
Re-Emergence of Usutu Virus and Spreading of West Nile Virus Neuroinvasive Infections During the 2024 Transmission Season in Croatia

 ,
,  , , ,
, , ,  ,
,  , , , and
, , , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients with Neuroinvasive Disease Sampling
2.2. Horse and Bird Sampling
2.3. Mosquito Sampling
2.4. Molecular Diagnostics and Sequencing
2.5. Serology
2.6. Climate Data Collection and Analysis
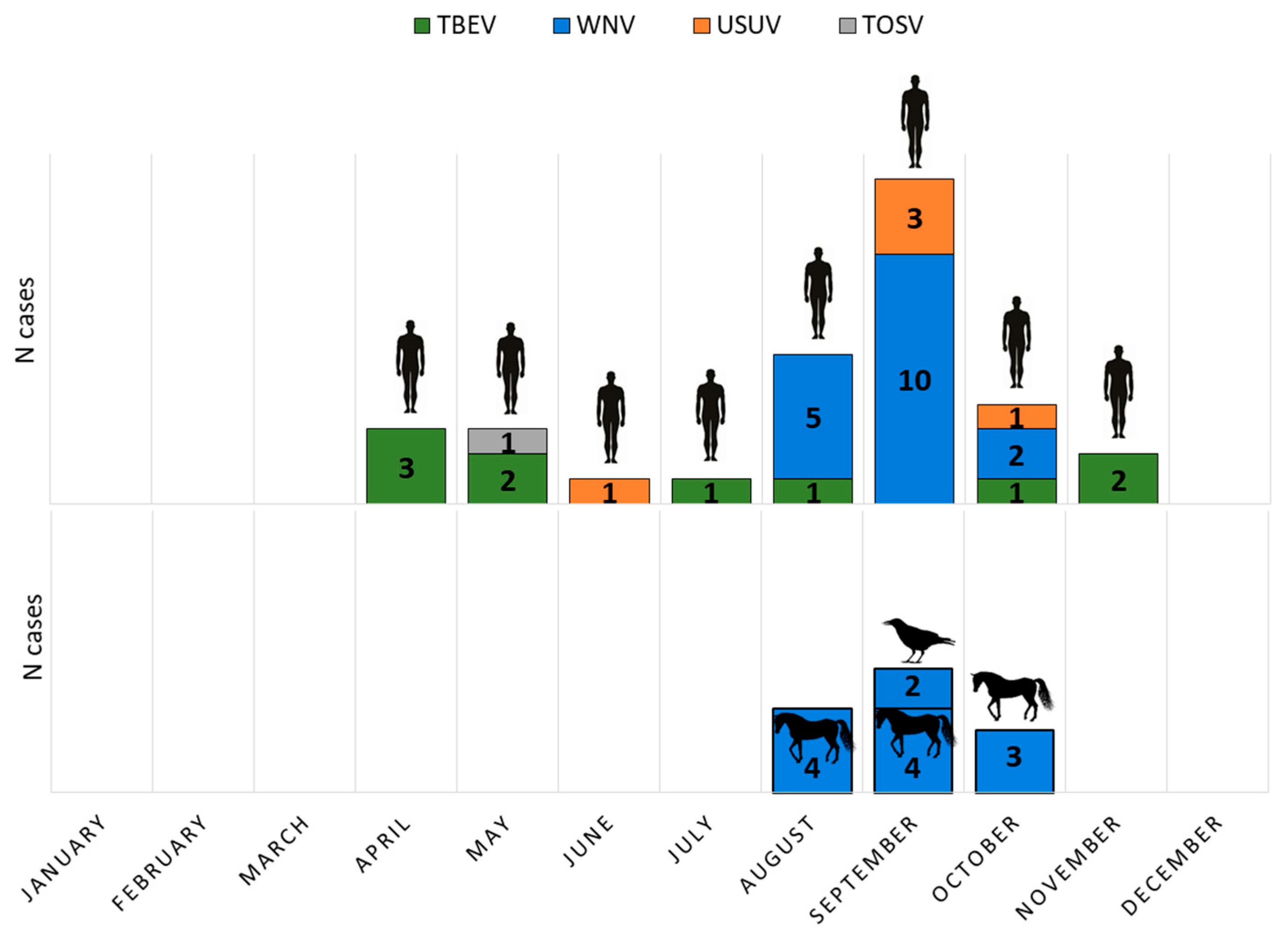
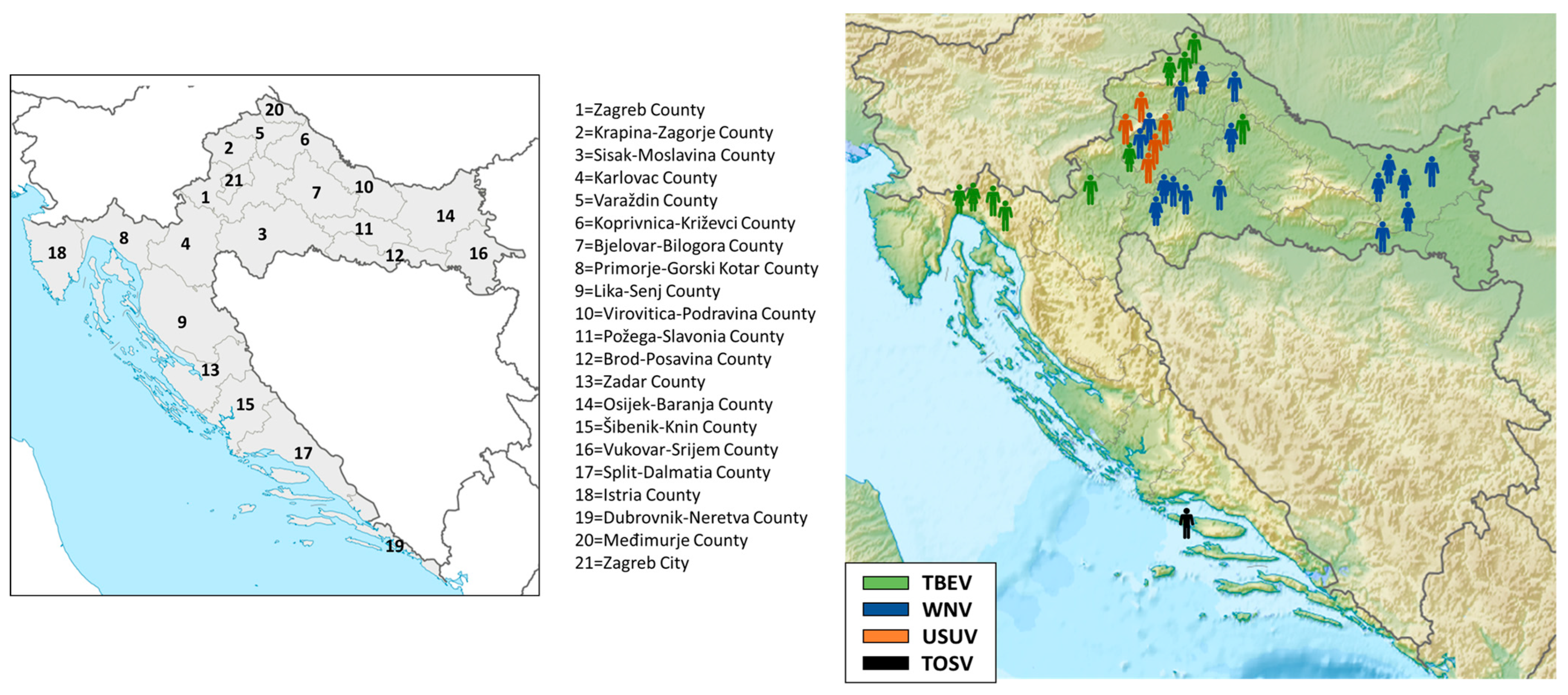
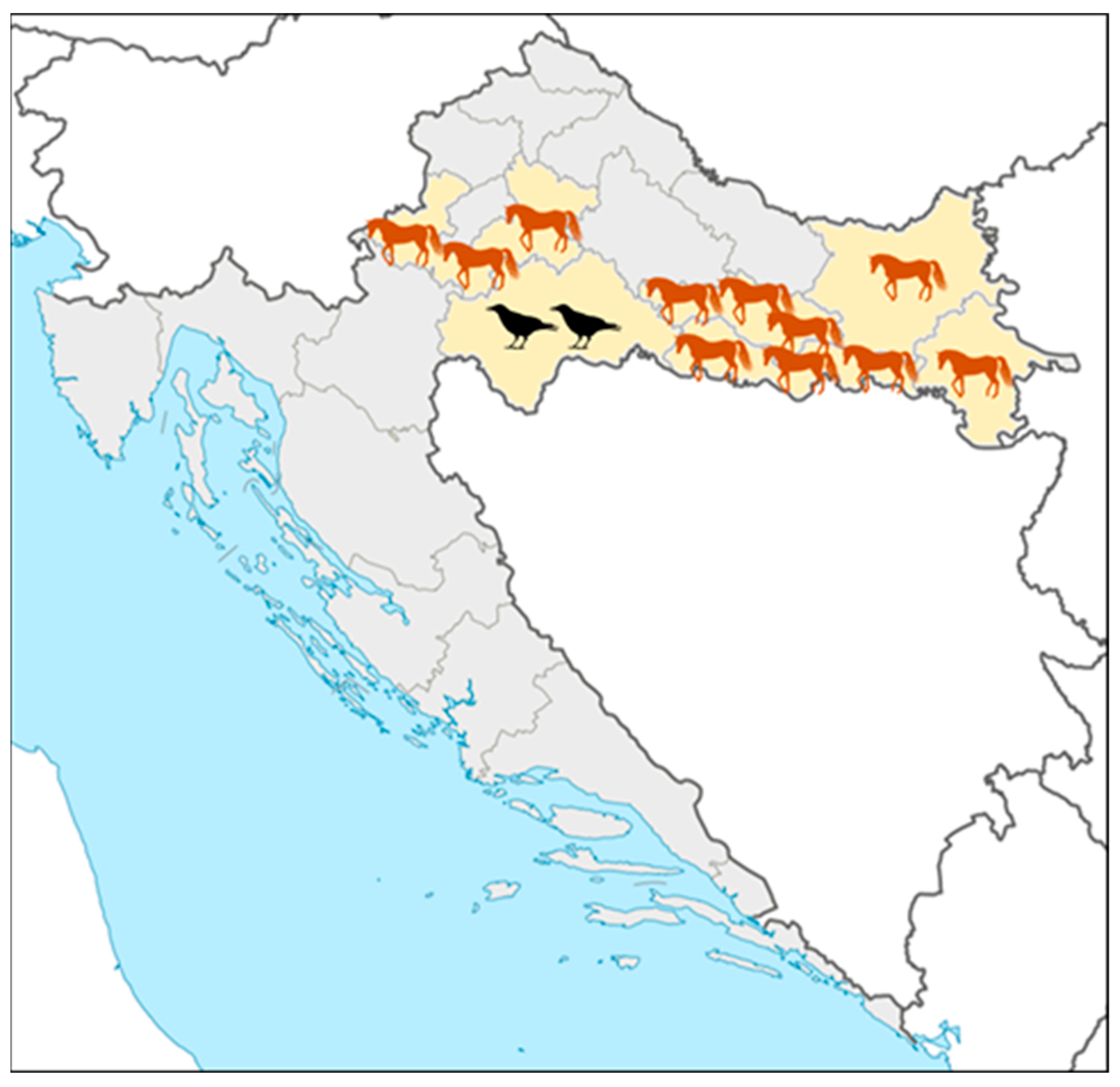
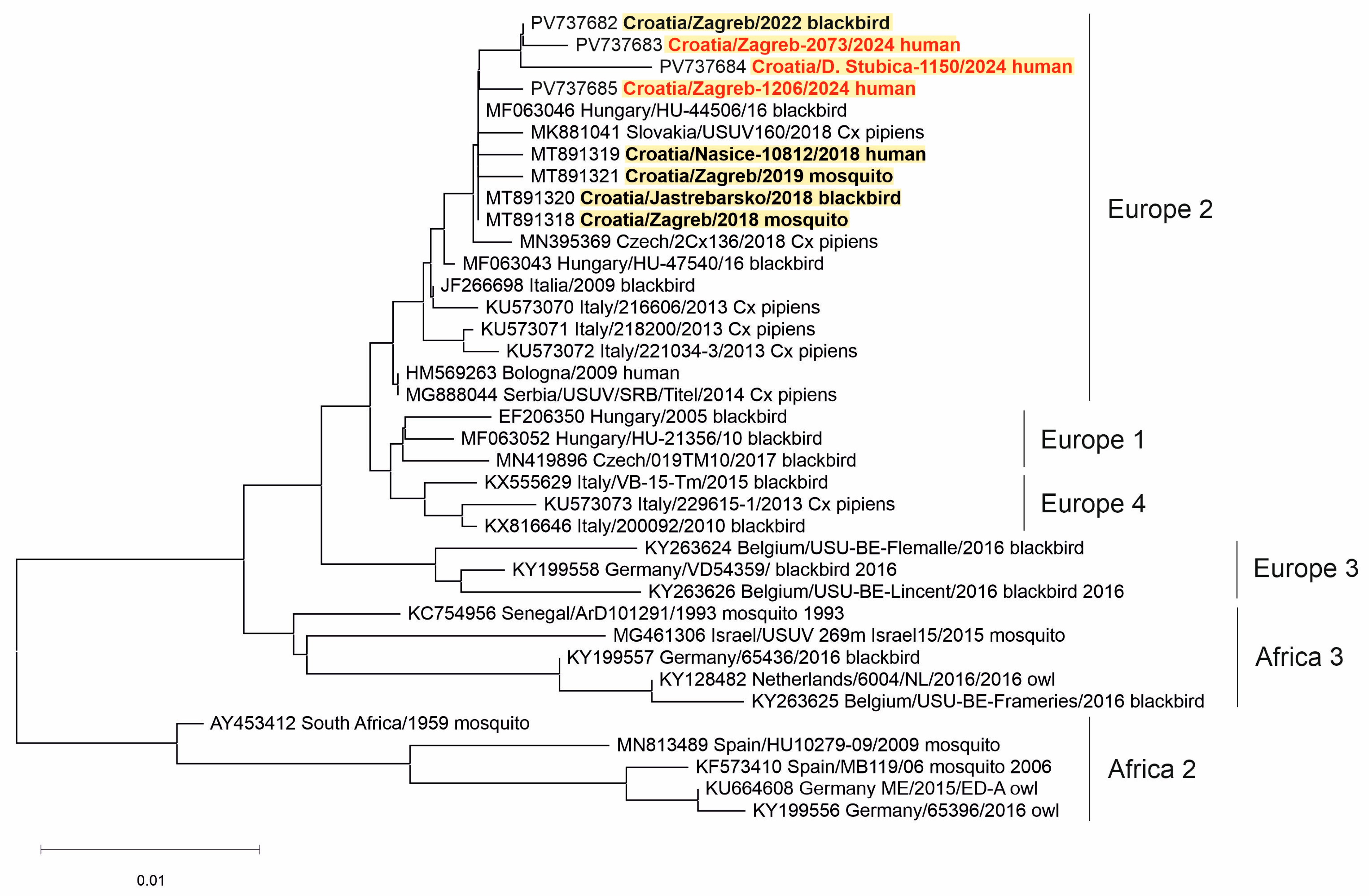
3. Results
3.1. Arboviral Infections in Patients with Neuroinvasive Diseases, Horses, and Birds
3.2. Flavivirus Seroprevalence in Horses
3.3. Mosquito Species Detected and Tested
3.4. Molecular Epidemiology of Arboviral Infections in Humans and Animals
3.5. Impact of Climate and Meteorological Conditions in 2024 on Arboviral Infections Occurrence
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization (WHO). Vector-Borne Diseases. Available online: https://www.who.int/news-room/fact-sheets/detail/vector-borne-diseases (accessed on 2 April 2025).
- Fleischmann, W.A.; Cao, L.C.; Nurjadi, D.; Velavan, T.P. Addressing the rise of autochthonous vector-borne diseases in a warming Europe. Int. J. Infect. Dis. 2024, 149, 107275. [Google Scholar] [CrossRef] [PubMed]
- Erazo, D.; Grant, L.; Ghisbain, G.; Marini, G.; Colón-González, F.J.; Wint, W.; Rizzoli, A.; Van Bortel, W.; Vogels, C.B.F.; Grubaugh, N.D.; et al. Contribution of climate change to the spatial expansion of West Nile virus in Europe. Nat. Commun. 2024, 15, 1196. [Google Scholar] [CrossRef] [PubMed]
- Daniel, M.; Danielová, V.; Fialová, A.; Malý, M.; Kříž, B.; Nuttall, P.A. Increased Relative Risk of Tick-Borne Encephalitis in Warmer Weather. Front. Cell. Infect. Microbiol. 2018, 8, 90. [Google Scholar] [CrossRef] [PubMed]
- Vilibić-Čavlek, T.; Bogdanić, M.; Savić, V.; Barbić, L.; Stevanović, V.; Kaić, B. Tick-borne Human Diseases around the Globe. In The TBE Book; Dobler, G., Erber, W., Bröker, M., Chitimia-Dobler, L., Schmitt, H.J., Eds.; Global Health Press: Singapore, 2024; Chapter 1; pp. 1–7. [Google Scholar] [CrossRef]
- Choi, H.; Lee, C.H. The impact of climate change on ecology of tick associated with tick-borne diseases. PLoS Comput. Biol. 2025, 21, e1012903. [Google Scholar] [CrossRef]
- Vilibić-Čavlek, T.; Savić, V.; Židovec-Lepej, S.; Bogdanić, M.; Stevanović, V.; Barbić, L. Emerging and Neglected Viral Zoonoses in Europe [Internet]. Current Topics in Zoonoses. IntechOpen. 2024. Available online: https://www.intechopen.com/chapters/88260 (accessed on 5 April 2025).
- Simonin, Y. Circulation of West Nile Virus and Usutu Virus in Europe: Overview and Challenges. Viruses 2024, 16, 599. [Google Scholar] [CrossRef]
- Ayhan, N.; Prudhomme, J.; Laroche, L.; Bañuls, A.L.; Charrel, R.N. Broader geographical distribution of Toscana virus in the Mediterranean region suggests the existence of larger varieties of sand fly vectors. Microorganisms 2020, 8, 114. [Google Scholar] [CrossRef]
- Keskek Turk, Y.; Ergunay, K.; Kohl, A.; Hughes, J.; McKimmie, C.S. Toscana virus—An emerging Mediterranean arbovirus transmitted by sand flies. J. Gen. Virol. 2024, 105, 002045. [Google Scholar] [CrossRef]
- Dobler, G.; Tkachev, S. General epidemiology of TBE. In The TBE Book, 6th ed.; Dobler, G., Erber, W., Bröker, M., Schmitt, H.J., Eds.; Global Health Press: Singapore, 2023; Chapter 11; pp. 1–7. [Google Scholar] [CrossRef]
- Rizzoli, A.; Jiménez-Clavero, M.A.; Barzon, L.; Cordioli, P.; Figuerola, J.; Koraka, P.; Martina, B.; Moreno, A.; Nowotny, N.; Pardigon, N.; et al. The challenge of West Nile virus in Europe: Knowledge gaps and research priorities. Euro Surveill. 2015, 20, 28–42. [Google Scholar] [CrossRef]
- Koch, R.T.; Erazo, D.; Folly, A.J.; Johnson, N.; Dellicour, S.; Grubaugh, N.D.; Vogels, C.B.F. Genomic epidemiology of West Nile virus in Europe. One Health 2023, 18, 100664. [Google Scholar] [CrossRef]
- Cadar, D.; Lühken, R.; van der Jeugd, H.; Garigliany, M.; Ziegler, U.; Keller, M.; Lahoreau, J.; Lachmann, L.; Becker, N.; Kik, M.; et al. Widespread activity of multiple lineages of Usutu virus, western Europe, 2016. Euro Surveill. 2017, 22, 11–17. [Google Scholar] [CrossRef]
- Bergmann, F.; Holicki, C.M.; Michel, F.; Bock, S.; Scuda, N.; Priemer, G.; Kenklies, S.; Siempelkamp, T.; Skuballa, J.; Sauerwald, C.; et al. Reconstruction of the molecular evolution of Usutu virus in Germany: Insights into virus emergence and circulation. PLoS Negl. Trop. Dis. 2023, 17, e0011203. [Google Scholar] [CrossRef] [PubMed]
- Cadar, D.; Simonin, Y. Human Usutu virus infections in Europe: A new risk on the horizon? Viruses 2022, 15, 77. [Google Scholar] [CrossRef]
- Pustijanac, E.; Buršić, M.; Talapko, J.; Škrlec, I.; Meštrović, T.; Lišnjić, D. Tick-borne encephalitis virus: A comprehensive review of transmission, pathogenesis, epidemiology, clinical manifestations, diagnosis, and prevention. Microorganisms 2023, 11, 1634. [Google Scholar] [CrossRef]
- Vinokur, M.; Small, G. West Nile radiculomyelitis. Neurology 2019, 92 (Suppl. 15), 4–9. [Google Scholar] [CrossRef]
- Nikolić, N.; Poluga, J.; Milošević, I.; Todorović, N.; Filipović, A.; Jegorović, B.; Mitrović, N.; Karić, U.; Gmizić, I.; Stevanović, G.; et al. Neurological and neuromuscular manifestations in patients with West Nile neuroinvasive disease, Belgrade area, Serbia, season 2022. Neurol. Sci. 2024, 45, 719–726. [Google Scholar] [CrossRef]
- Santini, M.; Zupetić, I.; Visković, K.; Krznarić, J.; Kutlesa, M.; Krajinović, V.; Polak, V.L.; Savić, V.; Tabain, I.; Barbić, L.; et al. Cauda equina arachnoiditis—A rare manifestation of West Nile virus neuroinvasive disease: A case report. World J. Clin. Cases 2020, 8, 3797–3803. [Google Scholar] [CrossRef] [PubMed]
- Agliani, G.; Giglia, G.; Marshall, E.M.; Gröne, A.; Rockx, B.H.G.; van den Brand, J.M.A. Pathological features of West Nile and Usutu virus natural infections in wild and domestic animals and in humans: A comparative review. One Health 2023, 16, 100525. [Google Scholar] [CrossRef]
- Ayhan, N.; Eldin, C.; Charrel, R. Toscana virus infection clinical characterization. medRxiv 2024. [Google Scholar] [CrossRef]
- Kutasi, O.; Fehér, O.; Sárdi, S.; Balogh, N.; Nagy, A.; Moravszki, L.; Bódai, E.; Szenci, O. Characterisation of the cerebrospinal fluid of horses with West Nile virus neuroinvasive disease. Acta Vet. Hung. 2020, 68, 177–185. [Google Scholar] [CrossRef]
- de Heus, P.; Bagó, Z.; Weidinger, P.; Lale, D.; Trachsel, D.S.; Revilla-Fernández, S.; Matiasek, K.; Nowotny, N. Severe neurologic disease in a horse caused by Tick-Borne Encephalitis Virus, Austria, 2021. Viruses 2023, 15, 2022. [Google Scholar] [CrossRef]
- Bröker, M. Tick-Borne Encephalitis (TBE) in 2024: A Preliminary Assessment. Available online: https://tbenews.com/tbe/tick-borne-encephalitis-tbe-in-2024-a-preliminary-assessment/ (accessed on 6 April 2025).
- European Centre for Disease Prevention and Control (ECDC). Surveillance of West Nile Virus Infections in Humans and Animals in Europe, Monthly Report. Available online: https://www.ecdc.europa.eu/en/infectious-disease-topics/west-nile-virus-infection/surveillance-and-disease-data/monthly-updates (accessed on 6 April 2025).
- Ilic, M.; Barbic, L.; Bogdanic, M.; Tabain, I.; Savić, V.; Kosanovic Licina, M.L.; Kaić, B.; Jungic, A.; Vucelja, M.; Angelov, V.; et al. Tick-borne encephalitis outbreak following raw goat milk consumption in a new micro-location, Croatia, June 2019. Ticks Tick Borne Dis. 2020, 11, 101513. [Google Scholar] [CrossRef] [PubMed]
- Vilibic-Cavlek, T.; Krcmar, S.; Bogdanic, M.; Tomljenovic, M.; Barbic, L.; Roncevic, D.; Sabadi, D.; Vucelja, M.; Santini, M.; Hunjak, B.; et al. An Overview of Tick-Borne Encephalitis Epidemiology in Endemic Regions of Continental Croatia, 2017–2023. Microorganisms 2024, 12, 386. [Google Scholar] [CrossRef] [PubMed]
- Vilibic-Cavlek, T.; Savić, V.; Klobucar, A.; Ferenc, T.; Ilic, M.; Bogdanic, M.; Tabain, I.; Stevanović, V.; Santini, M.; Curman Posavec, M.; et al. Emerging Trends in the West Nile Virus Epidemiology in Croatia in the ‘One Health’ Context, 2011–2020. Trop. Med. Infect. Dis. 2021, 6, 140. [Google Scholar] [CrossRef]
- Bogdanic, M.; Savić, V.; Klobucar, A.; Barbic, L.; Sabadi, D.; Tomljenovic, M.; Madić, J.; Hruskar, Z.; Curman Posavec, M.; Santini, M.; et al. The Re-Emergence of Neuroinvasive Flaviviruses in Croatia During the 2022 Transmission Season. Microorganisms 2024, 12, 2210. [Google Scholar] [CrossRef]
- Santini, M.; Vilibic-Cavlek, T.; Barsic, B.; Barbic, L.; Savić, V.; Stevanović, V.; Listes, E.; Di Gennaro, A.; Savini, G. First cases of human Usutu virus neuroinvasive infection in Croatia, August–September 2013: Clinical and laboratory features. J. Neurovirol. 2015, 21, 92–97. [Google Scholar] [CrossRef]
- Vilibic-Cavlek, T.; Savić, V.; Sabadi, D.; Peric, L.; Barbic, L.; Klobucar, A.; Miklausic, B.; Tabain, I.; Santini, M.; Vucelja, M.; et al. Prevalence and molecular epidemiology of West Nile and Usutu virus infections in Croatia in the ‘One Health’ context, 2018. Transbound. Emerg. Dis. 2019, 66, 1946–1957. [Google Scholar] [CrossRef] [PubMed]
- European Centre for Disease Prevention and Control (ECDC). EU Case Definitions. Available online: https://www.ecdc.europa.eu/en/all-topics/eu-case-definitions (accessed on 16 May 2025).
- Vilibic-Cavlek, T.; Bogdanic, M.; Savic, V.; Hruskar, Z.; Barbic, L.; Stevanovic, V.; Antolasic, L.; Milasincic, L.; Sabadi, D.; Miletic, G.; et al. Diagnosis of West Nile virus infections: Evaluation of different laboratory methods. World J. Virol. 2024, 13, 95986. [Google Scholar] [CrossRef]
- Tuncel, G.; Akan, G.; Kalaycı, M.; Baddal, B.; Bostancı, A.; Suer, K.; Özverel, C.S.; Şanlıdağ, T. Efficient Detection of West Nile Virus in Urine Specimens by a Novel In-House RT-qPCR Detection Kit. Can. J. Infect. Dis. Med. Microbiol. 2025, 2025, 6513971. [Google Scholar] [CrossRef]
- Becker, N.; Petric, D.; Zgomba, M.; Boase, C.; Madon, M.; Dahl, C.; Kaiser, A. Mosquitoes: Identification, Ecology and Control, 3rd ed.; Springer: Berlin/Heidelberg, Germany, 2020. [Google Scholar]
- Schwaiger, M.; Cassinotti, P. Development of a quantitative real-time RT-PCR assay with internal control for the laboratory detection of tick-borne encephalitis virus (TBEV) RNA. J. Clin. Virol. 2003, 27, 136–145. [Google Scholar] [CrossRef]
- Tang, Y.; Hapip, C.A.; Liu, B.; Fang, C.T. Highly sensitive TaqMan RT-PCR assay for detection and quantification of both lineages of West Nile virus RNA. J. Clin. Virol. 2006, 36, 177–182. [Google Scholar] [CrossRef]
- Nikolay, B.; Weidmann, M.; Dupressoir, A.; Faye, O.; Boye, C.S.; Diallo, M.; Sall, A.A. Development of a Usutu virus specific real-time reverse transcription PCR assay based on sequenced strains from Africa and Europe. J. Virol. Methods 2014, 197, 51–54. [Google Scholar] [CrossRef]
- Weidmann, M.; Sanchez-Seco, M.P.; Sall, A.A.; Ly, P.O.; Thiongane, Y.; Lô, M.M.; Schley, H.; Hufert, F.T. Rapid detection of important human pathogenic Phleboviruses. J. Clin. Virol. 2008, 41, 138–142. [Google Scholar] [CrossRef]
- Weissenböck, H.; Kolodziejek, J.; Url, A.; Lussy, H.; Rebel-Bauder, B.; Nowotny, N. Emergence of Usutu virus, an African mosquito-borne flavivirus of the Japanese encephalitis virus group, in central Europe. Emerg. Infect. Dis. 2002, 8, 652–656. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Di Gennaro, A.; Lorusso, A.; Casaccia, C.; Conte, A.; Monaco, F.; Savini, G. Serum neutralization assay can efficiently replace plaque reduction neutralization test for detection and quantitation of West Nile virus antibodies in human and animal serum samples. Clin. Vaccine Immunol. 2014, 21, 1460–1462. [Google Scholar] [CrossRef]
- Cindrić, K.; Juras, J.; Pasarić, Z. On precipitation monitoring with theoretical statistical distributions. Theor. Appl. Climatol. 2019, 136, 145–156. [Google Scholar] [CrossRef]
- Croatian Meteorological and Hydrological Service. Climate Data. Available online: https://meteo.hr/index_en.php (accessed on 6 April 2025).
- Zady, M.F. Correlation and Simple Least Squares Regression. October 2000. Available online: http://www.westgard.com/lesson44.htm (accessed on 30 May 2025).
- EEA Geospatial Data Catalogue. Biogeographical Regions, Europe 2016, ver. 1. Available online: https://sdi.eea.europa.eu/catalogue/geoss/api/records/c6d27566-e699-4d58-a132-bbe3fe01491b (accessed on 30 March 2025).
- Van Heuverswyn, J.; Hallmaier-Wacker, L.K.; Beauté, J.; Gomes Dias, J.; Haussig, J.M.; Busch, K.; Kerlik, J.; Markowicz, M.; Mäkelä, H.; Nygren, T.M.; et al. Spatiotemporal spread of tick-borne encephalitis in the EU/EEA, 2012 to 2020. Euro Surveill. 2023, 28, 2200543. [Google Scholar] [CrossRef]
- Bago, J.; Bjedov, L.; Vucelja, M.; Tomljanović, K.; Cetinić Balent, N.; Zember, S.; Margaletić, J.; Đaković Rode, O. The Influence of Biogeographic Diversity, Climate and Wildlife on the Incidence of Tick-Borne Encephalitis in Croatia. Viruses 2025, 17, 266. [Google Scholar] [CrossRef]
- Mor, Z.; Omari, H.; Indenbaum, V.; Kirstein, O.D.; Shatach Catabi, O.; Reicher, S.; Lustig, Y.; Davidovich-Cohen, M.; Kaliner, E.; Sheffer, R.; et al. Early rise of West Nile fever in Israel, June 2024. Euro Surveill. 2024, 29, 2400457. [Google Scholar] [CrossRef]
- Giglia, G.; Gianfilippo, A.; Mandara, M.T.; de Bruin, E.; Gröne, A.; van den Brand, J.M.A. Usutu virus avian and human infection after more than 25 years of circulation. Eur. J. Clin. Microbiol. Infect. Dis. 2024, 43, 805–807. [Google Scholar] [CrossRef]
- Campbell, L.G.; Martin, A.A.; Lanciotti, R.S.; Gubler, D.J. West Nile Virus. Lancet Infect. Dis. 2002, 2, 519–529. [Google Scholar] [CrossRef]
- Reiter, P. Climate change and mosquito-borne disease. Environ. Health Perspect. 2001, 109, 141–161. [Google Scholar]
- World Meteorological Organization (WMO). State of the Global Climate 2024 (WMO-No. 1368); WMO: Geneva, Switzerland, 2025. [Google Scholar]
- Cruz-Pacheco, G.; Esteva, L.; Vargas, C. Seasonality and outbreaks in West Nile virus infection. Bull. Math. Biol. 2009, 71, 1378–1393. [Google Scholar] [CrossRef]
- Marini, G.; Drakulovic, M.B.; Jovanovic, V.; Dagostin, F.; Wint, W.; Tagliapietra, V.; Vasic, M.; Rizzoli, A. Drivers and epidemiological patterns of West Nile virus in Serbia. Front. Public Health 2024, 12, 1429583. [Google Scholar] [CrossRef]
- Marini, G.; Pugliese, A.; Wint, W.; Alexander, N.S.; Rizzoli, A.; Rosà, R. Modelling the West Nile virus force of infection in the European human population. One Health 2022, 15, 100462. [Google Scholar] [CrossRef]
- Holicki, C.M.; Ziegler, U.; Răileanu, C.; Kampen, H.; Werner, D.; Schulz, J.; Silaghi, C.; Groschup, M.H.; Vasić, A. West Nile virus lineage 2 vector competence of indigenous Culex and Aedes mosquitoes from Germany at temperate climate conditions. Viruses 2020, 12, 561. [Google Scholar] [CrossRef]
- Kouroupis, D.; Charisi, K.; Pyrpasopoulou, A. The Ongoing Epidemic of West Nile Virus in Greece: The Contribution of Biological Vectors and Reservoirs and the Importance of Climate and Socioeconomic Factors Revisited. Trop. Med. Infect. Dis. 2023, 8, 453. [Google Scholar] [CrossRef]
- Sigfrid, L.; Chan, X.H.S.; Kasbergen, L.M.R.; Hookham, L.; Wei, J.; Chen, S.; Lee, J.L.; Sikkema, R.S.; de Bruin, E.; Corman, V.M.; et al. Prevalence, clinical management, and outcomes of adults hospitalised with endemic arbovirus illness in southeast Europe (MERMAIDS-ARBO): A prospective observational study. Lancet Infect. Dis. 2025, 25, 690–700. [Google Scholar] [CrossRef]
- Riccardo, F.; Bella, A.; Monaco, F.; Ferraro, F.; Petrone, D.; Mateo-Urdiales, A.; Andrianou, X.D.; Del Manso, M.; Venturi, G.; Fortuna, C.; et al. Italian Arbovirus Surveillance network. Rapid increase in neuroinvasive West Nile virus infections in humans, Italy, July 2022. Euro Surveill. 2022, 27, 2200653. [Google Scholar] [CrossRef]
- Vignjević, G.; Bušić, N.; Turić, N.; Varga, Z.; Zana, B.; Ábrahám, Á.; Kurucz, K.; Vrućina, I.; Merdić, E. First Detection of West Nile Virus Lineage 2 in Culex pipiens Vectors in Croatia. Pathogens 2024, 13, 1131. [Google Scholar] [CrossRef]
- Klobucar, A.; Savic, V.; Curman Posavec, M.; Petrinic, S.; Kuhar, U.; Toplak, I.; Madic, J.; Vilibic-Cavlek, T. Screening of mosquitoes for West Nile virus and Usutu virus in Croatia, 2015–2020. Trop. Med. Infect. Dis. 2021, 6, 45. [Google Scholar] [CrossRef]
- Valassina, M.; Meacci, F.; Valensin, P.E.; Cusi, M.G. Detection of neurotropic viruses circulating in Tuscany: The incisive role of Toscana virus. J. Med. Virol. 2000, 60, 86–90. [Google Scholar] [CrossRef]
- Fotakis, E.A.; Di Maggio, E.; Del Manso, M.; Mateo-Urdiales, A.; Petrone, D.; Fabiani, M.; Perego, G.; Bella, A.; Bongiorno, G.; Bernardini, I.; et al. Italian Arbovirus Surveillance Network. Human neuroinvasive Toscana virus infections in Italy from 2016 to 2023: Increased incidence in 2022 and 2023. Euro Surveill. 2025, 30, 2400203. [Google Scholar] [CrossRef] [PubMed]
- Vilibic-Cavlek, T.; Zidovec-Lepej, S.; Ledina, D.; Knezevic, S.; Savic, V.; Tabain, I.; Ivic, I.; Slavuljica, I.; Bogdanic, M.; Grgic, I.; et al. Clinical, virological, and immunological findings in patients with Toscana neuroinvasive disease in Croatia: Report of three cases. Trop. Med. Infect. Dis. 2020, 5, 144. [Google Scholar] [CrossRef] [PubMed]
- Ayhan, N.; Alten, B.; Ivovic, V.; Martinkovic, F.; Kasap, O.E.; Ozbel, Y.; de Lamballerie, X.; Charrel, R.N. Cocirculation of two lineages of Toscana virus in Croatia. Front. Public Health 2017, 5, 336. [Google Scholar] [CrossRef]
- Kasbergen, L.M.R.; de Bruin, E.; Chandler, F.; Sigfrid, L.; Chan, X.H.S.; Hookham, L.; Wei, J.; Chen, S.; GeurtsvanKessel, C.H.; Scherbeijn, S.; et al. Multi-antigen serology and a diagnostic algorithm for the detection of arbovirus infections as novel tools for arbovirus preparedness in southeast Europe (MERMAIDS-ARBO): A prospective observational study. Lancet Infect. Dis. 2025, 25, 678–689. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | Serology Method | Kit Name |
|---|---|---|
| TBEV | IgM/IgG ELISA, IgG avidity, VNT (in-house) | Anti-TBEV IgM ELISA; Anti-TBEV IgG ELISA 2.0; TBEV-Avidity ELISA (IgG); Euroimmun, Lübeck, Germany |
| WNV | IgM/IgG ELISA, IgG avidity, VNT (in-house) | Anti-WNV IgM ELISA; Anti-WNV IgG ELISA; WNV-Avidity ELISA (IgG); Euroimmun, Lübeck, Germany |
| USUV | IgG ELISA, VNT (in-house) | Anti-USUV IgG ELISA; Euroimmun, Lübeck, Germany |
| TOSV | IgM/IgG IFA | Sandfly Fever Mosaic IIF; Euroimmun, Lübeck, Germany |
| Virus | Acute Infections; N/% | IgG Seropositive; N/% (95%CI) |
|---|---|---|
| TBEV | 10/6.49 | 3/1.95 (0.40–5.59) |
| WNV | 17/11.03 | 1/0.65 (0.02–3.56) |
| USUV | 5/3.24 | 1/0.65 (002–3.56) |
| TOSV | 1/0.64 | 0/0 (0–2.37) * |
| Detected Virus | Serum ELISA N (%) Positive | CSF ELISA N (%) Positive | RT-qPCR N (%) Positive | Sequencing N | |||
|---|---|---|---|---|---|---|---|
| IgM | IgG | IgM | IgG | CSF | Urine | ||
| TBEV (n = 10) | 10 (100) | 9 (90.00) | 10 (100) | 8 (80.00) | 0 (0) | 0 (0) | 0 |
| WNV (n = 17) | 12 (70.58) | 10 (58.82) | 11 (91.66) | 3 (17.64) | 0 (0) | 5 (29.41) | 1 |
| USUV (n = 5) | NT | 2 (NA) | NT | 0 (NA) | 4 (NA) | 2 (NA) | 3 |
| Sample No. | ELISA TBEV | ELISA WNV | ELISA USUV | RT-qPCR CSF/Urine | Serum VNT Titer | |||
|---|---|---|---|---|---|---|---|---|
| Serum IgM/IgG | CSF IgM/IgG | Serum IgM/IgG | CSF IgM/IgG | Serum IgG | CSF IgG | |||
| 1 | Positive/ Positive | Negative/ Positive | Negative/ Positive | Negative/ Positive | Negative | Negative | Negative/ Negative | TBEV 20 |
| 2 | Negative/ Positive | Negative/ Negative | Positive/ Positive | Positive/ Negative | Negative | Negative | WNV urine positive | NT |
| 3 | Negative/ Positive | Negative/ Negative | Positive/ Positive | Negative/ Positive | Negative | Negative | WNV urine positive | NT |
| 4 | Negative/ Positive | Negative/ Negative | Positive/ Positive | Negative/ Positive | Positive | Negative | Negative/ Negative | WNV 10 |
| 5 | Negative/ Negative | Negative/ Negative | Positive/ Negative | Negative/ Negative | Positive | Negative | Negative/ Negative | USUV 40 |
| County | WNV IgG | TBEV IgG | ||||
|---|---|---|---|---|---|---|
| N | N (%) Positive | 95%CI | N | N (%) Positive | 95%CI | |
| Zagreb | 115 | 24 (20.87) | 13.85–29.44 | 9 | 0 (0) | 0–33.63 * |
| Krapina-Zagorje | 22 | 0 (0) | 0–15.44 * | 20 | 1 (5.00) | 0.13–24.87 |
| Sisak-Moslavina | 26 | 2 (7.69) | 0.95–25.13 | 19 | 3 (15.79) | 3.38–39.58 |
| Karlovac | 19 | 1 (5.26) | 0.13–26.03 | 15 | 1 (6.67) | 0.17–31.95 |
| Varaždin | 66 | 10 (15.15) | 7.51–26.10 | 11 | 1 (9.09) | 0.23–41.28 |
| Koprivnica-Križevci | 33 | 7 (21.21) | 8.98–38.91 | 11 | 3 (27.27) | 6.02–60.97 |
| Bjelovar-Bilogora | 72 | 9 (12.50) | 5.88–22.41 | 17 | 4 (23.53) | 6.81–49.90 |
| Primorje-Gorski Kotar | 66 | 3 (4.55) | 0.95–12.71 | 39 | 5 (12.82) | 4.30–27.43 |
| Lika-Senj | 22 | 2 (9.09) | 1.12–29.16 | 51 | 2 (3.92) | 0.48–13.46 |
| Virovitica-Podravina | 95 | 21 (22.11) | 14.23–31.78 | 20 | 3 (15.00) | 0.21–37.89 |
| Požega-Slavonia | 46 | 7 (15.22) | 6.34–28.87 | 16 | 3 (18.75) | 4.05–45.65 |
| Brod-Posavina | 86 | 20 (23.3) | 14.8–33.6 | 21 | 3 (14.3) | 3.1–36.3 |
| Zadar | 6 | 1 (NA) | NA | 19 | 0 (0) | 0–17.65 * |
| Osijek-Baranja | 202 | 101 (50.00) | 42.90–57.10 | 21 | 7 (33.33) | 14.59–56.97 |
| Šibenik-Knin | NT | NA | NA | 20 | 1 (5.00) | 0.13–24.87 |
| Vukovar-Srijem | 73 | 40 (54.79) | 42.7–66.5 | 22 | 16 (72.73) | 49.8–89.3 |
| Split-Dalmatia | 16 | 1 (6.25) | 0.16–30.23 | 28 | 0 (0) | 0–12.34 * |
| Istria | 45 | 3 (6.67) | 1.40–18.27 | 35 | 4 (11.43) | 3.20–26.74 |
| Dubrovnik-Neretva | NT | NA | NA | NT | NA | NA |
| Međimurje | 53 | 7 (13.21) | 5.48–25.34 | 13 | 2 (15.38) | 1.92–45.45 |
| Zagreb City | 105 | 17 (16.19) | 9.72–24.65 | 21 | 9 (42.86) | 21.82–65.98 |
| Mosquito Species | Total (%) | Number of Mosquito Pools |
|---|---|---|
| Anopheles claviger | 2 (0.03) | 1 |
| Anopheles plumbeus | 55 (0.71) | 11 |
| Aedes albopictus | 286 (3.70) | 44 |
| Aedes cantans | 53 (0.69) | 4 |
| Aedes cinereus | 35 (0.45) | 4 |
| Aedes geniculatus | 49 (0.63) | 6 |
| Aedes rossicus | 3 (0.04) | 1 |
| Aedes rusticus | 383 (4.96) | 12 |
| Aedes sticticus | 1556 (20.14) | 53 |
| Aedes vexans | 5152 (66.68) | 126 |
| Coquillettidia richiardii | 24 (0.31) | 2 |
| Culex pipiens complex | 128 (1.66) | 47 |
| Total | 7726 (100) | 311 |
| County | Temperature (°C) | Relative Humidity (%) | Total Precipitation (mm) | |||
|---|---|---|---|---|---|---|
| r | p | r | p | r | p | |
| Zagreb City | 0.2849 | 0.3694 | 0.0811 | 0.8021 | 0.6367 | 0.0260 |
| Zagreb County | 0.2897 | 0.3610 | 0.0356 | 0.9125 | 0.6493 | 0.0223 |
| Sisak-Moslavina | 0.4032 | 0.1938 | −0.1592 | 0.6211 | 0.3985 | 0.1995 |
| Osijek-Baranja | 0.2486 | 0.4359 | −0.1403 | 0.6637 | 0.4659 | 0.1269 |
| Brod-Posavina | 0.4849 | 0.1101 | −0.4535 | 0.1386 | −0.1763 | 0.5836 |
| Koprivnica-Križevci | 0.1474 | 0.6477 | −0.1270 | 0.6941 | 0.6192 | 0.0318 |
| Krapina-Zagorje | 0.1493 | 0.6433 | 0.0557 | 0.8636 | 0.5533 | 0.0620 |
| Bjelovar-Bilogora | 0.1477 | 0.6468 | 0.0059 | 0.9855 | 0.8496 | 0.0005 |
| Varaždin | 0.1098 | 0.7341 | 0.3033 | 0.3378 | 0.6076 | 0.0361 |
| Požega-Slavonia | 0.1839 | 0.5673 | 0.0466 | 0.8855 | −0.0546 | 0.8662 |
| Vukovar-Srijem | −0.0413 | 0.8985 | 0.2248 | 0.4823 | −0.0330 | 0.9190 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vilibić-Čavlek, T.; Barbić, L.; Klobučar, A.; Vucelja, M.; Bogdanić, M.; Sabadi, D.; Kutleša, M.; Gjurašin, B.; Stevanović, V.; Curman Posavec, M.; et al. Re-Emergence of Usutu Virus and Spreading of West Nile Virus Neuroinvasive Infections During the 2024 Transmission Season in Croatia. Viruses 2025, 17, 846. https://doi.org/10.3390/v17060846
Vilibić-Čavlek T, Barbić L, Klobučar A, Vucelja M, Bogdanić M, Sabadi D, Kutleša M, Gjurašin B, Stevanović V, Curman Posavec M, et al. Re-Emergence of Usutu Virus and Spreading of West Nile Virus Neuroinvasive Infections During the 2024 Transmission Season in Croatia. Viruses. 2025; 17(6):846. https://doi.org/10.3390/v17060846
Chicago/Turabian StyleVilibić-Čavlek, Tatjana, Ljubo Barbić, Ana Klobučar, Marko Vucelja, Maja Bogdanić, Dario Sabadi, Marko Kutleša, Branimir Gjurašin, Vladimir Stevanović, Marcela Curman Posavec, and et al. 2025. "Re-Emergence of Usutu Virus and Spreading of West Nile Virus Neuroinvasive Infections During the 2024 Transmission Season in Croatia" Viruses 17, no. 6: 846. https://doi.org/10.3390/v17060846
APA StyleVilibić-Čavlek, T., Barbić, L., Klobučar, A., Vucelja, M., Bogdanić, M., Sabadi, D., Kutleša, M., Gjurašin, B., Stevanović, V., Curman Posavec, M., Bjedov, L., Boljfetić, M., Jozić-Novinc, T., Škara, R., Tomljenović, M., Hruškar, Ž., Al-Mufleh, M., Potočnik-Hunjadi, T., Rončević, I., & Savić, V. (2025). Re-Emergence of Usutu Virus and Spreading of West Nile Virus Neuroinvasive Infections During the 2024 Transmission Season in Croatia. Viruses, 17(6), 846. https://doi.org/10.3390/v17060846