.; Cao, S.; Blacker, B.F.; Ahmed, T.; Armah, G.; Bines, J.E.; Brewer, T.G.; Colombara, D.V.; et al. Rotavirus Vaccination and the Global Burden of Rotavirus Diarrhea Among Children Younger Than 5 Years. JAMA Pediatr. 2018, 172, 958–965. [Google Scholar] [CrossRef]
- Tate, J.E.; Burton, A.H.; Boschi-Pinto, C.; Parashar, U.D. Global, Regional, and National Estimates of Rotavirus Mortality in Children <5 Years of Age, 2000–2013. Clin. Infect. Dis. 2016, 62 (Suppl. S2), S96–S105. [Google Scholar] [CrossRef] [PubMed]
- Sadiq, A.; Khan, J. Rotavirus in developing countries: Molecular diversity, epidemiological insights, and strategies for effective vaccination. Front. Microbiol. 2023, 14, 1297269. [Google Scholar] [CrossRef] [PubMed]
- Burnett, E.; Parashar, U.D.; Tate, J.E. Real-world effectiveness of rotavirus vaccines, 2006–2019: A literature review and meta-analysis. Lancet Glob. Health 2020, 8, e1195–e1202. [Google Scholar] [CrossRef]
- Desselberger, U. Differences of Rotavirus Vaccine Effectiveness by Country: Likely Causes and Contributing Factors. Pathogens 2017, 6, 65. [Google Scholar] [CrossRef]
- Parker, E.P.; Ramani, S.; Lopman, B.A.; Church, J.A.; Iturriza-Gomara, M.; Prendergast, A.J.; Grassly, N.C. Causes of impaired oral vaccine efficacy in developing countries. Future Microbiol. 2018, 13, 97–118. [Google Scholar] [CrossRef]
- Desselberger, U. Rotaviruses. Virus Res. 2014, 190, 75–96. [Google Scholar] [CrossRef]
- Matthijnssens, J.; Otto, P.H.; Ciarlet, M.; Desselberger, U.; Van Ranst, M.; Johne, R. VP6-sequence-based cutoff values as a criterion for rotavirus species demarcation. Arch. Virol. 2012, 157, 1177–1182. [Google Scholar] [CrossRef]
- Matthijnssens, J.; Ciarlet, M.; McDonald, S.M.; Attoui, H.; Banyai, K.; Brister, J.R.; Buesa, J.; Esona, M.D.; Estes, M.K.; Gentsch, J.R.; et al. Uniformity of rotavirus strain nomenclature proposed by the Rotavirus Classification Working Group (RCWG). Arch. Virol. 2011, 156, 1397–1413. [Google Scholar] [CrossRef] [PubMed]
- RCWG. 2023. Available online: https://rega.kuleuven.be/cev/viralmetagenomics/virus-classification/rcwg (accessed on 15 March 2025).
- Sadiq, A.; Bostan, N.; Jadoon, K.; Aziz, A. Effect of rotavirus genetic diversity on vaccine impact. Rev. Med. Virol. 2022, 32, e2259. [Google Scholar] [CrossRef] [PubMed]
- Doro, R.; Farkas, S.L.; Martella, V.; Banyai, K. Zoonotic transmission of rotavirus: Surveillance and control. Expert Rev. Anti Infect. Ther. 2015, 13, 1337–1350. [Google Scholar] [CrossRef] [PubMed]
- Bányai, K.; Pitzer, V.E. Chapter 2.10—Molecular Epidemiology and Evolution of Rotaviruses. In Viral Gastroenteritis; Svensson, L., Desselberger, U., Greenberg, H.B., Estes, M.K., Eds.; Academic Press: Boston, MA, USA, 2016; pp. 279–299. [Google Scholar]
- Bányai, K.; Gentsch, J. Special issue on ‘Genetic diversity and evolution of rotavirus strains: Possible impact of global immunization programs’. Infect. Genet. Evol. 2014, 28, 375–376. [Google Scholar] [CrossRef]
- Vita, D.; Lemos, M.; Neto, Z.; Evans, M.; Francisco, N.M.; Fortes, F.; Fernandes, E.; Cunha, C.; Istrate, C. High Detection Rate of Rotavirus Infection Among Children Admitted with Acute Gastroenteritis to Six Public Hospitals in Luanda Province After the Introduction of Rotarix((R)) Vaccine: A Cross-Sectional Study. Viruses 2024, 16, 1949. [Google Scholar] [CrossRef]
- Gentsch, J.R.; Glass, R.I.; Woods, P.; Gouvea, V.; Gorziglia, M.; Flores, J.; Das, B.K.; Bhan, M.K. Identification of group A rotavirus gene 4 types by polymerase chain reaction. J. Clin. Microbiol. 1992, 30, 1365–1373. [Google Scholar] [CrossRef]
- Iturriza-Gomara, M.; Kang, G.; Gray, J. Rotavirus genotyping: Keeping up with an evolving population of human rotaviruses. J. Clin. Virol. 2004, 31, 259–265. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignement editor and analysis program for windows 95/98/NT. Nucleic Acids Symp. 1999, 41, 95–98. [Google Scholar]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef]
- Nicholas, K.B.; Nicholas, H.B.; Deerfield, D.W. GeneDoc: Analysis and visualization of genetic variation. EMBnet News 1997, 4, 1–4. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. Evolutionary trees from DNA sequences: A maximum likelihood approach. J. Mol. Evol. 1981, 17, 368–376. [Google Scholar] [CrossRef]
- Aoki, S.T.; Settembre, E.C.; Trask, S.D.; Greenberg, H.B.; Harrison, S.C.; Dormitzer, P.R. Structure of Rotavirus Outer-Layer Protein VP7 Bound with a Neutralizing Fab. Science 2009, 324, 1444–1447. [Google Scholar] [CrossRef] [PubMed]
- Dormitzer, P.R.; Nason, E.B.; Prasad, B.V.; Harrison, S.C. Structural rearrangements in the membrane penetration protein of a non-enveloped virus. Nature 2004, 430, 1053–1058. [Google Scholar] [CrossRef] [PubMed]
- Dormitzer, P.R.; Sun, Z.Y.; Wagner, G.; Harrison, S.C. The rhesus rotavirus VP4 sialic acid binding domain has a galectin fold with a novel carbohydrate binding site. EMBO J. 2002, 21, 885–897. [Google Scholar] [CrossRef]
- Zeller, M.; Patton, J.T.; Heylen, E.; Coster, S.D.; Ciarlet, M.; Ranst, M.V.; Matthijnssens, J. Genetic Analyses Reveal Differences in the VP7 and VP4 Antigenic Epitopes between Human Rotaviruses Circulating in Belgium and Rotaviruses in Rotarix and RotaTeq. J. Clin. Microbiol. 2012, 50, 966–976. [Google Scholar] [CrossRef]
- Esteves, A.; Nordgren, J.; Pereira, J.; Fortes, F.; Dimbu, R.; Saraiva, N.; Mendes, C.; Istrate, C. Molecular epidemiology of rotavirus in four provinces of Angola before vaccine introduction. J. Med. Virol. 2016, 88, 1511–1520. [Google Scholar] [CrossRef] [PubMed]
- Omatola, C.A.; Ogunsakin, R.E.; Olaniran, A.O. Prevalence, Pattern and Genetic Diversity of Rotaviruses among Children under 5 Years of Age with Acute Gastroenteritis in South Africa: A Systematic Review and Meta-Analysis. Viruses 2021, 13, 1905. [Google Scholar] [CrossRef]
- Bawa, F.K.; Mutocheluh, M.; Dassah, S.D.; Ansah, P.; Oduro, A.R. Genetic diversity of rotavirus infection among young children with diarrhoea in the Kassena-Nankana Districts of Northern Ghana: A seasonal cross-sectional survey. Pan Afr. Med. J. 2023, 44, 148. [Google Scholar] [CrossRef]
- Munlela, B.; Joao, E.D.; Strydom, A.; Bauhofer, A.F.L.; Chissaque, A.; Chilaule, J.J.; Mauricio, I.L.; Donato, C.M.; O’Neill, H.G.; de Deus, N. Whole-Genome Characterization of Rotavirus G9P[6] and G9P[4] Strains That Emerged after Rotavirus Vaccine Introduction in Mozambique. Viruses 2024, 16, 1140. [Google Scholar] [CrossRef]
- Gasparinho, C.; Piedade, J.; Mirante, M.C.; Mendes, C.; Mayer, C.; Vaz Nery, S.; Brito, M.; Istrate, C. Characterization of rotavirus infection in children with acute gastroenteritis in Bengo province, Northwestern Angola, prior to vaccine introduction. PLoS ONE 2017, 12, e0176046. [Google Scholar] [CrossRef] [PubMed]
- Bonura, F.; Mangiaracina, L.; Filizzolo, C.; Bonura, C.; Martella, V.; Ciarlet, M.; Giammanco, G.M.; De Grazia, S. Impact of Vaccination on Rotavirus Genotype Diversity: A Nearly Two-Decade-Long Epidemiological Study before and after Rotavirus Vaccine Introduction in Sicily, Italy. Pathogens 2022, 11, 424. [Google Scholar] [CrossRef] [PubMed]
- Joao, E.D.; Munlela, B.; Chissaque, A.; Chilaule, J.; Langa, J.; Augusto, O.; Boene, S.S.; Anapakala, E.; Sambo, J.; Guimaraes, E.; et al. Molecular Epidemiology of Rotavirus A Strains Pre- and Post-Vaccine (Rotarix®) Introduction in Mozambique, 2012–2019: Emergence of Genotypes G3P[4] and G3P[8]. Pathogens 2020, 9, 671. [Google Scholar] [CrossRef] [PubMed]
- Mhango, C.; Banda, A.; Chinyama, E.; Mandolo, J.J.; Kumwenda, O.; Malamba-Banda, C.; Barnes, K.G.; Kumwenda, B.; Jambo, K.C.; Donato, C.M.; et al. Comparative whole genome analysis reveals re-emergence of human Wa-like and DS-1-like G3 rotaviruses after Rotarix vaccine introduction in Malawi. Virus Evol. 2023, 9, vead030. [Google Scholar] [CrossRef]
- Seheri, L.M.; Magagula, N.B.; Peenze, I.; Rakau, K.; Ndadza, A.; Mwenda, J.M.; Weldegebriel, G.; Steele, A.D.; Mphahlele, M.J. Rotavirus strain diversity in Eastern and Southern African countries before and after vaccine introduction. Vaccine 2018, 36, 7222–7230. [Google Scholar] [CrossRef]
- Ramachandran, M.; Das, B.K.; Vij, A.; Kumar, R.; Bhambal, S.S.; Kesari, N.; Rawat, H.; Bahl, L.; Thakur, S.; Woods, P.A.; et al. Unusual diversity of human rotavirus G and P genotypes in India. J. Clin. Microbiol. 1996, 34, 436–439. [Google Scholar] [CrossRef]
- Page, N.A.; Seheri, L.M.; Groome, M.J.; Moyes, J.; Walaza, S.; Mphahlele, J.; Kahn, K.; Kapongo, C.N.; Zar, H.J.; Tempia, S.; et al. Temporal association of rotavirus vaccination and genotype circulation in South Africa: Observations from 2002 to 2014. Vaccine 2018, 36, 7231–7237. [Google Scholar] [CrossRef]
- Gurgel, R.Q.; Cuevas, L.E.; Vieira, S.C.; Barros, V.C.; Fontes, P.B.; Salustino, E.F.; Nakagomi, O.; Nakagomi, T.; Dove, W.; Cunliffe, N.; et al. Predominance of rotavirus P[4]G2 in a vaccinated population, Brazil. Emerg. Infect. Dis. 2007, 13, 1571–1573. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| G-type | N | % |
| G1 | 9 | 7.4 |
| G2 | 18 | 14.9 |
| G3 | 18 | 14.9 |
| G8 | 15 | 12.4 |
| G9 | 46 | 38.8 |
| G12 | 11 | 9.1 |
| Gnt a | 3 | 2.5 |
| Total | 121 | 100 |
| P-type | N | % |
| P4 | 18 | 14.8 |
| P6 | 57 | 47.1 |
| P8 | 34 | 28.0 |
| Pnt b | 12 | 9.9 |
| Total | 121 | 100 |
| Genotype Combination | Cacuaco n (%) | Luanda n (%) | Zango n (%) | Cajueiros n (%) | Talatona n (%) | K. Kiaxi n (%) | Total n (%) |
|---|---|---|---|---|---|---|---|
| G1P[6] | 0 (0.0) | 2 (6.9) | 3 (9.7) | 0 (0.0) | 0 (00.0) | 0 (0.0) | 5 (4.1) |
| G1P[8] | 0 (0.0) | 0 (0.0) | 0 (0.0) | 0 (0.0) | 3 (17.6) | 0 (0.0) | 3 (2.5) |
| G1Pnt * | 0 (0.0) | 1 (3.4) | 0 (0.0) | 0 (0.0) | 0 (0.0) | 0 (0.0) | 1 (0.8) |
| G2P[4] | 5 (31.2) | 3 (10.3) | 9 (29.0) | 0 (0.0) | 1 (5.9) | 0 (0.0) | 18 (14.9) |
| G3P[6] | 0 (0.0) | 2 (6.9) | 1 (3.2) | 1 (10) | 2 (11.8) | 10 (55.6) | 16 (13.2) |
| G3Pnt * | 0 (0.0) | 0 (0.0) | 1 (3.2) | 0 (0.0) | 0 (0.0) | 1 (5.6) | 2 (1.6) |
| G8P[6] | 1 (6.2) | 3 (10.3) | 3 (9.7) | 4 (40.0) | 2 (11.8) | 1 (5.6) | 14 (11.5) |
| G8Pnt * | 0 (0.0) | 1 (3.4) | 0 (0.0) | 0 (0.0) | 0 (0.0) | 0 (0.0) | 1 (1.7) |
| G9P[6] | 3 (18.8) | 10(34.5) | 5 (16.1) | 1 (10) | 1 (5.9) | 1 (5.6) | 21 (17.3) |
| G9P[8] | 6 (37.5) | 3(10.3) | 7 (22.6) | 0 (0.0) | 0 (0.0) | 4 (22.0) | 20 (16.5) |
| G9Pnt * | 1 (6.3) | 0 (0.0) | 0 (0.0) | 3 (30.0) | 1 (5.9) | 1 (5.6) | 6 (4.9) |
| G12P[8] | 0 (0.0) | 2 (6.9) | 2 (6.5) | 0 (0.0) | 7 (41.1) | 0 (0.0) | 11 (9.1) |
| * GntP[6] | 0 (0.0) | 0 (0.0) | 0 (0.0) | 1 (10) | 0 (0.0) | 0 (0.0) | 1 (0.8) |
| * GntPnt | 0 (0.0) | 2 (6.9) | 0 (0.0) | 0 (0.0) | 0 (0.0) | 0 (0.0) | 2 (1.7) |
| Total | 16 (100) | 29 (100) | 31 (100) | 10 (100) | 17 (100) | 18 (100) | 121 (100) |
| Age Group (Months) | |||||
|---|---|---|---|---|---|
| Genotype | 0–6 n (%) | 7–12 n (%) | 13–24 n (%) | >24 n (%) | Total |
| G1P[6] | 3 (3.4) | 2 (6.9) | 0 (0.0) | 0 (0.0) | 5 (4.1) |
| G1P[8] | 3 (3.4) | 0 (0.0) | 0 (0.0) | 0 (0.0) | 3 (2.5) |
| G1PN | 1 (1.1) | 0 (0.0) | 0 (0.0) | 0 (0.0) | 1 (0.8) |
| G2P[4] | 14 (16.1) | 4 (13.8) | 0 (0.0) | 0 (0.0) | 18 (14.9) |
| G3P[6] | 12 (13.8) | 3 (10.3) | 0 (0.0) | 1 (33.3) | 16 (13.2) |
| G3PN | 2 (2.3) | 0 (0.0) | 0 (0.0) | 0 (0.0) | 2 (1.6) |
| G8P[6] | 11 (12.6) | 3 (10.3) | 0 (0.0) | 0 (0.0) | 14 (11.5) |
| G8PN | 1 (1.1) | 0 (0.0) | 0 (0.0) | 0 (0.0) | 1 (1.7) |
| G9P[6] | 9 (10.3) | 8 (27.5) | 2 (100) | 2 (66.7) | 21 (17.3) |
| G9P[8] | 14 (16.1) | 6 (20.7) | 0 (0.0) | 0 (0.0) | 20 (16.5) |
| G9PN | 4 (4.6) | 2 (6.9) | 0 (0.0) | 0 (0.0) | 6 (4.9) |
| G12P[8] | 10 (11.5) | 1 (3.4) | 0 (0.0) | 0 (0.0) | 11 (9.1) |
| * GntP[6] | 1 (1.1) | 0 (0.0) | 0 (0.0) | 0 (0.0) | 1 (0.8) |
| * GntPN | 2 (2.9) | 0 (0.0) | 0 (0.0) | 0 (0.0) | 2 (1.7) |
| Total | 87 (100) | 29 (100) | 2 (100) | 3 (100) | 121 (100) |
| Severity of Diarrhea | ||||
|---|---|---|---|---|
| Genotype | Mild (<7) n (%) | Moderate (7–10) n (%) | Severe (≥11) n (%) | Total n (%) |
| G1P[6] | 0 (0.0) | 0 (0.0) | 5 (4.5) | 5 (4.1) |
| G1P[8] | 0 (0.0) | 1 (10.0) | 2 (1.8) | 3 (2.5) |
| G1Pnt | 0 (0.0) | 0 (0.0) | 1 (0.9) | 1 (0.8) |
| G2P[4] | 0 (0.0) | 0 (0.0) | 18 (16.5) | 18 (14.9) |
| G3P[6] | 0 (0.0) | 1 (10.0) | 15 (15.6) | 16 (13.2) |
| G3PN | 0 (0.0) | 0 (0.0) | 2 (1.8) | 2 (1.6) |
| G8P[6] | 0 (0.0) | 2 (20.0) | 12 (11.0) | 14 (11.5) |
| G8Pnt | 0 (0.0) | 0 (0.0) | 1 (0.9) | 1 (1.7) |
| G9P[6] | 2 (100) | 1 (10.0) | 18 (16.5) | 21 (17.3) |
| G9P[8] | 0 (0.0) | 2 (20.0) | 18 (16.5) | 20 (16.5) |
| G9PN | 0 (0.0) | 2 (20.0) | 4 (3.6) | 6 (4.9) |
| G12P[8] | 0 (0.0) | 1 (10.0) | 10 (9.2) | 11 (9.1) |
| * GntP[6] | 0 (0.0) | 0 (0.0) | 1 (0.9) | 1 (0.8) |
| * GntPN | 0 (0.0) | 0 (0.0) | 2 (1.8) | 2 (1.7) |
| Total | 2 (100) | 10 (100) | 109 (100) | 121 (100) |
| Lineage | Strain | VP4 (VP8 *) EPITOPE | ||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 8-1 | 8-2 | 8-3 | 8-4 | |||||||||||||||||||||||||||
| 100 | 146 | 148 | 150 | 188 | 190 | 192 | 193 | 194 | 195 | 196 | 180 | 183 | 113 | 114 | 115 | 116 | 125 | 131 | 132 | 133 | 135 | 87 | 88 | 89 | ||||||
| P[8] | III | RVA/Hu/AGO/DV105HMT/2021/G12P[8] | D | S | Q | D | S | T | N | L | N | G | I | T | A | N | P | V | D | N | R | N | D | D | N | T | T | |||
| RVA/Hu/AGO/DV117HMT/2021/G12P[8] | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | |||||
| RVA/Hu/AGO/DV66HMZ/2022/G12P[8] | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | |||||
| RVA/Hu/MWI/BID1LN/2013/G1P[8] | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | N | |||||
| RVA/Hu/USA/SSCRTV 00075/2015/G12P[8] | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | N | |||||
| Hu/GER126-08/GER/2008/G12P[8] | . | . | . | . | . | . | . | . | D | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | N | |||||
| RVA/Hu/AGO/CSP10/2013/G1P[8] | . | . | . | . | . | . | . | . | . | . | . | . | . | D | . | . | . | . | . | . | . | . | . | . | N | |||||
| RVA/Hu/BRA/BA20142/2011/G12P[8] | . | . | . | . | . | . | . | . | . | . | . | . | . | D | . | . | . | . | . | . | . | . | . | . | N | |||||
| RVA/Hu/FRA/E9779/2013/G1P[8] | . | . | . | . | . | . | . | . | D | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | N | |||||
| IV | RVA/Hu/BTN-49/BTN/2010/G9P[8] | . | . | . | E | . | . | D | . | T | S | . | . | . | D | . | . | . | S | . | . | . | N | . | . | N | ||||
| RVA/Hu/IND/MMC38/2009/GNDP[8] | . | G | . | E | . | . | D | . | T | S | . | . | . | D | . | . | . | S | . | . | . | N | . | . | N | |||||
| II | Vac/RotaTeq-WI79-4/USA/1992/G6P[8] | . | . | . | E | . | . | . | . | . | D | . | . | . | . | . | . | . | . | . | . | . | . | . | . | N | ||||
| RVA/Bethesda/DC140/1975/G3P[8] | . | . | . | E | . | . | . | . | . | D | . | . | . | . | . | . | . | . | . | . | . | . | . | . | N | |||||
| I | Vac/Rotarix/USA/2009/G1P[8] | . | . | . | E | . | . | . | . | . | N | . | . | . | . | . | . | . | S | S | . | . | N | . | . | N | ||||
| RVA/Hu/USA/Wa/1974/G1P[8] | . | . | . | E | . | . | . | . | . | N | . | . | . | . | . | . | . | S | S | . | . | N | . | . | N | |||||
| P[6] | II | RVA/porcine/Gottfried/1990/GNDP[6] | . | N | N | D | . | . | . | . | P | D | V | . | . | P | S | Q | . | V | E | . | S | . | I | N | K | |||
| Ia | RVA/Hu/AGO/DV114HKK/2021/G3P[6] | . | . | S | E | . | . | . | . | S | E | V | . | . | T | S | Q | S | T | E | . | N | S | T | N | Q | ||||
| RVA/Hu/AGO/DV68HKK/2021/G3P[6] | . | . | S | E | . | . | . | . | S | E | V | . | . | T | S | Q | S | T | E | . | N | S | T | N | Q | |||||
| RVA/Hu/AGO/DV177HKK/2021/G3P[6] | . | . | S | E | . | . | . | . | S | E | V | . | . | T | S | Q | S | T | E | . | N | S | T | N | Q | |||||
| RVA/Hu/AGO/CDV120HKK/2021/GNDP[6] | . | . | S | E | . | . | . | . | S | E | V | . | . | T | S | Q | S | T | E | . | N | S | T | N | Q | |||||
| RVA/Hu/AGO/C27/2012/G1P[6] | . | . | S | E | . | . | . | . | S | E | V | . | . | T | N | Q | S | T | E | . | N | S | T | N | Q | |||||
| Ib/c | RVA/swine/ITA/22104-7/2006/GNDP[6] | . | . | S | E | . | . | . | . | S | E | V | . | . | T | S | Q | S | T | E | . | N | N | T | N | Q | ||||
| IV | RVA/Hu/HUN/BP133800/2004/G4P[6] | . | N | S | E | . | . | . | . | S | E | . | . | . | T | N | Q | S | T | E | . | S | N | T | N | Q | ||||
| V | RVA/porcine/ITA/13404-7/2006/GNDP[6] | . | N | S | E | . | . | . | . | P | D | . | . | . | T | N | Q | S | M | E | . | N | S | T | N | Q | ||||
| Lineage | Strain | VP7 EPITOPE | ||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 7-1a | 7-1b | 7-2 | ||||||||||||||||||||||||||||||||
| 87 | 91 | 94 | 96 | 97 | 98 | 99 | 100 | 104 | 123 | 125 | 129 | 130 | 291 | 201 | 211 | 212 | 213 | 238 | 242 | 143 | 145 | 146 | 147 | 148 | 190 | 217 | 221 | 264 | ||||||
| G12 | II | RVA/Hu/AGO/DV117HMT/2021/G12P[8] | S | T | T | P | D | W | T | N | Q | D | S | V | D | … | Q | D | V | T | N | N | Q | Q | N | S | L | S | E | A | G | |||
| RVA/Hu/Py1135ASR07/PRY/2007/G12P[9] | . | . | . | . | . | . | . | . | . | . | A | . | . | K | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | |||||
| III | RVA/Hu/2119/PV/2011/G12P[ND] | . | . | . | . | . | . | . | . | . | . | . | . | . | … | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | ||||
| RVA/Hu/AGO/AH6/2012/G12P[6] | . | . | . | . | . | . | . | . | . | . | . | . | . | … | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | |||||
| I | Hu/L26/PHL/1987/G12P[4] | N | . | . | . | . | . | . | H | . | . | A | . | N | K | . | . | . | A | . | . | . | . | . | . | . | . | . | . | . | ||||
| IV | Po/RU172/IND/2002/G12PX | N | . | . | . | . | . | . | . | . | . | A | . | . | K | . | . | . | A | . | . | . | . | . | . | . | . | . | . | . | ||||
| G1 | Vac/Rotarix/USA/2009/G1P[8] | T | . | N | G | E | . | K | D | . | S | V | . | . | K | . | N | . | D | . | T | K | D | Q | N | . | . | M | N | . | ||||
| G3 | I | Vac/RotaTeq-W178-8/USA/1992/G3P[5] | T | . | N | N | S | . | K | D | . | . | A | . | . | K | . | . | A | N | K | D | K | D | A | T | . | . | . | . | . | |||
| RVA/Hu/AGO/DV114HKK/2021/G3P[6] | T | . | N | N | S | . | K | . | . | . | A | . | . | K | . | . | T | N | . | . | K | D | V | T | . | . | . | D | . | |||||
| RVA/Hu/AGO/DV177HKK/2021/G3P[6] | T | . | N | N | S | . | K | . | . | . | A | . | . | K | . | . | T | N | . | . | K | D | V | T | . | . | . | D | . | |||||
| RVA/Hu/AGO/DV68HKK/2021/G3P[6] | T | . | N | N | S | . | K | . | . | . | A | . | . | … | . | . | T | N | . | . | K | D | V | T | . | . | . | D | . | |||||
| II | RVA/Simian/USA/RRV/1975/G3P[3] | T | . | N | N | S | . | K | D | . | . | A | . | . | K | . | . | T | A | D | A | K | D | A | T | . | . | . | . | . | ||||
| III | RVA/Dog/USA/CU-1/1982/G3P[ND] | T | . | N | N | S | . | K | D | . | . | A | . | . | K | . | . | . | S | D | T | K | D | A | A | . | . | . | T | . | ||||
| IV | RVA/Rabbit/ITA/30-96/1996/G3P[14] | T | . | N | N | S | . | K | D | . | . | V | . | . | K | . | N | A | A | D | A | K | D | A | A | . | . | . | . | . | ||||
| G8 | V | RVA/Hu/AGO/DV540HGL/2022/G8P[ND] | … | … | … | … | … | … | … | … | … | … | … | … | … | K | . | . | T | . | . | T | … | … | … | … | … | . | . | . | . | |||
| RVA/Hu/AGO/DV34HECJ/2022/G8P[ND] | … | … | … | … | … | … | … | … | … | … | … | … | … | K | . | . | T | . | . | T | … | … | … | … | … | . | . | . | . | |||||
| RVA/Hu/AGO/CSMI10/2012/G8P[6] | A | . | A | N | S | . | K | D | . | . | A | I | N | … | . | . | T | . | . | T | K | N | T | N | S | . | . | . | . | |||||
| RVA/Hu/MWI/QEC287/2006/G8P[8] | A | . | A | S | S | . | K | D | . | . | A | I | N | … | . | . | T | . | . | T | K | N | A | N | S | . | . | . | . | |||||
| I | RVA/Hu/Croatia/CR2006/2006/G8P[8] | T | . | A | S | S | . | K | E | . | . | A | I | N | K | . | . | T | . | . | T | K | S | A | N | S | . | . | . | . | ||||
| II | RVA/Hu/ITA/PR1300/2004/G8P[14] | V | . | A | S | S | . | K | D | . | . | A | I | N | K | . | . | T | . | . | T | K | N | A | N | S | . | . | . | . | ||||
| III | RVA/Hu/AUS/BG8.01/1996/G8P[ND] | V | . | A | S | S | . | K | D | . | . | A | I | N | K | . | . | T | . | . | T | K | N | A | N | S | . | . | . | . | ||||
| IV | RVA/Hu/THA/TM-12505_60/2018/G8P[8] | T | . | A | S | S | . | K | D | . | . | A | I | N | … | . | . | T | . | . | T | K | N | A | D | S | . | . | . | . | ||||
| G9 | VI | RVA/Hu/AGO/DV72HMCC/2022/G9P[ND] | T | . | G | T | E | . | K | . | . | . | A | I | . | … | . | N | T | A | D | . | K | D | S | T | . | . | . | S | . | |||
| RVA/Hu/VNM/VE17426/2020/G9P[8] | T | . | G | T | E | . | K | . | . | . | A | I | . | K | . | N | T | A | D | . | K | D | S | T | . | . | . | S | . | |||||
| I | RVA/Hu/USA/F45/1987/G9P[ND] | A | . | G | T | E | . | K | D | . | . | A | I | . | K | . | N | T | A | D | T | K | D | S | T | . | . | . | S | . | ||||
| II | RVA/Hu/IND/116E/1986/G9P[11] | I | . | G | T | E | . | K | G | . | . | A | I | . | K | . | N | T | A | D | . | K | N | S | T | . | . | . | N | . | ||||
| III | RVA/Hu/Malawi/MW47/1997/G9P[ND] | T | . | G | T | E | . | K | D | . | . | A | I | . | K | . | N | T | A | D | . | K | D | S | T | . | . | . | S | . | ||||
| RVA/Hu/AGO/C84/2012/G9P[6] | T | . | G | T | E | . | K | D | . | . | A | I | . | … | . | N | T | A | D | . | K | D | S | T | . | . | . | S | . | |||||
| IV | RVA/Hu/CHN/97’SZ37/1997/G9P[ND] | T | . | G | T | E | . | K | D | . | . | A | I | . | K | . | N | T | A | D | . | K | D | S | T | . | . | . | S | . | ||||
| V | RVA/Hu/USA/OM46/1997/G9P[8] | T | . | G | T | E | . | K | D | . | . | A | I | . | K | . | N | T | A | D | . | K | D | S | T | . | . | . | S | . | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vita, D.G.; Santiso-Bellón, C.; Lemos, M.; Neto, Z.; Fortes-Gabriel, E.; Brito, M.; Sebastião, C.S.; Rodriguez-Diaz, J.; Cunha, C.; Istrate, C. Rotavirus alphagastroenteritidis: Circulating Strains After the Introduction of the Rotavirus Vaccine (Rotarix®) in Luanda Province of Angola. Viruses 2025, 17, 858. https://doi.org/10.3390/v17060858
Vita DG, Santiso-Bellón C, Lemos M, Neto Z, Fortes-Gabriel E, Brito M, Sebastião CS, Rodriguez-Diaz J, Cunha C, Istrate C. Rotavirus alphagastroenteritidis: Circulating Strains After the Introduction of the Rotavirus Vaccine (Rotarix®) in Luanda Province of Angola. Viruses. 2025; 17(6):858. https://doi.org/10.3390/v17060858
Chicago/Turabian StyleVita, Dikudila G., Cristina Santiso-Bellón, Manuel Lemos, Zoraima Neto, Elsa Fortes-Gabriel, Miguel Brito, Cruz S. Sebastião, Jesus Rodriguez-Diaz, Celso Cunha, and Claudia Istrate. 2025. "Rotavirus alphagastroenteritidis: Circulating Strains After the Introduction of the Rotavirus Vaccine (Rotarix®) in Luanda Province of Angola" Viruses 17, no. 6: 858. https://doi.org/10.3390/v17060858
APA StyleVita, D. G., Santiso-Bellón, C., Lemos, M., Neto, Z., Fortes-Gabriel, E., Brito, M., Sebastião, C. S., Rodriguez-Diaz, J., Cunha, C., & Istrate, C. (2025). Rotavirus alphagastroenteritidis: Circulating Strains After the Introduction of the Rotavirus Vaccine (Rotarix®) in Luanda Province of Angola. Viruses, 17(6), 858. https://doi.org/10.3390/v17060858