
Characterization of the Virome in Mosquitoes Across Distinct Habitats in the Yucatán Peninsula, Mexico

 ,
,  , , , , , , , , , , , , , and
, , , , , , , , , , , , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
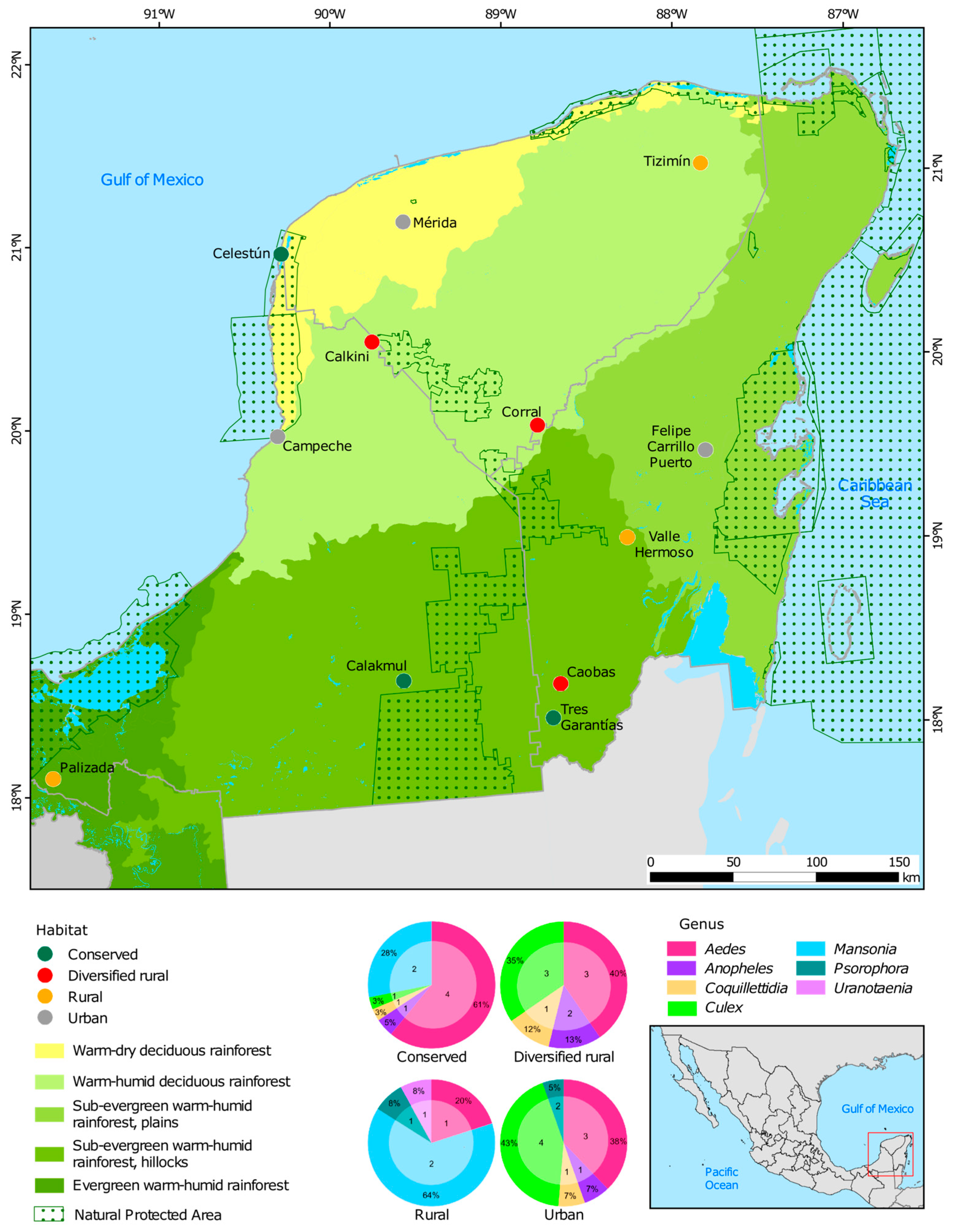
2.1. Site Study
2.2. Sampling Collection and Species Identification
2.3. Sampling Selection and Processing
2.4. Extraction of Genetic Material
2.5. Genomic Libraries and Massive Sequencing
2.6. Bioinformatic Analysis
2.7. Virus Abundance and Diversity Metrics
3. Results
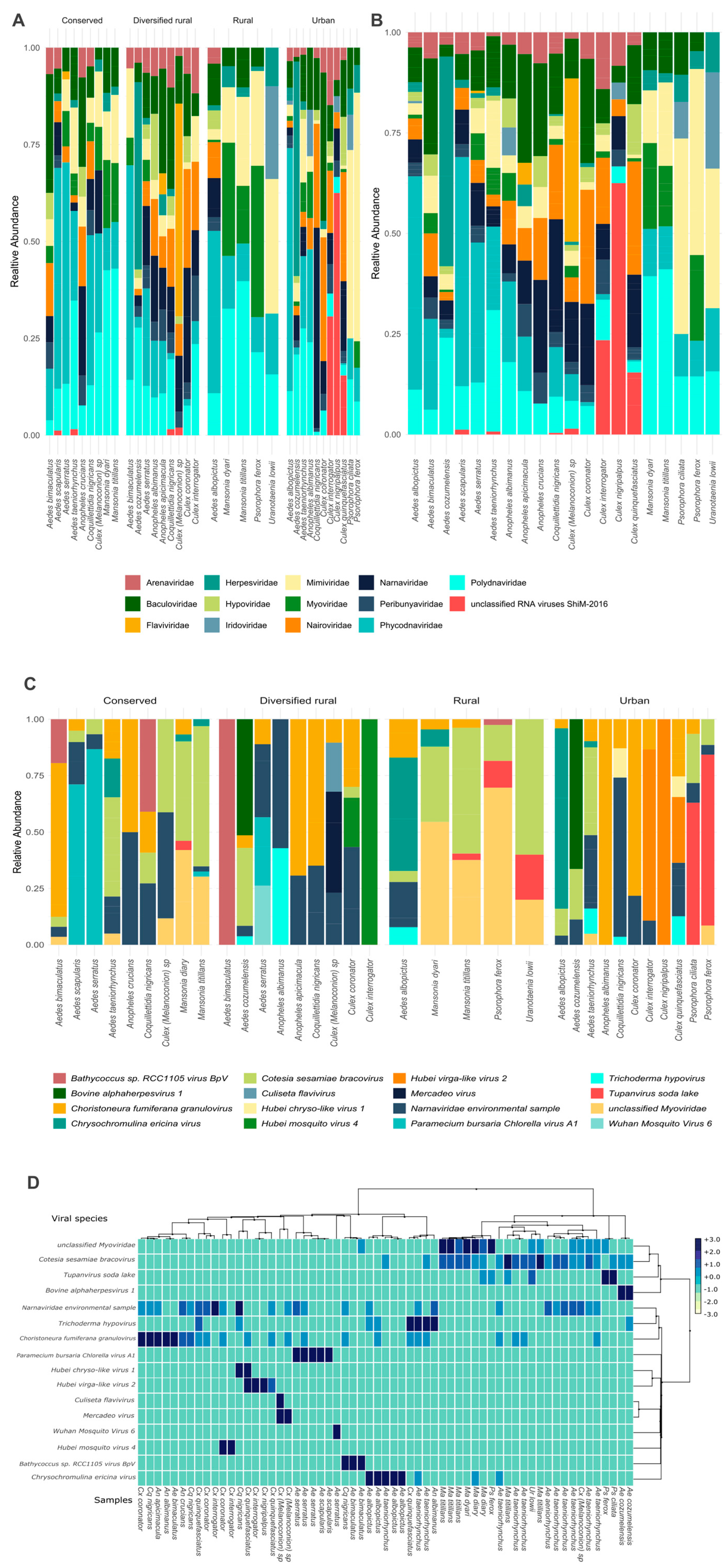
3.1. Diversity of the Viral Community
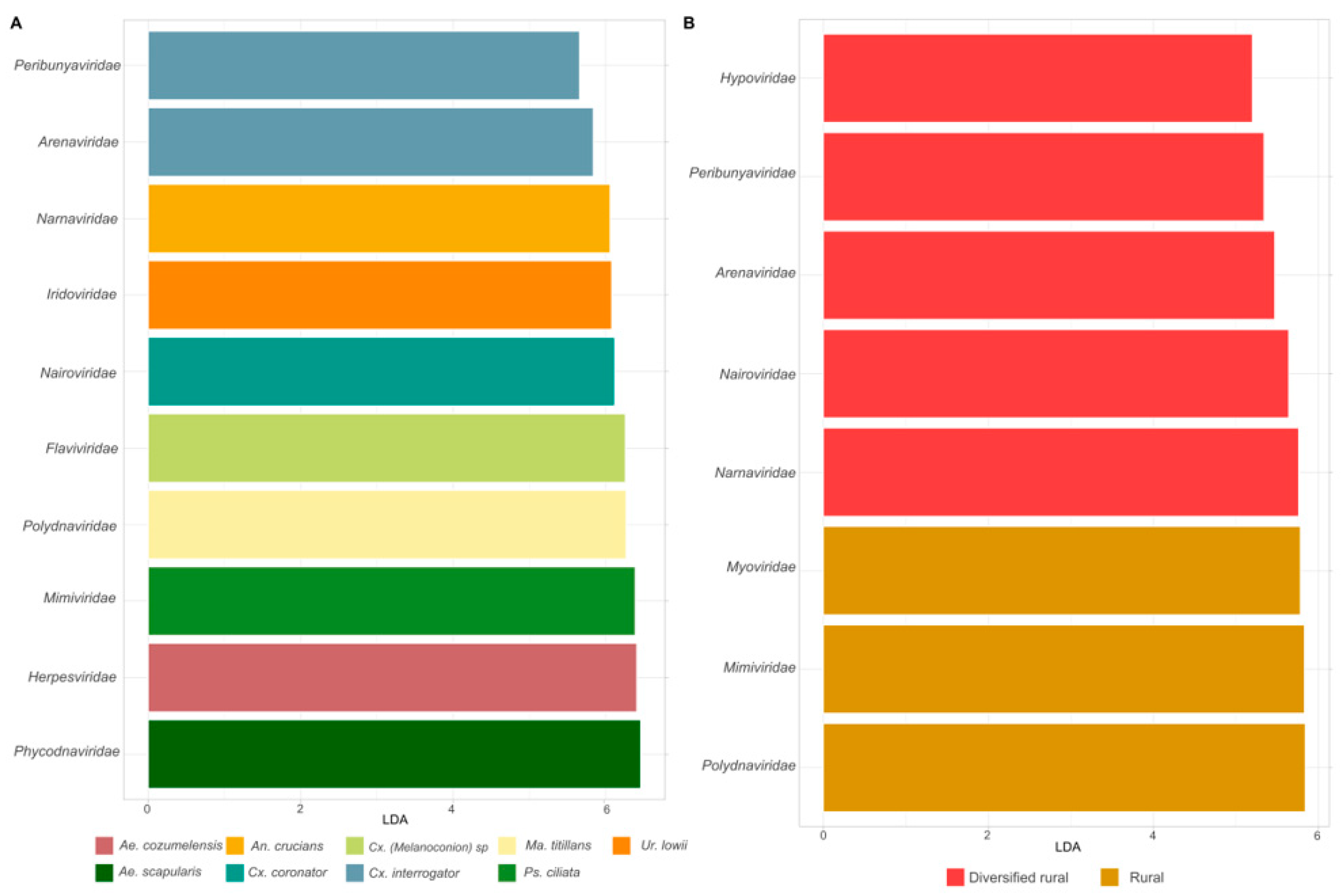
3.2. Differential Abundance of Viral Families
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| NGS | Next-generation sequencing |
| DENV | Dengue virus |
| CHIKV | Chikungunya virus |
| WNV | West Nile virus |
| ZIKV | Zika virus |
| ISVs | Insect-specific viruses |
| CFAV | Cell fusing agent virus |
| ISFVs | Insect-specific flaviviruses |
| vOTU | Viral operational taxonomic unit |
| LCA | Lowest Common Ancestor |
| PCoA | Principal Coordinate Analysis |
| PERMANOVA | Permutational analysis of variance |
| LDA | Linear discriminant analysis |
| LEfSe | Effect size |
| CxFV | Culex Flavivirus |
References
- de Oliveira Guimarães, L.; de Oliveira Ribeiro, G.; da Couto, R.; do Socorro Foro Ramos, E.; dos Santos Morais, V.; Telles-de-Deus, J.; Helfstein, V.C.; dos Santos, J.M.; Deng, X.; Delwart, E.; et al. Exploring mosquito virome dynamics within São Paulo Zoo: Insights into mosquito-virus-environment interactions. Front. Cell. Infect. Microbiol. 2025, 14, 1496126. [Google Scholar] [CrossRef]
- Hermanns, K.; Marklewitz, M.; Zirkel, F.; Kopp, A.; Kramer-Schadt, S.; Junglen, S. Mosquito community composition shapes virus prevalence patterns along anthropogenic disturbance gradients. eLife 2023, 12, e66550. [Google Scholar] [CrossRef] [PubMed]
- Rakotonirina, A.; Maquart, P.-O.; Flamand, C.; Sokha, C.; Boyer, S. Mosquito diversity (Diptera: Culicidae) and medical importance in four Cambodian forests. Parasites Vectors 2023, 16, 110. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.; Yu, X.; Wang, P.; Cheng, G. Arbovirus lifecycle in mosquito: Acquisition, propagation and transmission. Expert Rev. Mol. Med. 2019, 21, e1. [Google Scholar] [CrossRef]
- Kapuscinski, M.L.; Bergren, N.A.; Russell, B.J.; Lee, J.S.; Borland, E.M.; Hartman, D.A.; King, D.C.; Hughes, H.R.; Burkhalter, K.L.; Kading, R.C.; et al. Genomic characterization of 99 viruses from the bunyavirus families Nairoviridae, Peribunyaviridae, and Phenuiviridae, including 35 previously unsequenced viruses. PLoS Pathog. 2021, 17, e1009315. [Google Scholar] [CrossRef]
- Hegde, S.; Rasgon, J.L.; Hughes, G.L. The microbiome modulates arbovirus transmission in mosquitoes. Curr. Opin. Virol. 2015, 15, 97–102. [Google Scholar] [CrossRef]
- Roiz, D.; Pontifes, P.A.; Jourdain, F.; Diagne, C.; Leroy, B.; Vaissière, A.-C.; Tolsá-García, M.J.; Salles, J.-M.; Simard, F.; Courchamp, F. The rising global economic costs of invasive Aedes mosquitoes and Aedes-borne diseases. Sci. Total Environ. 2024, 933, 173054. [Google Scholar] [CrossRef]
- Gómez, M.; Martínez, D.; Páez-Triana, L.; Luna, N.; Salas, J.L.D.L.; Hernández, C.; Flórez, A.Z.; Muñoz, M.; Ramírez, J.D. Characterizing viral species in mosquitoes (Culicidae) in the Colombian Orinoco: Insights from a preliminary metagenomic study. Sci. Rep. 2023, 13, 22081. [Google Scholar] [CrossRef]
- Hall, M.; Tamïr, D. Killing Mosquitoes: Think before You Swat. In Mosquitopia: The Place of Pests in a Healthy World; Hall, M., Tamïr, D., Eds.; Routledge: New York, NY, USA, 2022. [Google Scholar]
- Reinhold, J.M.; Lahondère, C. Mosquitoes feeding on ectothermic hosts: From host seeking to pathogen transmission. Curr. Opin. Insect Sci. 2024, 66, 101273. [Google Scholar] [CrossRef]
- de Almeida, J.P.; Aguiar, E.R.; Armache, J.N.; Olmo, R.P.; Marques, J.T. The virome of vector mosquitoes. Curr. Opin. Virol. 2021, 49, 7–12. [Google Scholar] [CrossRef]
- Fortes Aragão, C.; Patroca da Silva, S.; Laís Sena do Nascimento, B.; Silva da Silva, F.; Pinto Nunes Neto, J.; Soares Pinheiro, V.C.; Ribeiro Cruz, A.C. Shotgun Metagenomic Sequencing Reveals Virome Composition of Mosquitoes from a Transition Ecosystem of North-Northeast Brazil. Genes 2023, 14, 1443. [Google Scholar] [CrossRef] [PubMed]
- Thongsripong, P.; Chandler, J.A.; Kittayapong, P.; Wilcox, B.A.; Kapan, D.D.; Bennett, S.N. Metagenomic shotgun sequencing reveals host species as an important driver of virome composition in mosquitoes. Sci. Rep. 2021, 11, 8448. [Google Scholar] [CrossRef] [PubMed]
- Chandler, J.A.; Liu, R.M.; Bennett, S.N. RNA shotgun metagenomic sequencing of northern California (USA) mosquitoes uncovers viruses, bacteria, and fungi. Front. Microbiol. 2015, 24, 185. [Google Scholar] [CrossRef]
- Wang, Y.; Lin, X.; Li, C.; Liu, G.; Wang, S.; Chen, M.; Wei, X.; Wen, H.; Tao, Z.; Xu, Y. Metagenomic sequencing reveals viral diversity of mosquitoes from Shandong Province, China. Microbiol. Spectr. 2024, 12, e0393223. [Google Scholar] [CrossRef]
- Margalho Braga, C.; Patroca da Silva, S.; Pinto Nunes Neto, J.; Barbosa de Almeida Medeiros, D.; Ribeiro Cruz, A.C.; Laís Sena do Nascimento, B.; Santana Pinheiro, L.R.; Carício Martins, L. Viral metagenomics of hematophagous insects collected in the Carajas mining complex, Pará State, Brazil. Acta Trop. 2025, 263, 107551. [Google Scholar] [CrossRef]
- Shi, C.; Beller, L.; Deboutte, W.; Yinda, K.C.; Delang, L.; Vega-Rúa, A.; Failloux, A.-B.; Matthijnssens, J. Stable distinct core eukaryotic viromes in different mosquito species from Guadeloupe, using single mosquito viral metagenomics. Microbiome 2019, 7, 121. [Google Scholar] [CrossRef]
- Potter-Birriel, J.M.; Pollio, A.R.; Knott, B.D.; Chunashvili, T.; Fung, C.K.; Conte, M.A.; Reinbold-Wasson, D.D.; Hang, J. Metagenomics analysis reveals presence of the Merida-like virus in Georgia. Front. Microbiol. 2023, 14, 1258810. [Google Scholar] [CrossRef]
- Ali, R.; Jayaraj, J.; Mohammed, A.; Chinnaraja, C.; Carrington, C.V.F.; Severson, D.W.; Ramsubhag, A. Characterization of the virome associated with Haemagogus mosquitoes in Trinidad, West Indies. Sci. Rep. 2021, 11, 16584. [Google Scholar] [CrossRef]
- Sadeghi, M.; Altan, E.; Deng, X.; Barker, C.M.; Fang, Y.; Coffey, L.L.; Delwart, E. Virome of >12 thousand Culex mosquitoes from throughout California. Virology 2018, 523, 74–88. [Google Scholar] [CrossRef]
- Frey, K.G.; Biser, T.; Hamilton, T.; Santos, C.J.; Pimentel, G.; Mokashi, V.P.; Bishop-Lilly, K.A. Bioinformatic Characterization of Mosquito Viromes within the Eastern United States and Puerto Rico: Discovery of Novel Viruses. Evol. Bioinform. 2016, 12, 1–12. [Google Scholar] [CrossRef]
- Gómez, M.; Martinez, D.; Muñoz, M.; Ramírez, J.D. Aedes aegypti and Ae. albopictus microbiome/virome: New strategies for controlling arboviral transmission? Parasites Vectors 2022, 15, 287. [Google Scholar] [CrossRef] [PubMed]
- Moonen, J.P.; Schinkel, M.; van der Most, T.; Miesen, P.; van Rij, R.P. Composition and global distribution of the mosquito virome—A comprehensive database of insect-specific viruses. One Health 2023, 16, 100490. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, V.L.; Long, M.T. Insect-Specific Viruses: An overview and their relationship to arboviruses of concern to humans and animals. Virology 2021, 557, 34–43. [Google Scholar] [CrossRef]
- de Faria, I.J.S.; de Almeida, J.P.P.; Marques, J.T. Impact of symbiotic insect-specific viruses on mosquito vector competence for arboviruses. Curr. Opin. Insect Sci. 2024, 63, 101194. [Google Scholar] [CrossRef]
- Agboli, E.; Leggewie, M.; Altinli, M.; Schnettler, E. Mosquito-Specific Viruses—Transmission and Interaction. Viruses 2019, 11, 873. [Google Scholar] [CrossRef]
- Altinli, M.; Schnettler, E.; Sicard, M. Symbiotic Interactions Between Mosquitoes and Mosquito Viruses. Front. Cell. Infect. Microbiol. 2021, 30, 694020. [Google Scholar] [CrossRef]
- Fujita, R.; Kato, F.; Kobayashi, D.; Murota, K.; Takasaki, T.; Tajima, S.; Lim, C.-K.; Saijo, M.; Isawa, H.; Sawabe, K. Persistent viruses in mosquito cultured cell line suppress multiplication of flaviviruses. Heliyon 2018, 4, e00736. [Google Scholar] [CrossRef]
- Öhlund, P.; Lundén, H.; Blomström, A.-L. Insect-specific virus evolution and potential effects on vector competence. Virus Genes 2019, 55, 127–137. [Google Scholar] [CrossRef]
- Olmo, R.P.; Todjro, Y.M.H.; Aguiar, E.R.G.R.; de Almeida, J.P.P.; Ferreira, F.V.; Armache, J.N.; de Faria, I.J.S.; Ferreira, A.G.A.; Amadou, S.C.G.; Silva, A.T.S.; et al. Mosquito vector competence for dengue is modulated by insect-specific viruses. Nat. Microbiol. 2023, 8, 135–149. [Google Scholar] [CrossRef]
- Peterson, A.J.; Hall, R.A.; Harrison, J.J.; Hobson-Peters, J.; Hugo, L.E. Unleashing Nature’s Allies: Comparing the Vertical Transmission Dynamics of Insect-Specific and Vertebrate-Infecting Flaviviruses in Mosquitoes. Viruses 2024, 16, 1499. [Google Scholar] [CrossRef]
- Patterson, E.I.; Villinger, J.; Muthoni, J.N.; Dobel-Ober, L.; Hughes, G.L. Exploiting insect-specific viruses as a novel strategy to control vector-borne disease. Curr. Opin. Insect Sci. 2020, 39, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Calzolari, M.; Zé-Zé, L.; Vázquez, A.; Paz Sánchez Seco, M.; Amaro, F.; Dottori, M. Insect-specific flaviviruses, a worldwide widespread group of viruses only detected in insects. Infect. Genet. Evol. 2016, 40, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Baidaliuk, A.; Miot, E.F.; Lequime, S.; Moltini-Conclois, I.; Delaigue, F.; Dabo, S.; Dickson, L.B.; Aubry, F.; Merkling, S.H.; Cao-Lormeau, V.-M.; et al. Cell-Fusing Agent Virus Reduces Arbovirus Dissemination in Aedes aegypti Mosquitoes In Vivo. J. Virol. 2019, 93, e00705-19. [Google Scholar] [CrossRef] [PubMed]
- Zhou, N.; Huang, E.; Guo, X.; Xiong, Y.; Xie, J.; Cai, T.; Du, Y.; Wu, Q.; Guo, S.; Han, W.; et al. Cell fusing agent virus isolated from Aag2 cells does not vertically transmit in Aedes aegypti via artificial infection. Parasites Vectors 2023, 16, 402. [Google Scholar] [CrossRef]
- Chen, J.; Deng, S.; Peng, H. Insect-specific viruses used in biocontrol of mosquito-borne diseases. Interdiscip. Med. 2023, 1, e20220001. [Google Scholar] [CrossRef]
- Lequime, S.; Lambrechts, L. Discovery of flavivirus-derived endogenous viral elements in Anopheles mosquito genomes supports the existence of Anopheles-associated insect-specific flaviviruses. Virus Evol. 2017, 3, vew035. [Google Scholar] [CrossRef]
- Liu, Q.; Cui, F.; Liu, X.; Fu, Y.; Fang, W.; Kang, X.; Lu, H.; Li, S.; Liu, B.; Guo, W.; et al. Association of virome dynamics with mosquito species and environmental factors. Microbiome 2023, 11, 101. [Google Scholar] [CrossRef]
- da Silva, H.; Oliveira, T.M.P.; Sallum, M.A.M. Bacterial Community Diversity and Bacterial Interaction Network in Eight Mosquito Species. Genes 2022, 13, 2052. [Google Scholar] [CrossRef]
- Nebbak, A.; Monteil-Bouchard, S.; Berenger, J.-M.; Almeras, L.; Parola, P.; Desnues, C. Virome Diversity among Mosquito Populations in a Sub-Urban Region of Marseille, France. Viruses 2021, 13, 768. [Google Scholar] [CrossRef]
- Wilke, A.B.; Benelli, G.; Beier, J.C. Anthropogenic changes and associated impacts on vector-borne diseases. Trends Parasitol. 2021, 37, 1027–1030. [Google Scholar] [CrossRef]
- Perrin, A.; Glaizot, O.; Christe, P. Worldwide impacts of landscape anthropization on mosquito abundance and diversity: A meta-analysis. Glob. Change Biol. 2022, 28, 6857–6871. [Google Scholar] [CrossRef] [PubMed]
- Ren, N.; Wang, S.; Shi, C.; Yu, P.; Zhao, L.; Huang, D.; Ma, H.; Xiao, S.; Wang, F.; Yuan, Z.; et al. Dynamic Surveillance of Mosquitoes and Their Viromes in Wuhan During 2020. Zoonoses 2021, 1. [Google Scholar] [CrossRef]
- Khan, S.; Abel, M.G.; Bibbs, C.S.; Faraji, A.; Chaves, L.F. Mosquito (Diptera: Culicidae) species diversity and abundance patterns across tree height and microclimatic gradients in Indiana, USA. Biologia 2024, 79, 2433–2443. [Google Scholar] [CrossRef]
- García-Suárez, O.; Tolsá-García, M.; Arana-Guardia, R.; Rodríguez-Valencia, V.; Talaga, S.; Pontifes, P.; Machain-Williams, C.; Suzán, G.; Roiz, D. Seasonal mosquito (Diptera: Culicidae) dynamics and the influence of environmental variables in a land use gradient from Yucatan, Mexico. Acta Trop. 2024, 257, 107275. [Google Scholar] [CrossRef]
- Galaz, V.; Rocha, J.; Sánchez-García, P.A.; Dauriach, A.; Roukny, T.; Jørgensen, P.S. Financial influence on global risks of zoonotic emerging and re-emerging diseases: An integrative analysis. Lancet Planet. Health 2023, 7, e951–e962. [Google Scholar] [CrossRef]
- Swaddle, J.P.; Calos, S.E. Increased Avian Diversity Is Associated with Lower Incidence of Human West Nile Infection: Observation of the Dilution Effect. PLoS ONE 2008, 3, e2488. [Google Scholar] [CrossRef]
- National Research Council (USA); Avise, J.C.; Hubbell, S.P.; Ayala, F.J. Homage to Linnaeus: How Many Parasites? How Many Hosts? National Academies Press: Washington, DC, USA, 2008. [Google Scholar]
- Huang, Z.Y.X.; VAN Langevelde, F.; Estrada-Peña, A.; Suzán, G.; DE Boer, W.F. The diversity–disease relationship: Evidence for and criticisms of the dilution effect. Parasitology 2016, 143, 1075–1086. [Google Scholar] [CrossRef]
- Talaga, S.; le Goff, G.; Arana-Guardia, R.; Baak-Baak, C.M.; García-Rejón, J.E.; García-Suárez, O.; Rodríguez-Valencia, V.M.; Tolsá-García, M.J.; Suzán, G.; Roiz, D. The mosquitoes (Diptera: Culicidae) of the Mexican Yucatan Peninsula: A comprehensive review on the use of taxonomic names. J. Med. Èntomol. 2023, 61, 274–308. [Google Scholar] [CrossRef]
- Sánchez-Soto, M.F.; Gaona, O.; Mercado-Juárez, R.; Yanez-Montalvo, A.; de León-Lorenzana, A.; Borja-Martínez, G.; Zaldívar, D.; Rodríguez-González, S.; Hernández-Villegas, E.N.; Moreira-Soto, A.; et al. Ecological-based insights into bat populations in the Yucatán Peninsula under a One Health approach: Coexistence or biophobia. Front. Conserv. Sci. 2025, 5, 1488378. [Google Scholar] [CrossRef]
- Falcón-Brindis, A.; León-Cortés, J.L.; Montañez-Reyna, M. How effective are conservation areas to preserve biodiversity in Mexico? Perspect. Ecol. Conserv. 2021, 19, 399–410. [Google Scholar] [CrossRef]
- Haro, P.; Trasviña-Muñoz, E.; May-Concha, I.; López-Valencia, G.; Monge-Navarro, F.; Herrera-Ramírez, C.; Mercado-Rodríguez, J.A.; Villanueva-Alonzo, H.; Waleckx, E. Historical Spatial Distribution of Zoonotic Diseases in Domestic, Synanthropic, and Wild Animals in the Mexican Territory of the Yucatan Peninsula. J. Trop. Med. 2021, 2021, 8699455. [Google Scholar] [CrossRef] [PubMed]
- Marcial Baak-Baak, C.; Cigarroa-Toledo, N.; Pech-May, A.; Cruz-Escalona, G.A.; Cetina-Trejo, R.C.; Tzuc-Dzul, J.C.; Talavera-Aguilar, L.G.; Flores-Ruiz, S.; Machain-Williams, C.; Torres-Chable, O.M.; et al. Entomological and virological surveillance for dengue virus in churches in Merida, Mexico. Rev. Inst. Med. Trop. Sao Paulo 2019, 61, e9. [Google Scholar] [CrossRef] [PubMed]
- Chu, Y.-K.; Owen, R.D.; Sánchez-Hernández, C.; de Lourdes Romero-Almaraz, M.; Jonsson, C.B. Genetic characterization and phylogeny of a hantavirus from Western Mexico. Virus Res. 2008, 131, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Chaves, A.; Sotomayor-Bonilla, J.; Monge, O.; Ramírez, A.; Galindo, F.; Sarmiento-Silva, R.E.; Gutiérrez-Espeleta, G.A.; Suzán, G. West Nile Virus in Resident Birds from Yucatan, Mexico. J. Wildl. Dis. 2016, 52, 159–163. [Google Scholar] [CrossRef]
- Sotomayor-Bonilla, J.; Chaves, A.; Rico-Chávez, O.; Rostal, M.K.; Ojeda-Flores, R.; Salas-Rojas, M.; Aguilar-Setien, Á.; Ibáñez-Bernal, S.; Barbachano-Guerrero, A.; Gutiérrez-Espeleta, G.; et al. Dengue Virus in Bats from Southeastern Mexico. Am. J. Trop. Med. Hyg. 2014, 91, 129–131. [Google Scholar] [CrossRef]
- Farfan-Ale, J.A.; Loroño-Pino, M.A.; Garcia-Rejon, J.E.; Hovav, E.; Powers, A.M.; Lin, M.; Dorman, K.S.; Platt, K.B.; Bartholomay, L.C.; Soto, V.; et al. Detection of RNA from a Novel West Nile-like Virus and High Prevalence of an Insect-specific Flavivirus in Mosquitoes in the Yucatan Peninsula of Mexico. Am. J. Trop. Med. Hyg. 2009, 80, 85–95. [Google Scholar] [CrossRef]
- Charles, J.; Firth, A.E.; Loroño-Pino, M.A.; Garcia-Rejon, J.E.; Farfan-Ale, J.A.; Lipkin, W.I.; Blitvich, B.J.; Briese, T. Merida virus, a putative novel rhabdovirus discovered in Culex and Ochlerotatus spp. mosquitoes in the Yucatan Peninsula of Mexico. J. Gen. Virol. 2016, 97, 977–987. [Google Scholar] [CrossRef] [PubMed]
- Charles, J.; Tangudu, C.S.; Hurt, S.L.; Tumescheit, C.; Firth, A.E.; Garcia-Rejon, J.E.; Machain-Williams, C.; Blitvich, B.J. Detection of novel and recognized RNA viruses in mosquitoes from the Yucatan Peninsula of Mexico using metagenomics and characterization of their in vitro host ranges. J. Gen. Virol. 2018, 99, 1729–1738. [Google Scholar] [CrossRef]
- Briese, T.; Loroño-Pino, M.A.; Garcia-Rejon, J.E.; Farfan-Ale, J.A.; Machain-Williams, C.; Dorman, K.S.; Lipkin, W.I.; Blitvich, B.J. Complete genome sequence of T’Ho virus, a novel putative flavivirus from the Yucatan Peninsula of Mexico. Virol. J. 2017, 14, 110. [Google Scholar] [CrossRef]
- ArcGIS Desktop 10.6.1. Available online: https://desktop.arcgis.com/en/quick-start-guides/10.6/arcgis-desktop-quick-start-guide.htm (accessed on 10 May 2025).
- Andrews, S. FastQC: A Quality Control Analysis Tool for High Throughput Sequencing Data; Github: San Francisco, CA, USA, 2024. [Google Scholar]
- Krueger, F. TrimGalore: A Wrapper around Cutadapt and FastQC to Consistently Apply Adapter and Quality Trimming to FastQ Files, with Extra Functionality for RRBS Data; Github: San Francisco, CA, USA, 2025. [Google Scholar]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Nurk, S.; Meleshko, D.; Korobeynikov, A.; Pevzner, P.A. metaSPAdes: A new versatile metagenomic assembler. Genome Res. 2017, 27, 824–834. [Google Scholar] [CrossRef] [PubMed]
- Castelán-Sánchez, H.G.; Lopéz-Rosas, I.; García-Suastegui, W.A.; Peralta, R.; Dobson, A.D.; Batista-García, R.A.; Dávila-Ramos, S. Extremophile deep-sea viral communities from hydrothermal vents: Structural and functional analysis. Mar. Genom. 2019, 46, 16–28. [Google Scholar] [CrossRef] [PubMed]
- Huson, D.H.; Auch, A.F.; Qi, J.; Schuster, S.C. MEGAN analysis of metagenomic data. Genome Res. 2007, 17, 377–386. [Google Scholar] [CrossRef]
- Valero-Mora, P.M. ggplot2: Elegant graphics for data analysis. J. Stat. Softw. 2010, 35, 1–3. [Google Scholar] [CrossRef]
- Oksanen, J.; Kindt, R.; Legendre, P.; O’Hara, B. The Vegan Package. Available online: https://www.researchgate.net/publication/228975085_The_Vegan_Package (accessed on 10 May 2025).
- Kassambara, A. Ggpubr: “ggplot2” Based Publication Ready Plots, 0.6.0; Github: San Francisco, CA, USA, 2023. [Google Scholar]
- Orme, D.; Villeger, S.; De Bortoli, J.; Leprieur, F.; Logez, M. Betapart: Partitioning Beta Diversity into Turnover and Nestedness Components. R Package Version 2018. Available online: https://cran.r-project.org/web/packages/betapart/betapart.pdf (accessed on 10 May 2025).
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2008, 26, 32–46. [Google Scholar] [CrossRef]
- The Microbial Package for Microbial Community Analysis. Available online: https://rdrr.io/cran/microbial/f/vignettes/microbial.Rmd (accessed on 3 May 2024).
- Shi, M.; Lin, X.-D.; Tian, J.-H.; Chen, L.-J.; Chen, X.; Li, C.-X.; Qin, X.-C.; Li, J.; Cao, J.-P.; Eden, J.-S.; et al. Redefining the invertebrate RNA virosphere. Nature 2016, 540, 539–543. [Google Scholar] [CrossRef]
- Xiao, P.; Li, C.; Zhang, Y.; Han, J.; Guo, X.; Xie, L.; Tian, M.; Li, Y.; Wang, M.; Liu, H.; et al. Metagenomic Sequencing from Mosquitoes in China Reveals a Variety of Insect and Human Viruses. Front. Cell. Infect. Microbiol. 2018, 8, 364. [Google Scholar] [CrossRef]
- Atoni, E.; Wang, Y.; Karungu, S.; Waruhiu, C.; Zohaib, A.; Obanda, V.; Agwanda, B.; Mutua, M.; Xia, H.; Yuan, Z. Metagenomic Virome Analysis of Culex Mosquitoes from Kenya and China. Viruses 2018, 10, 30. [Google Scholar] [CrossRef]
- Tokarz-Deptuła, B.; Niedźwiedzka-Rystwej, P.; Czupryńska, P.; Deptuła, W. Protozoal giant viruses: Agents potentially infectious to humans and animals. Virus Genes 2019, 55, 574–591. [Google Scholar] [CrossRef]
- Zheng, L.; Liang, X.; Shi, R.; Li, P.; Zhao, J.; Li, G.; Wang, S.; Han, S.; Radosevich, M.; Zhang, Y. Viral Abundance and Diversity of Production Fluids in Oil Reservoirs. Microorganisms 2020, 8, 1429. [Google Scholar] [CrossRef] [PubMed]
- Bah, A.; Lucarotti, C.J.; Arella, M.; Guertin, C. Choristoneura fumiferana granulovirus: Sequence analysis and 5′ characterization of ORF891. Arch. Virol. 1999, 144, 737–746. [Google Scholar] [CrossRef]
- Escasa, S.R.; Lauzon, H.A.M.; Mathur, A.C.; Krell, P.J.; Arif, B.M. Sequence analysis of the Choristoneura occidentalis granulovirus genome. J. Gen. Virol. 2006, 87, 1917–1933. [Google Scholar] [CrossRef] [PubMed]
- Gangopadhayya, A.; Lole, K.; Ghuge, O.; Ramdasi, A.; Kamble, A.; Roy, D.; Thakar, S.; Nath, A.; Sudeep, A.; Cherian, S. Metagenomic Analysis of Viromes of Aedes Mosquitoes across India. Viruses 2024, 16, 109. [Google Scholar] [CrossRef]
- Chersoni, L.; Checcucci, A.; Malfacini, M.; Puggioli, A.; Balestrino, F.; Carrieri, M.; Piunti, I.; Dindo, M.L.; Mattarelli, P.; Bellini, R. The Possible Role of Microorganisms in Mosquito Mass Rearing. Insects 2021, 12, 645. [Google Scholar] [CrossRef]
- Gonzalez Da Silva, A.; Bach, E.; Ellwanger, J.H.; Bogo Chies, J.A. Tips and tools to obtain and assess mosquito viromes. Arch. Microbiol. 2024, 206, 132. [Google Scholar] [CrossRef]
- Mwanga, E.P.; Mchola, I.S.; Makala, F.E.; Mshani, I.H.; Siria, D.J.; Mwinyi, S.H.; Abbasi, S.; Seleman, G.; Mgaya, J.N.; Jiménez, M.G.; et al. Rapid assessment of the blood-feeding histories of wild-caught malaria mosquitoes using mid-infrared spectroscopy and machine learning. Malar. J. 2024, 23, 86. [Google Scholar] [CrossRef]
- He, W.; Chen, Y.; Zhang, X.; Peng, M.; Xu, D.; He, H.; Gao, Y.; Chen, J.; Zhang, J.; Li, Z.; et al. Virome in adult Aedes albopictus captured during different seasons in Guangzhou City, China. Parasites Vectors 2021, 14, 415. [Google Scholar] [CrossRef]
- Ascoli, V.; Facchinelli, L.; Valerio, L.; Zucchetto, A.; Maso, L.D.; Coluzzi, M. Distribution of mosquito species in areas with high and low incidence of classic Kaposi’s sarcoma and seroprevalence for HHV-8. Med. Vet. Èntomol. 2006, 20, 198–208. [Google Scholar] [CrossRef]
- Gilden, D.H.; Mahalingam, R.; Cohrs, R.J.; Tyler, K.L. Herpesvirus infections of the nervous system. Nat. Clin. Pract. Neurol. 2007, 3, 82–94. [Google Scholar] [CrossRef]
- Schneider, C.A.; Calvo, E.; Peterson, K.E. Arboviruses: How Saliva Impacts the Journey from Vector to Host. Int. J. Mol. Sci. 2021, 22, 9173. [Google Scholar] [CrossRef] [PubMed]
- Carrera, J.-P.; Guzman, H.; Beltrán, D.; Díaz, Y.; López-Vergès, S.; Torres-Cosme, R.; Popov, V.; Widen, S.G.; Wood, T.G.; Weaver, S.C.; et al. Mercadeo Virus: A Novel Mosquito-Specific Flavivirus from Panama. Am. J. Trop. Med. Hyg. 2015, 93, 1014–1019. [Google Scholar] [CrossRef] [PubMed]
- Miranda, J.; Mattar, S.; Gonzalez, M.; Hoyos-López, R.; Aleman, A.; Aponte, J. First report of Culex flavivirus infection from Culex coronator (Diptera: Culicidae), Colombia. Virol. J. 2019, 16, 1. [Google Scholar] [CrossRef]
- Konstantinidis, K.; Bampali, M.; de Courcy Williams, M.; Dovrolis, N.; Gatzidou, E.; Papazilakis, P.; Nearchou, A.; Veletza, S.; Karakasiliotis, I. Dissecting the Species-Specific Virome in Culicoides of Thrace. Front. Microbiol. 2022, 13, 802577. [Google Scholar] [CrossRef]
- De Coninck, L.; Matthijnssens, J. The mosquito core virome: Beyond the buzz. Trends Parasitol. 2024, 40, 201–202. [Google Scholar] [CrossRef]
- Koh, C.; Saleh, M.-C. Mosquito core viromes: Do they exist? Trends Parasitol. 2024, 40, 203–204. [Google Scholar] [CrossRef]
- Torres, R.; Samudio, R.; Carrera, J.-P.; Young, J.; Márquez, R.; Hurtado, L.; Weaver, S.; Chaves, L.F.; Tesh, R.; Cáceres, L. Enzootic mosquito vector species at equine encephalitis transmission foci in the República de Panamá. PLoS ONE 2017, 12, e0185491. [Google Scholar] [CrossRef]
- Li, C.-X.; Shi, M.; Tian, J.-H.; Lin, X.-D.; Kang, Y.-J.; Chen, L.-J.; Qin, X.-C.; Xu, J.; Holmes, E.C.; Zhang, Y.-Z. Unprecedented genomic diversity of RNA viruses in arthropods reveals the ancestry of negative-sense RNA viruses. eLife 2015, 4, e05378. [Google Scholar] [CrossRef]
- De Coninck, L.; Soto, A.; Wang, L.; De Wolf, K.; Smitz, N.; Deblauwe, I.; Donfack, K.C.M.; Müller, R.; Delang, L.; Matthijnssens, J. Lack of abundant core virome in Culex mosquitoes from a temperate climate region despite a mosquito species-specific virome. mSystems 2024, 9, e0001224. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hernández-Villegas, E.N.; Castelán-Sánchez, H.G.; Moreira-Soto, A.; Vigueras-Galván, A.L.; Jiménez-Rico, M.A.; Rico-Chávez, O.; Rodríguez-González, S.; Tolsá-García, M.J.; Roiz, D.; Martínez-Duque, P.; et al. Characterization of the Virome in Mosquitoes Across Distinct Habitats in the Yucatán Peninsula, Mexico. Viruses 2025, 17, 758. https://doi.org/10.3390/v17060758
Hernández-Villegas EN, Castelán-Sánchez HG, Moreira-Soto A, Vigueras-Galván AL, Jiménez-Rico MA, Rico-Chávez O, Rodríguez-González S, Tolsá-García MJ, Roiz D, Martínez-Duque P, et al. Characterization of the Virome in Mosquitoes Across Distinct Habitats in the Yucatán Peninsula, Mexico. Viruses. 2025; 17(6):758. https://doi.org/10.3390/v17060758
Chicago/Turabian StyleHernández-Villegas, Erika N., Hugo G. Castelán-Sánchez, Andres Moreira-Soto, Ana Laura Vigueras-Galván, Marco A. Jiménez-Rico, Oscar Rico-Chávez, Stephany Rodríguez-González, María José Tolsá-García, David Roiz, Paola Martínez-Duque, and et al. 2025. "Characterization of the Virome in Mosquitoes Across Distinct Habitats in the Yucatán Peninsula, Mexico" Viruses 17, no. 6: 758. https://doi.org/10.3390/v17060758
APA StyleHernández-Villegas, E. N., Castelán-Sánchez, H. G., Moreira-Soto, A., Vigueras-Galván, A. L., Jiménez-Rico, M. A., Rico-Chávez, O., Rodríguez-González, S., Tolsá-García, M. J., Roiz, D., Martínez-Duque, P., Arana-Guardía, R., García-Súarez, O., Jiménez, M. Z., Falcón, L. I., Roche, B., Sarmiento-Silva, R. E., Arnal, A., Drexler, J. F., & Suzán, G. (2025). Characterization of the Virome in Mosquitoes Across Distinct Habitats in the Yucatán Peninsula, Mexico. Viruses, 17(6), 758. https://doi.org/10.3390/v17060758