
Development of a Noninfectious Japanese Encephalitis Virus Replicon for Antiviral Drug Screening and Gene Function Studies

 , , ,
, , ,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Cells, Virus, and Antibodies
2.2. Construction of the JEV-GFP Replicon Expressing Green Fluorescent Protein (GFP)
2.3. Transfection of the JEV-GFP Replicon
2.4. Western Blot, Immunofluorescence (IFA), and qRT-PCR Analyses
2.5. Antiviral Drug Screening
2.6. Cytotoxicity Assay
2.7. Analysis of the Inhibitory Effect of Antiviral Drugs
2.8. Statistical Analysis
3. Results
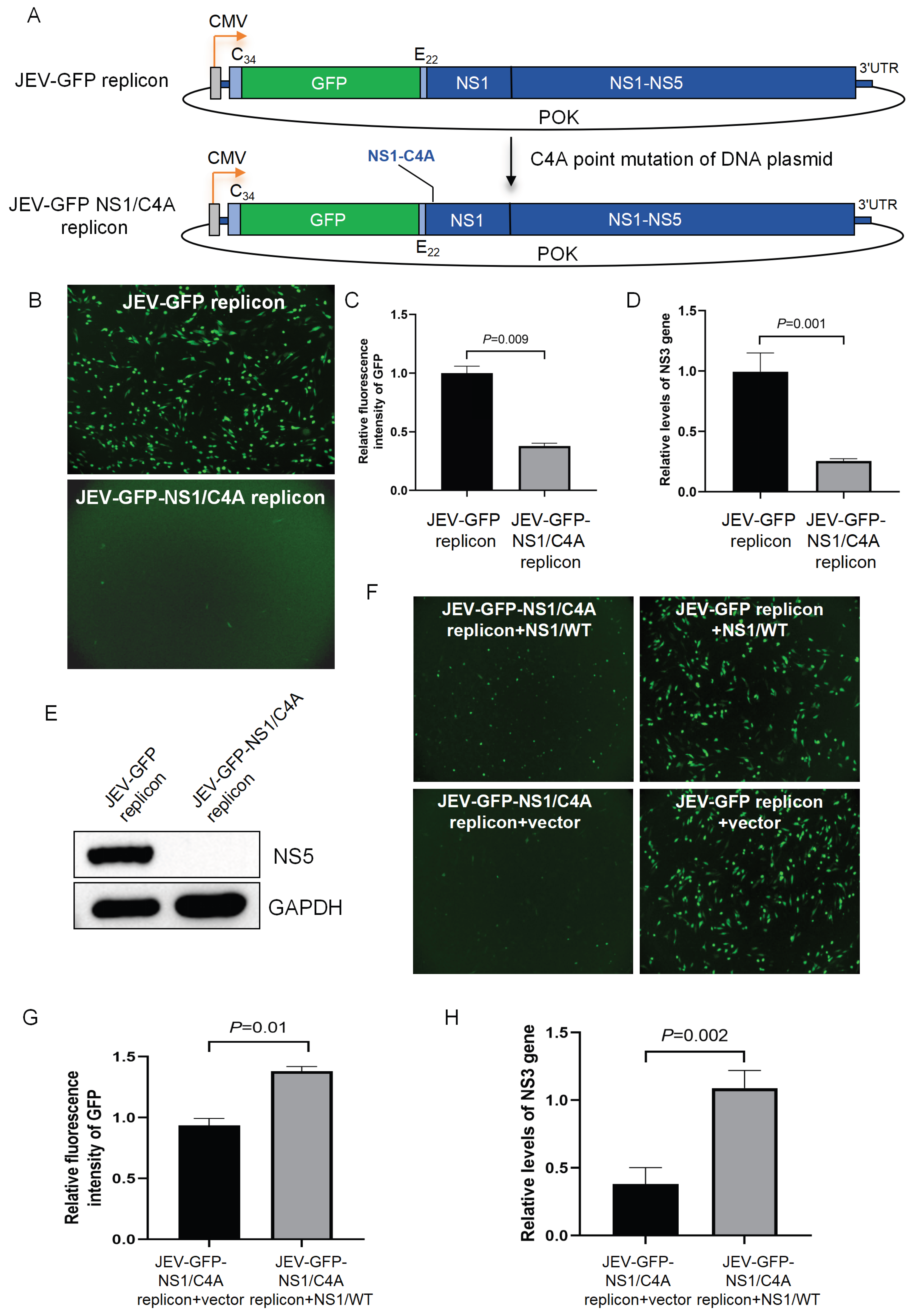
3.1. Construction of the JEV-GFP Replicon
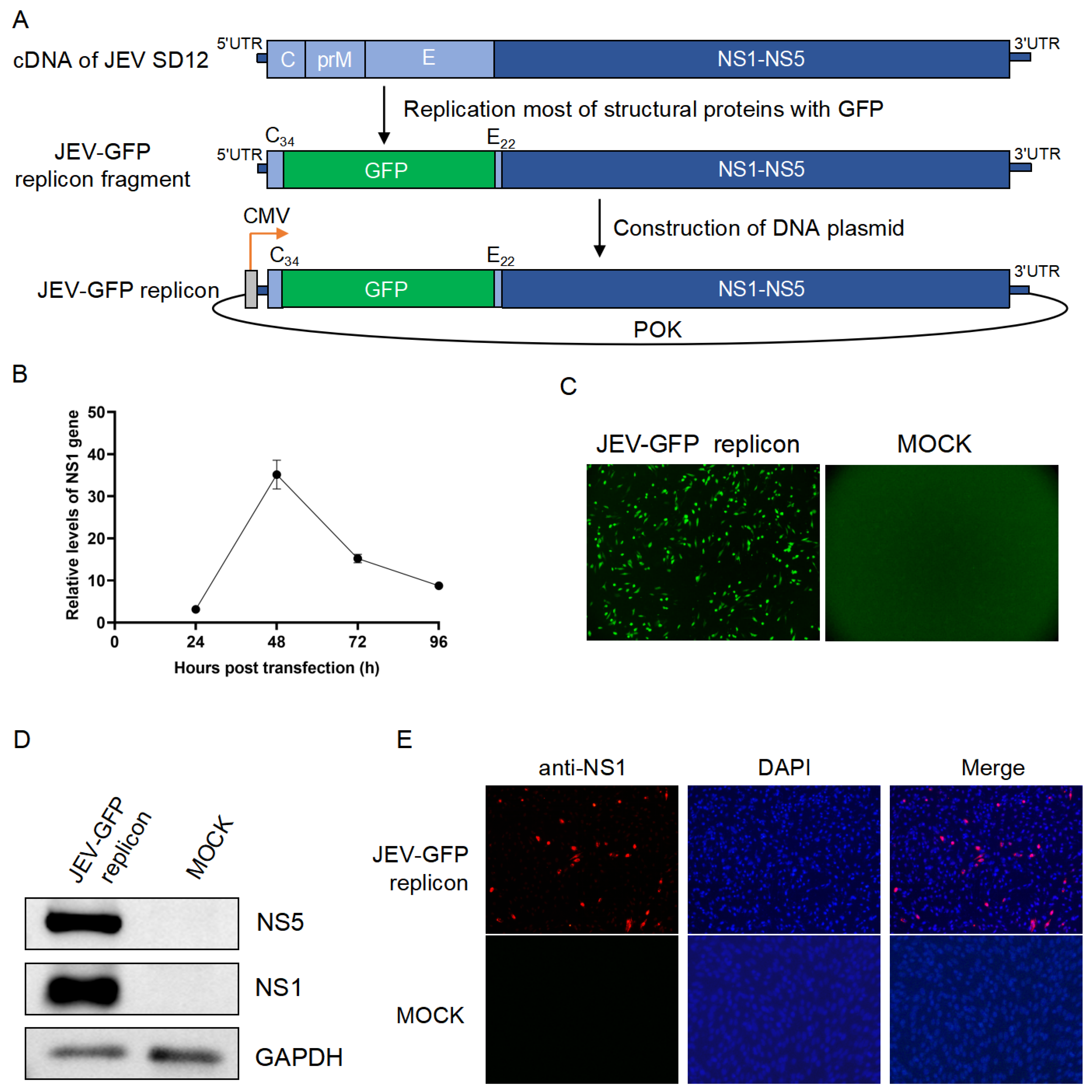
3.2. The JEV-GFP Replicon Used for Antiviral Drug Screening
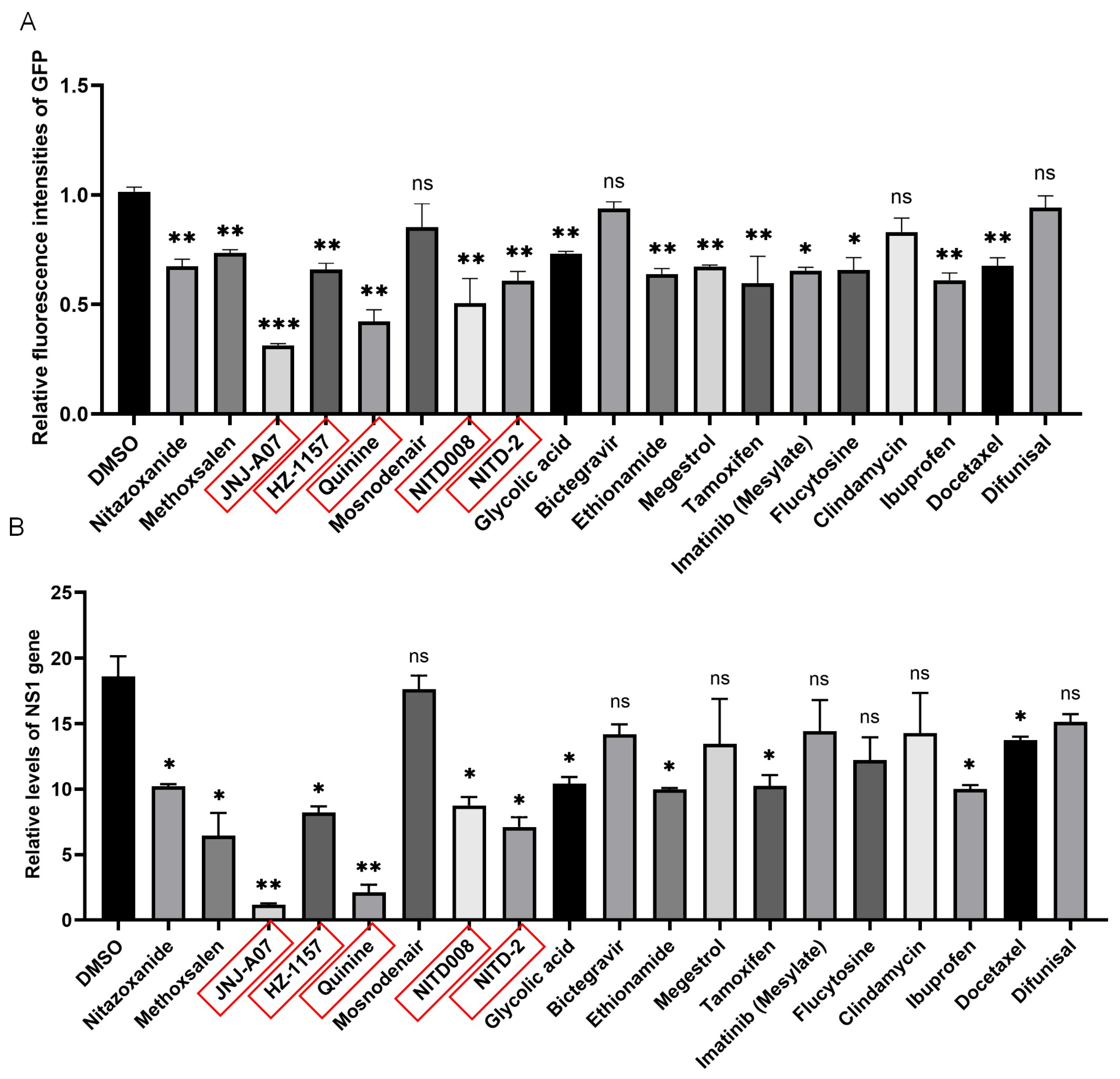

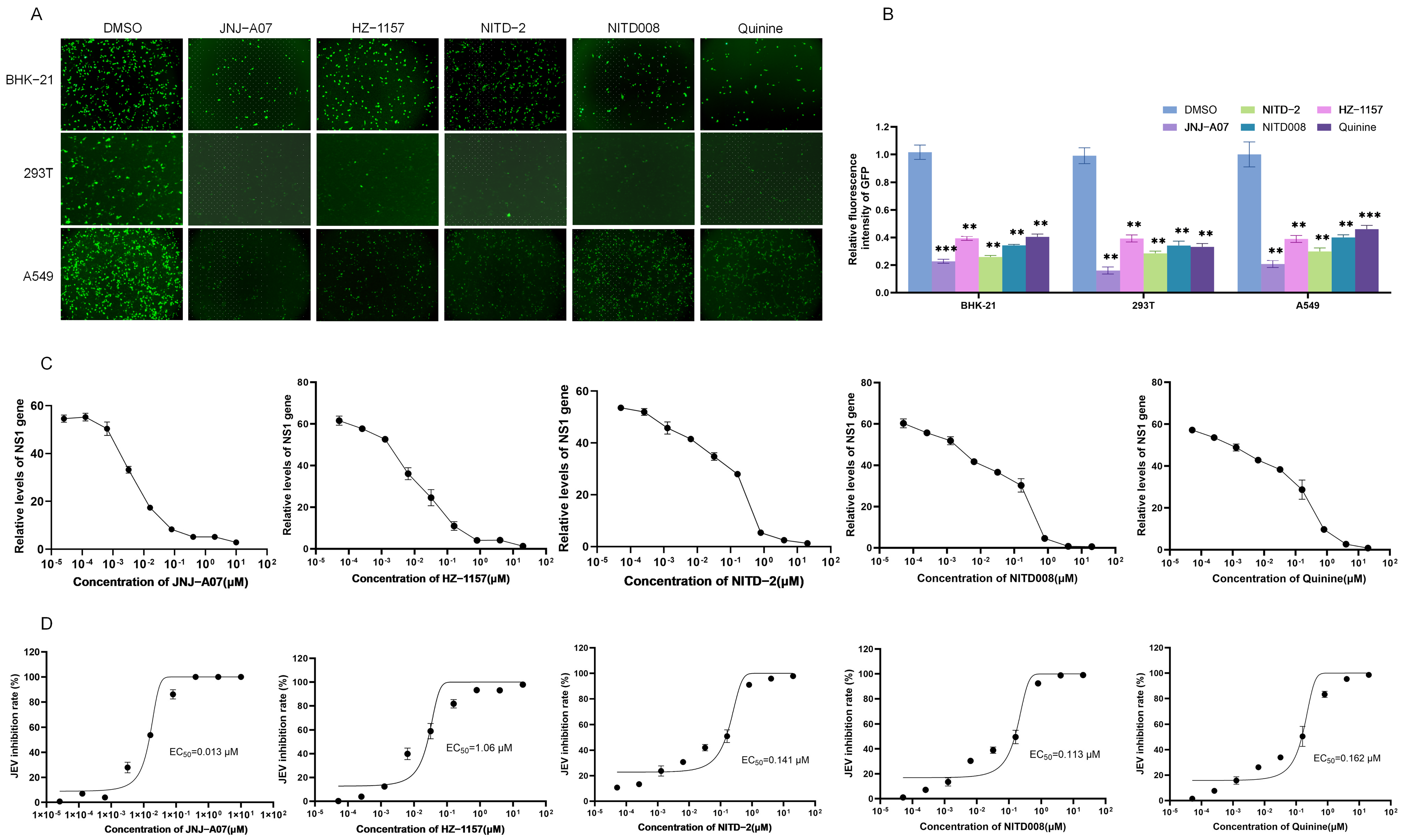
3.3. Antiviral Effect of JNJ-A07, HZ-1157, NITD-2, Quinine, and NITD008 on JEV Replication In Vitro
3.4. The JEV-GFP Replicon Used for the Identification of the NS1 Residue Required for Viral RNA Replication
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Huang, Y.; Liu, S.; Yang, P.; Wang, C.; Du, Y.; Yu, W.; Sun, Z. Replicon-based Japanese encephalitis virus vaccines elicit immune response in mice. J. Virol. Methods 2012, 179, 217–225. [Google Scholar] [CrossRef]
- Xu, J.; Wahaab, A.; Khan, S.; Nawaz, M.; Anwar, M.N.; Liu, K.; Wei, J.; Hameed, M.; Ma, Z. Recent Population Dynamics of Japanese Encephalitis Virus. Viruses 2023, 15, 1312. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.Y.; Li, X.D.; Liu, S.Q.; Deng, C.L.; Zhang, B.; Ye, H.Q. Development of a stable Japanese encephalitis virus replicon cell line for antiviral screening. Arch. Virol. 2017, 162, 3417–3423. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.Y.; Liu, S.Q.; Li, X.D.; Li, J.Q.; Zhang, Y.N.; Deng, C.L.; Zhang, H.L.; Li, X.F.; Fang, C.X.; Yang, F.X.; et al. Sequence duplication in 3’ UTR modulates virus replication and virulence of Japanese encephalitis virus. Emerg. Microbes Infect. 2022, 11, 123–135. [Google Scholar] [CrossRef]
- Liang, Z.J.; Pan, J.H.; Xie, S.D.; Yang, X.M.; Cao, R.B. Interaction between hTIM-1 and Envelope Protein Is Important for JEV Infection. Viruses 2023, 15, 1589. [Google Scholar] [CrossRef]
- Huang, R.; He, Y.; Zhang, C.; Luo, Y.; Chen, C.; Tan, N.; Ren, Y.; Xu, K.; Yuan, L.; Yang, J. The mutation of Japanese encephalitis virus envelope protein residue 389 attenuates viral neuroinvasiveness. Virol. J. 2024, 21, 128. [Google Scholar] [CrossRef]
- Hegde, N.R.; Gore, M.M. Japanese encephalitis vaccines: Immunogenicity, protective efficacy, effectiveness, and impact on the burden of disease. Hum. Vaccin. Immunother. 2017, 13, 1–18. [Google Scholar] [CrossRef]
- Hills, S.L.; Netravathi, M.; Solomon, T. Japanese Encephalitis among Adults: A Review. Am. J. Trop. Med. Hyg. 2023, 108, 860–864. [Google Scholar] [CrossRef]
- Wahaab, A.; Mustafa, B.E.; Hameed, M.; Stevenson, N.J.; Anwar, M.N.; Liu, K.; Wei, J.; Qiu, Y.; Ma, Z. Potential Role of Flavivirus NS2B-NS3 Proteases in Viral Pathogenesis and Anti-flavivirus Drug Discovery Employing Animal Cells and Models: A Review. Viruses 2021, 14, 44. [Google Scholar] [CrossRef]
- van den Elsen, K.; Chew, B.L.A.; Ho, J.S.; Luo, D.H. Flavivirus nonstructural proteins and replication complexes as antiviral drug targets. Curr. Opin. Virol. 2023, 59, 101305. [Google Scholar] [CrossRef]
- Duan, Y.; Zeng, M.; Jiang, B.; Zhang, W.; Wang, M.; Jia, R.; Zhu, D.; Liu, M.; Zhao, X.; Yang, Q.; et al. Flavivirus RNA-Dependent RNA Polymerase Interacts with Genome UTRs and Viral Proteins to Facilitate Flavivirus RNA Replication. Viruses 2019, 11, 929. [Google Scholar] [CrossRef] [PubMed]
- Corver, J.; Lenches, E.; Smith, K.; Robison, R.A.; Sando, T.; Strauss, E.G.; Strauss, J.H. Fine mapping of a-acting sequence element in yellow fever virus RNA that is required for RNA replication and cyclization. J. Virol. 2003, 77, 2265–2270. [Google Scholar] [CrossRef] [PubMed]
- Molenkamp, R.; Kooi, E.A.; Lucassen, M.A.; Greve, S.; Thijssen, J.C.P.; Spaan, W.J.M.; Bredenbeek, P.J. Yellow fever virus replicons as an expression system for hepatitis C virus structural proteins. J. Virol. 2003, 77, 1644–1648. [Google Scholar] [CrossRef]
- Ishikawa, T.; Narita, K.; Matsuyama, K.; Masuda, M. Dissemination of the Flavivirus Subgenomic Replicon Genome and Viral Proteins by Extracellular Vesicles. Viruses 2024, 16, 524. [Google Scholar] [CrossRef]
- Huang, S.H.; Shi, P.D.; Fan, X.X.; Yang, Y.; Qin, C.F.; Zhao, H.; Shi, L.; Ci, Y.L. The glycosylation deficiency of flavivirus NS1 attenuates virus replication through interfering with the formation of viral replication compartments. J. Biomed. Sci. 2024, 31, 60. [Google Scholar] [CrossRef]
- Huang, Y.T.; Liao, J.T.; Yen, L.C.; Chang, Y.K.; Lin, Y.L.; Liao, C.L. Japanese encephalitis virus replicon-based vaccine expressing enterovirus-71 epitope confers dual protection from lethal challenges. J. Biomed. Sci. 2015, 22, 74. [Google Scholar] [CrossRef]
- Suzuki, R.; Ishikawa, T.; Konishi, E.; Matsuda, M.; Watashi, K.; Aizaki, H.; Takasaki, T.; Wakita, T. Production of single-round infectious chimeric flaviviruses with DNA-based Japanese encephalitis virus replicon. J. Gen. Virol. 2014, 95, 60–65. [Google Scholar] [CrossRef]
- Xie, X.; Wang, Q.Y.; Xu, H.Y.; Qing, M.; Kramer, L.; Yuan, Z.; Shi, P.Y. Inhibition of dengue virus by targeting viral NS4B protein. J. Virol. 2011, 85, 11183–11195. [Google Scholar] [CrossRef]
- Schnellrath, L.C.; Damaso, C.R. Potent antiviral activity of brequinar against the emerging Cantagalo virus in cell culture. Int. J. Antimicrob. Agents 2011, 38, 435–441. [Google Scholar] [CrossRef]
- Sourimant, J.; Lieber, C.M.; Aggarwal, M.; Cox, R.M.; Wolf, J.D.; Yoon, J.J.; Toots, M.; Ye, C.; Sticher, Z.; Kolykhalov, A.A.; et al. 4′-Fluorouridine is an oral antiviral that blocks respiratory syncytial virus and SARS-CoV-2 replication. Science 2022, 375, 161–167. [Google Scholar] [CrossRef]
- Kummerer, B.M. Establishment and Application of Flavivirus Replicons. Adv. Exp. Med. Biol. 2018, 1062, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Scaturro, P.; Cortese, M.; Chatel-Chaix, L.; Fischl, W.; Bartenschlager, R. Dengue Virus Non-structural Protein 1 Modulates Infectious Particle Production via Interaction with the Structural Proteins. PLoS Pathog. 2015, 11, e1005277. [Google Scholar] [CrossRef]
- Edeling, M.A.; Diamond, M.S.; Fremont, D.H. Structural basis of Flavivirus NS1 assembly and antibody recognition. Proc. Natl. Acad. Sci. USA 2014, 111, 4285–4290. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Zou, J.; Zhang, X.; Zhou, Y.; Routh, A.L.; Kang, C.; Popov, V.L.; Chen, X.; Wang, Q.Y.; Dong, H.; et al. Dengue NS2A Protein Orchestrates Virus Assembly. Cell Host Microbe 2019, 26, 606–622.e8. [Google Scholar] [CrossRef]
- Xie, X.; Zou, J.; Shan, C.; Yang, Y.; Kum, D.B.; Dallmeier, K.; Neyts, J.; Shi, P.Y. Zika Virus Replicons for Drug Discovery. eBioMedicine 2016, 12, 156–160. [Google Scholar] [CrossRef]
- Li, W.; Ma, L.; Guo, L.P.; Wang, X.L.; Zhang, J.W.; Bu, Z.G.; Hua, R.H. West Nile virus infectious replicon particles generated using a packaging-restricted cell line is a safe reporter system. Sci. Rep. 2017, 7, 3286. [Google Scholar] [CrossRef]
- Lundstrom, K. Application of DNA Replicons in Gene Therapy and Vaccine Development. Pharmaceutics 2023, 15, 947. [Google Scholar] [CrossRef]
- Lim, J.W.; Vu, T.T.H.; Le, V.P.; Yeom, M.; Song, D.; Jeong, D.G.; Park, S.K. Advanced Strategies for Developing Vaccines and Diagnostic Tools for African Swine Fever. Viruses 2023, 15, 2169. [Google Scholar] [CrossRef]
- Yun, S.I.; Song, B.H.; Koo, Y.; Jeon, I.; Byun, S.J.; Park, J.H.; Joo, Y.S.; Kim, S.Y.; Lee, Y.M. Japanese encephalitis virus-based replicon RNAs/particles as an expression system for HIV-1 Pr55 that is capable of producing virus-like particles. Virus Res. 2009, 144, 298–305. [Google Scholar] [CrossRef]
- Kiemel, D.; Kroell, A.H.; Denolly, S.; Haselmann, U.; Bonfanti, J.F.; Andres, J.I.; Ghosh, B.; Geluykens, P.; Kaptein, S.J.F.; Wilken, L.; et al. Pan-serotype dengue virus inhibitor JNJ-A07 targets NS4A-2K-NS4B interaction with NS2B/NS3 and blocks replication organelle formation. Nat. Commun. 2024, 15, 6080. [Google Scholar] [CrossRef]
- Kaptein, S.J.F.; Goethals, O.; Kiemel, D.; Marchand, A.; Kesteleyn, B.; Bonfanti, J.F.; Bardiot, D.; Stoops, B.; Jonckers, T.H.M.; Dallmeier, K.; et al. A pan-serotype dengue virus inhibitor targeting the NS3-NS4B interaction. Nature 2021, 598, 504–509. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Jing, J.F.; Tong, X.K.; He, P.L.; Li, Y.C.; Hu, Y.H.; Tang, W.; Zuo, J.P. Discovering novel anti-HCV compounds with inhibitory activities toward HCV NS3/4A protease. Acta Pharmacol. Sin. 2014, 35, 1074–1081. [Google Scholar] [CrossRef] [PubMed]
- Niyomrattanakit, P.; Chen, Y.L.; Dong, H.; Yin, Z.; Qing, M.; Glickman, J.F.; Lin, K.; Mueller, D.; Voshol, H.; Lim, J.Y.; et al. Inhibition of dengue virus polymerase by blocking of the RNA tunnel. J. Virol. 2010, 84, 5678–5686. [Google Scholar] [CrossRef]
- Yao, G.Q.; Yu, J.C.; Lin, C.; Zhu, Y.J.; Duan, A.N.; Li, M.F.; Yuan, J.; Zhang, J.C. Design, synthesis, and biological evaluation of novel 2’-methyl-2’-fluoro-6-methyl-7-alkynyl-7-deazapurine nucleoside analogs as anti-Zika virus agents. Eur. J. Med. Chem. 2022, 234, 114275. [Google Scholar] [CrossRef]
- Malakar, S.; Sreelatha, L.; Dechtawewat, T.; Noisakran, S.; Yenchitsomanus, P.T.; Chu, J.J.H.; Limjindaporn, T. Drug repurposing of quinine as antiviral against dengue virus infection. Virus Res. 2018, 255, 171–178. [Google Scholar] [CrossRef]
- Li, S.H.; Li, X.F.; Zhao, H.; Deng, Y.Q.; Yu, X.D.; Zhu, S.Y.; Jiang, T.; Ye, Q.; Qin, E.D.; Qin, C.F. Development and characterization of the replicon system of Japanese encephalitis live vaccine virus SA14-14-2. Virol. J. 2013, 10, 64. [Google Scholar] [CrossRef]
- Barnard, T.R.; Abram, Q.H.; Lin, Q.F.; Wang, A.B.; Sagan, S.M. Molecular Determinants of Flavivirus Virion Assembly. Trends Biochem. Sci. 2021, 46, 378–390. [Google Scholar] [CrossRef]
- Ci, Y.L.; Liu, Z.Y.; Zhang, N.N.; Niu, Y.Q.; Yang, Y.; Xu, C.M.; Yang, W.; Qin, C.F.; Shi, L. Zika NS1-induced ER remodeling is essential for viral replication. J. Cell Biol. 2020, 219, e201903062. [Google Scholar] [CrossRef]
- Brown, W.C.; Akey, D.L.; Konwerski, J.R.; Tarrasch, J.T.; Skiniotis, G.; Kuhn, R.J.; Smith, J.L. Extended surface for membrane association in Zika virus NS1 structure. Nat. Struct. Mol. Biol. 2016, 23, 865–867. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CC50 | EC50 | SI | |
|---|---|---|---|
| JNJ-A07 | 29.86 µM | 0.013 µM | 2296.92 |
| HZ-1157 | 21.22 µM | 1.06 µM | 20.02 |
| NITD-2 | 49.17 µM | 0.141 µM | 348.72 |
| NITD008 | 37.76 µM | 0.113 µM | 334.16 |
| Quinine | 45.76 µM | 0.162 µM | 282.47 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.; Zheng, J.; Lin, Y.; Zhang, Y.; Zhao, Q.; Zhang, H.; Zhang, J.; Li, Z.; Liu, K.; Li, B.; et al. Development of a Noninfectious Japanese Encephalitis Virus Replicon for Antiviral Drug Screening and Gene Function Studies. Viruses 2025, 17, 759. https://doi.org/10.3390/v17060759
Yang Y, Zheng J, Lin Y, Zhang Y, Zhao Q, Zhang H, Zhang J, Li Z, Liu K, Li B, et al. Development of a Noninfectious Japanese Encephalitis Virus Replicon for Antiviral Drug Screening and Gene Function Studies. Viruses. 2025; 17(6):759. https://doi.org/10.3390/v17060759
Chicago/Turabian StyleYang, Yang, Jiayang Zheng, Yafang Lin, Yan Zhang, Qianming Zhao, Hailong Zhang, Junjie Zhang, Zongjie Li, Ke Liu, Beibei Li, and et al. 2025. "Development of a Noninfectious Japanese Encephalitis Virus Replicon for Antiviral Drug Screening and Gene Function Studies" Viruses 17, no. 6: 759. https://doi.org/10.3390/v17060759
APA StyleYang, Y., Zheng, J., Lin, Y., Zhang, Y., Zhao, Q., Zhang, H., Zhang, J., Li, Z., Liu, K., Li, B., Shao, D., Qiu, Y., Ma, Z., & Wei, J. (2025). Development of a Noninfectious Japanese Encephalitis Virus Replicon for Antiviral Drug Screening and Gene Function Studies. Viruses, 17(6), 759. https://doi.org/10.3390/v17060759