
Detection of a New Recombinant Rabbit Hemorrhagic Disease Virus 2 in China and Development of Virus-like Particle-Based Vaccine

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Genotype Detection
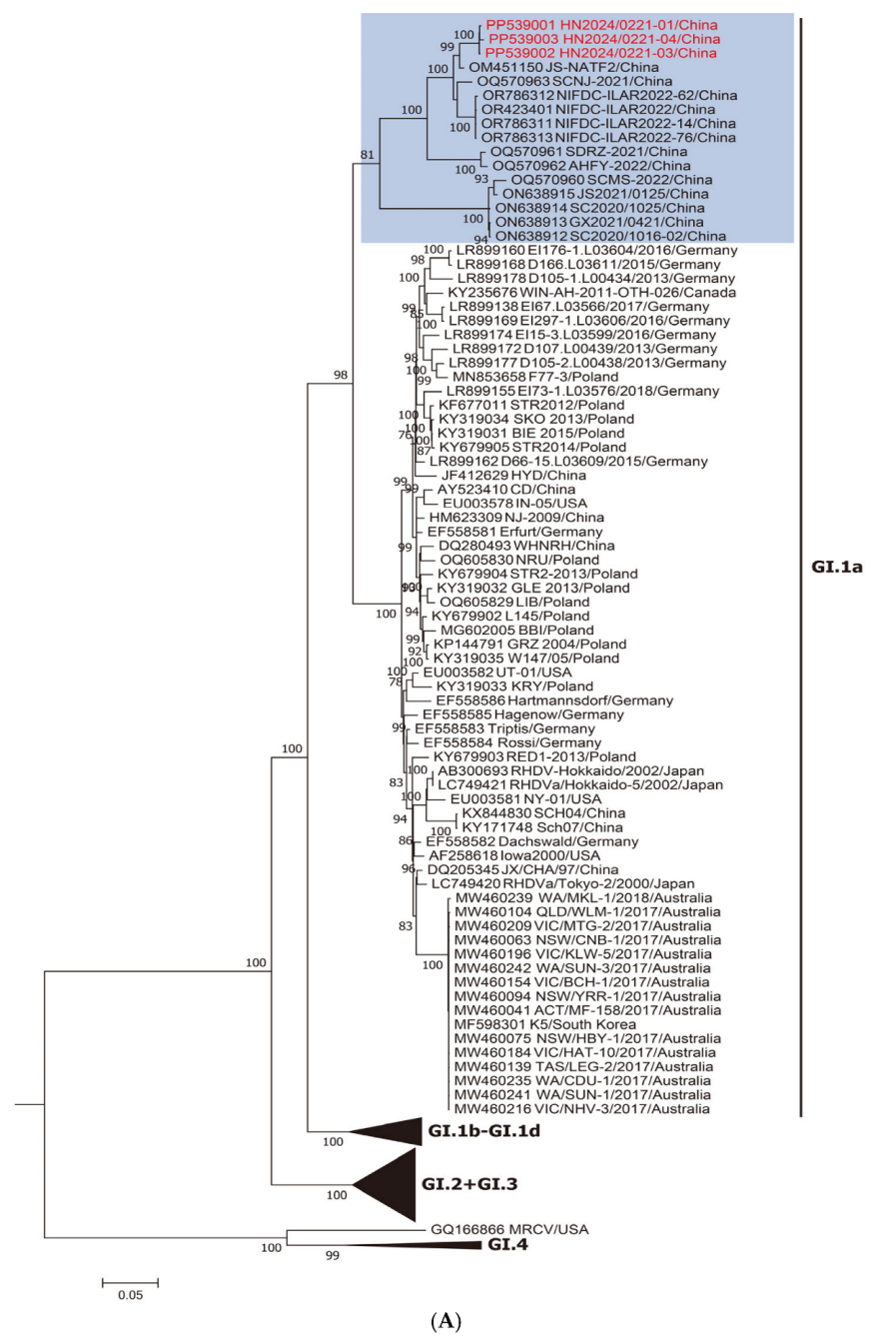
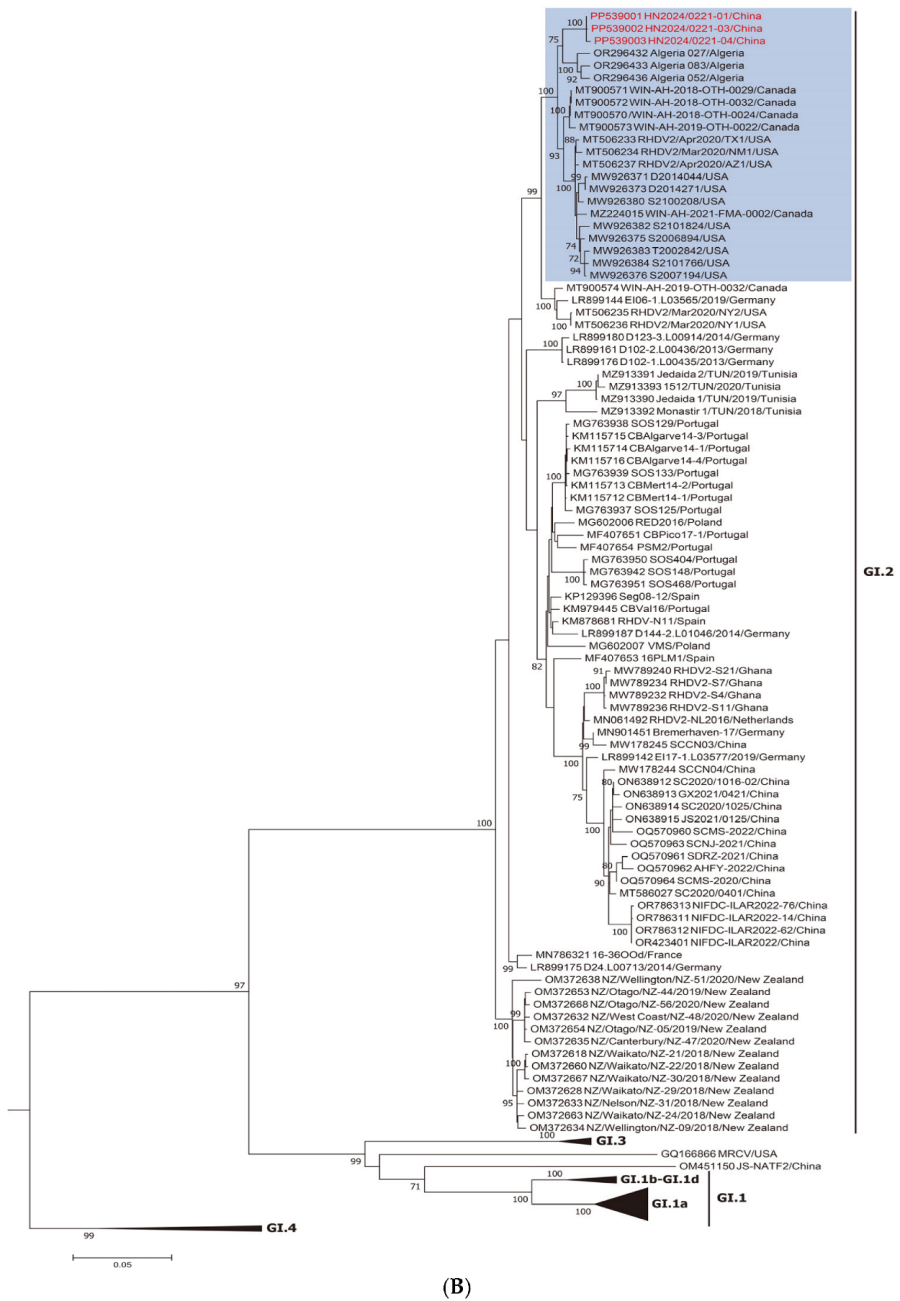
2.2. Phylogenetic Analysis
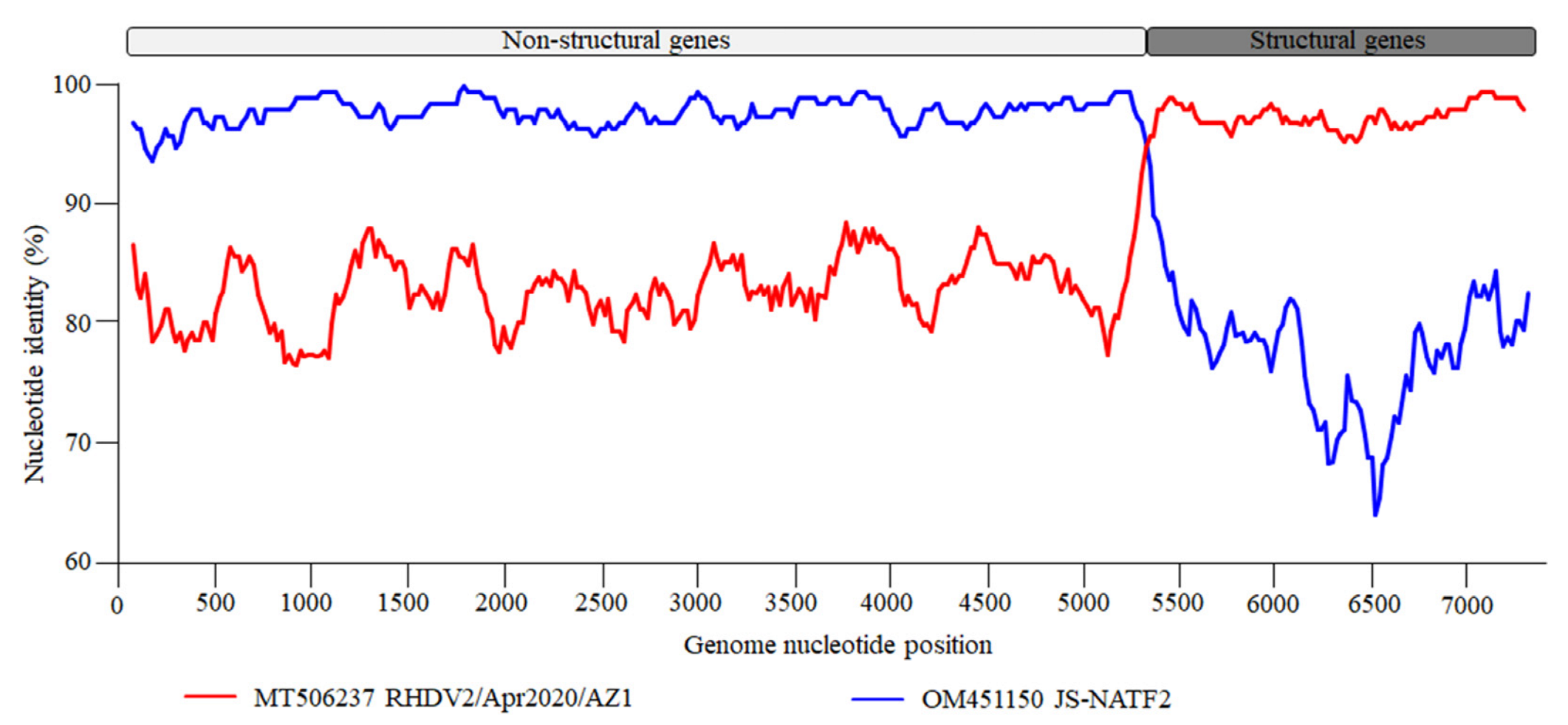
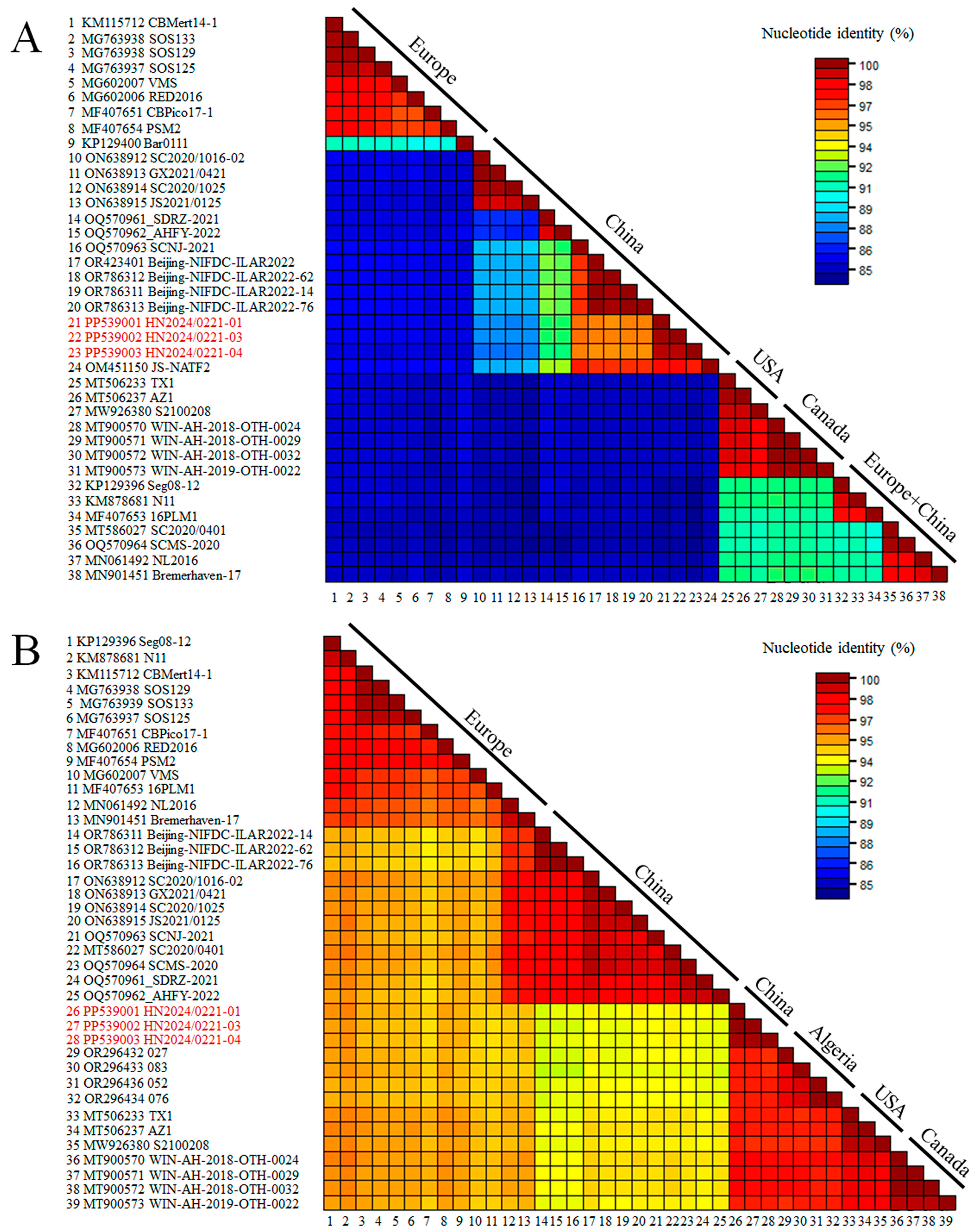
2.3. Recombination and Sequences Analysis
2.4. Challenge Study
3. Results
3.1. Genotype Detection and Phylogenetic Analysis
3.2. Recombination and Sequences Analysis
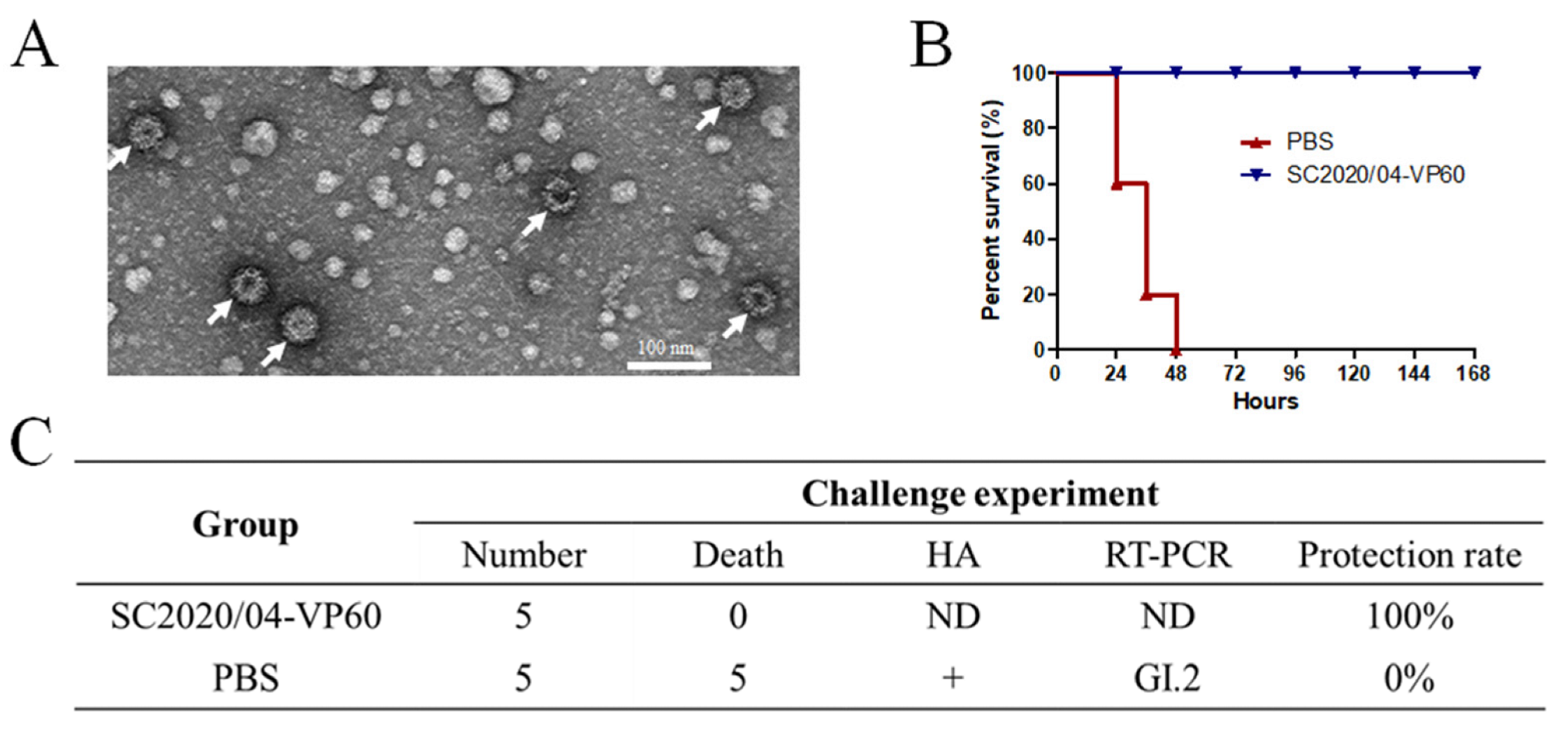
3.3. Generation of RHDV2 VLPs and Challenge Study
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| RHD | rabbit hemorrhagic disease |
| RHDV | rabbit hemorrhagic disease virus |
| VLPs | virus-like particles |
| ML | maximum likelihood |
| TEM | transmission electron microscope |
| HAU | hemagglutination units |
| EBHSV | European brown hare syndrome virus |
References
- Le Pendu, J.; Abrantes, J.; Bertagnoli, S.; Guitton, J.S.; Le Gall-Recule, G.; Lopes, A.M.; Marchandeau, S.; Alda, F.; Almeida, T.; Celio, A.P.; et al. Proposal for a unified classification system and nomenclature of lagoviruses. J. Gen. Virol. 2017, 98, 1658–1666. [Google Scholar] [CrossRef]
- Liu, S.; Xue, H.; Pu, B.; Qian, N. A new viral disease in rabbits. Anim. Husb. Vet. Med. 1984, 16, 253–255. [Google Scholar]
- Le Gall-Recule, G.; Lavazza, A.; Marchandeau, S.; Bertagnoli, S.; Zwingelstein, F.; Cavadini, P.; Martinelli, N.; Lombardi, G.; Guerin, J.L.; Lemaitre, E.; et al. Emergence of a new lagovirus related to Rabbit Haemorrhagic Disease Virus. Vet. Res. 2013, 44, 81. [Google Scholar] [CrossRef] [PubMed]
- Mahar, J.E.; Hall, R.N.; Peacock, D.; Kovaliski, J.; Piper, M.; Mourant, R.; Huang, N.; Campbell, S.; Gu, X.; Read, A.; et al. Rabbit Hemorrhagic Disease Virus 2 (RHDV2; GI.2) is replacing endemic strains of RHDV in the Australian landscape within 18 months of its arrival. J. Virol. 2018, 92, e01374-17. [Google Scholar] [CrossRef]
- Strive, T.; Piper, M.; Huang, N.; Mourant, R.; Kovaliski, J.; Capucci, L.; Cox, T.E.; Smith, I. Retrospective serological analysis reveals presence of the emerging lagovirus RHDV2 in Australia in wild rabbits at least five months prior to its first detection. Transbound. Emerg. Dis. 2020, 67, 822–833. [Google Scholar] [CrossRef]
- Lopes, A.M.; Rouco, C.; Esteves, P.J.; Abrantes, J. GI.1b/GI.1b/GI.2 recombinant rabbit hemorrhagic disease virus 2 (Lagovirus europaeus/GI.2) in Morocco, Africa. Arch. Virol. 2019, 164, 279–283. [Google Scholar] [CrossRef]
- Ben Chehida, F.; Lopes, A.M.; Côrte-Real, J.V.; Sghaier, S.; Aouini, R.; Messadi, L.; Abrantes, J. Multiple Introductions of Rabbit Hemorrhagic Disease Virus Lagovirus europaeus/GI.2 in Africa. Biology 2021, 10, 883. [Google Scholar] [CrossRef]
- Ambagala, A.; Schwantje, H.; Laurendeau, S.; Snyman, H.; Joseph, T.; Pickering, B.; Hooper-McGrevy, K.; Babiuk, S.; Moffat, E.; Lamboo, L.; et al. Incursions of rabbit haemorrhagic disease virus 2 in Canada-Clinical, molecular and epidemiological investigation. Transbound. Emerg. Dis. 2021, 68, 1711–1720. [Google Scholar] [CrossRef]
- Asin, J.; Rejmanek, D.; Clifford, D.L.; Mikolon, A.B.; Henderson, E.E.; Nyaoke, A.C.; Macías-Rioseco, M.; Streitenberger, N.; Beingesser, J.; Woods, L.W.; et al. Early circulation of rabbit haemorrhagic disease virus type 2 in domestic and wild lagomorphs in southern California, USA (2020–2021). Transbound. Emerg. Dis. 2022, 69, e394–e405. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Wei, H.; Fan, Z.; Song, Y.; Chen, M.; Qiu, R.; Zhu, W.; Xu, W.; Xue, J.; Wang, F. Emergence of rabbit haemorrhagic disease virus 2 in China in 2020. Vet. Med. Sci. 2021, 7, 236–239. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Fan, Z.; Hu, B.; Song, Y.; Wei, H.; Qiu, R.; Zhu, W.; Xu, W.; Wang, F. Pathogenicity of the newly emerged Lagovirus europaeus GI.2 strain in China in experimentally infected rabbits. Vet. Microbiol. 2022, 265, 109311. [Google Scholar] [CrossRef]
- Qi, R.; Meng, C.; Zhu, J.; Li, H.; Miao, Q.; Tang, J.; Tang, A.; Guo, H.; Liu, C.; Li, C.; et al. The outbreak of rabbit hemorrhagic virus type 2 in the interior of China may be related to imported semen. Virol. Sin. 2022, 37, 623–626. [Google Scholar] [CrossRef]
- Lopes, A.M.; Dalton, K.P.; Magalhaes, M.J.; Parra, F.; Esteves, P.J.; Holmes, E.C.; Abrantes, J. Full genomic analysis of new variant rabbit hemorrhagic disease virus revealed multiple recombination events. J. Gen. Virol. 2015, 96, 1309–1319. [Google Scholar] [CrossRef] [PubMed]
- Forrester, N.L.; Moss, S.R.; Turner, S.L.; Schirrmeier, H.; Gould, E.A. Recombination in rabbit haemorrhagic disease virus: Possible impact on evolution and epidemiology. Virology 2008, 376, 390–396. [Google Scholar] [CrossRef]
- Mahar, J.E.; Jenckel, M.; Huang, N.; Smertina, E.; Holmes, E.C.; Strive, T.; Hall, R.N. Frequent intergenotypic recombination between the non-structural and structural genes is a major driver of epidemiological fitness in caliciviruses. Virus Evol. 2021, 7, veab080. [Google Scholar] [CrossRef] [PubMed]
- Abrantes, J.; Droillard, C.; Lopes, A.M.; Lemaitre, E.; Lucas, P.; Blanchard, Y.; Marchandeau, S.; Esteves, P.J.; Le Gall-Recule, G. Recombination at the emergence of the pathogenic rabbit haemorrhagic disease virus Lagovirus europaeus/GI.2. Sci. Rep. 2020, 10, 14502. [Google Scholar] [CrossRef]
- Silverio, D.; Lopes, A.M.; Melo-Ferreira, J.; Magalhaes, M.J.; Monterroso, P.; Serronha, A.; Maio, E.; Alves, P.C.; Esteves, P.J.; Abrantes, J. Insights into the evolution of the new variant rabbit haemorrhagic disease virus (GI.2) and the identification of novel recombinant strains. Transbound. Emerg. Dis. 2018, 65, 983–992. [Google Scholar] [CrossRef]
- Hall, R.N.; Mahar, J.E.; Read, A.J.; Mourant, R.; Piper, M.; Huang, N.; Strive, T. A strain-specific multiplex RT-PCR for Australian rabbit haemorrhagic disease viruses uncovers a new recombinant virus variant in rabbits and hares. Transbound. Emerg. Dis. 2018, 65, e444–e456. [Google Scholar] [CrossRef]
- Cavadini, P.; Trogu, T.; Velarde, R.; Lavazza, A.; Capucci, L. Recombination between non-structural and structural genes as a mechanism of selection in lagoviruses: The evolutionary dead-end of an RHDV2 isolated from European hare. Virus Res. 2024, 339, 199257. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Fan, Z.; Qiu, R.; Chen, M.; Wei, H.; Song, Y.; Liu, W.; Xu, W.; Wang, F. Novel recombinant rabbit hemorrhagic disease virus 2 (RHDV2) is circulating in China within 12 months after original RHDV2 arrival. Transbound. Emerg. Dis. 2023, 2023, 4787785. [Google Scholar] [CrossRef]
- Li, Y.; Du, D.; Zhou, L.; Su, L.; You, C.; Zhang, H.; Yu, J.; Xiao, L.; Huang, J. First report of GI.1aP-GI.2 recombinants of rabbit hemorrhagic disease virus in domestic rabbits in China. Front. Microbiol. 2023, 14, 1188380. [Google Scholar] [CrossRef]
- Dalton, K.P.; Abrantes, J.; Lopes, A.M.; Nicieza, I.; Álvarez, Á.L.; Esteves, P.J.; Parra, F. Complete genome sequence of two rabbit hemorrhagic disease virus variant b isolates detected on the Iberian Peninsula. Arch. Virol. 2015, 160, 877–881. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Dong, W.; Fan, Z.; Wei, H.; Song, Y.; Chen, M.; Qiu, R.; Ge, L.; Li, Y.; Xu, W.; et al. Genetic variability and phylogenetic analysis of rabbit hemorrhagic disease virus (RHDV) in China from 2020 to 2022. In Proceedings of the 13th World Rabbit Congress, Tarragona, Spain, 2–4 October 2024; Spain-Pathology and Hygiene Session: Tarragona, Spain; pp. 629–632. [Google Scholar]
- Nagesha, H.S.; Wang, L.F.; Hyatt, A.D.; Morrissy, C.J.; Lenghaus, C.; Westbury, H.A. Self-assembly, antigenicity, and immunogenicity of the rabbit haemorrhagic disease virus (Czechoslovakian strain V-351) capsid protein expressed in baculovirus. Arch. Virol. 1995, 140, 1095–1108. [Google Scholar] [CrossRef]
- Bárcena, J.; Guerra, B.; Angulo, I.; González, J.; Valcárcel, F.; Mata, C.P.; Castón, J.R.; Blanco, E.; Alejo, A. Comparative analysis of rabbit hemorrhagic disease virus (RHDV) and new RHDV2 virus antigenicity, using specific virus-like particles. Vet. Res. 2015, 46, 106. [Google Scholar] [CrossRef]
- Fitzner, A.; Kesy, A.; Bulenger, K.; Niedbalski, W. Evidence of independent introductions of RHDV2 strains in Poland based on the genome analysis of viral isolates from 2016–2018. Acta. Biochim. Pol. 2021, 68, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Dalton, K.P.; Alvarado, C.; Reytor, E.; Del, C.N.M.; Podadera, A.; Martinez-Alonso, D.; Alonso, J.; Nicieza, I.; Gomez-Sebastian, S.; Dalton, R.M.; et al. Chimeric VLPs Bearing VP60 from Two Serotypes of Rabbit Haemorrhagic Disease Virus Are Protective against Both Viruses. Vaccines 2021, 9, 1005. [Google Scholar] [CrossRef]
- Muller, C.; Ulrich, R.; Schinkothe, J.; Muller, M.; Kollner, B. Characterization of protective humoral and cellular immune responses against RHDV2 induced by a new vaccine based on recombinant baculovirus. Vaccine 2019, 37, 4195–4203. [Google Scholar] [CrossRef]
- Rahali, N.; Sghaier, S.; Kbaier, H.; Zanati, A.; Bahloul, C. Genetic characterization and phylogenetic analysis of rabbit hemorrhagic disease virus isolated in Tunisia from 2015 to 2018. Arch. Virol. 2019, 164, 2327–2332. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Martin, D.P.; Varsani, A.; Roumagnac, P.; Botha, G.; Maslamoney, S.; Schwab, T.; Kelz, Z.; Kumar, V.; Murrell, B. RDP5: A computer program for analyzing recombination in, and removing signals of recombination from, nucleotide sequence datasets. Virus Evol. 2021, 7, veaa087. [Google Scholar] [CrossRef]
- Lole, K.S.; Bollinger, R.C.; Paranjape, R.S.; Gadkari, D.; Kulkarni, S.S.; Novak, N.G.; Ingersoll, R.; Sheppard, H.W.; Ray, S.C. Full-length human immunodeficiency virus type 1 genomes from subtype C-infected seroconverters in India, with evidence of intersubtype recombination. J. Virol. 1999, 73, 152–160. [Google Scholar] [CrossRef] [PubMed]
- Muhire, B.M.; Varsani, A.; Martin, D.P. SDT: A virus classification tool based on pairwise sequence alignment and identity calculation. PLoS ONE 2014, 9, e108277. [Google Scholar] [CrossRef]
- Chen, M.; Song, Y.; Fan, Z.; Jiang, P.; Hu, B.; Xue, J.; Wei, H.; Wang, F. Immunogenicity of different recombinant rabbit hemorrhagic disease virus-like particles carrying CD8+ T cell epitope from chicken ovalbumin (OVA). Virus Res. 2014, 183, 15–22. [Google Scholar] [CrossRef]
- San-Miguel, B.; Crespo, I.; Vallejo, D.; Alvarez, M.; Prieto, J.; Gonzalez-Gallego, J.; Tunon, M.J. Melatonin modulates the autophagic response in acute liver failure induced by the rabbit hemorrhagic disease virus. J. Pineal. Res. 2014, 56, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Sahraoui, L.; Lahouassa, H.; Maziz-Bettahar, S.; Lopes, A.M.; Almeida, T.; Ainbaziz, H.; Abrantes, J. First detection and molecular characterization of rabbit hemorrhagic disease virus (RHDV) in Algeria. Front. Vet. Sci. 2023, 10, 1235123. [Google Scholar] [CrossRef]
- Wu, L. Rabbit meat trade of major countries: Regional pattern and driving forces. World Rabbit Sci. 2022, 30, 69–82. [Google Scholar] [CrossRef]
- Esteves, P.J.; Abrantes, J.; Bertagnoli, S.; Cavadini, P.; Gavier-Widen, D.; Guitton, J.S.; Lavazza, A.; Lemaitre, E.; Letty, J.; Lopes, A.M.; et al. Emergence of Pathogenicity in Lagoviruses: Evolution from Pre-existing Nonpathogenic Strains or through a Species Jump? PLoS Pathog. 2015, 11, e1005087. [Google Scholar] [CrossRef]
- He, W.T.; Hou, X.; Zhao, J.; Sun, J.; He, H.; Si, W.; Wang, J.; Jiang, Z.; Yan, Z.; Xing, G.; et al. Virome characterization of game animals in China reveals a spectrum of emerging pathogens. Cell 2022, 185, 1117–1129. [Google Scholar] [CrossRef]
- Bosco-Lauth, A.M.; Schueler, A.; Midthun, E.; Tyra, H.; Held, A.; Hood, C.; Quilici, M.; Erickson, S.; Glover, S.; Gustafson, B.; et al. Vaccination against Rabbit Hemorrhagic Disease Virus 2 (RHDV2) Using a Baculovirus Recombinant Vaccine Provides Durable Immunity in Rabbits. Viruses 2024, 16, 538. [Google Scholar] [CrossRef]
- Akache, B.; Read, A.J.; Dudani, R.; Harrison, B.A.; Williams, D.; Deschatelets, L.; Jia, Y.; Chandan, V.; Stark, F.C.; Agbayani, G.; et al. Sulfated Lactosyl Archaeol Archaeosome-Adjuvanted Vaccine Formulations Targeting Rabbit Hemorrhagic Disease Virus Are Immunogenic and Efficacious. Vaccines 2023, 11, 1043. [Google Scholar] [CrossRef]
- Perozo-Mur, E.; Fontseca, M.; Nadal, G. Efficacy evaluation of YURVAC® RHD against different strains of rabbit haemorrhagic disease. In Proceedings of the 13th World Rabbit Congress, Tarragona, Spain, 2–4 October 2024; Spain-Pathology and Hygiene Session: Tarragona, Spain; pp. 637–639. [Google Scholar]
- European Medical Agency. Fatrovax RHD (Rabbit Haemorrhagic Disease Vaccine [Inactivated, Recombinant]). 2021. Available online: https://www.ema.europa.eu/en/medicines/veterinary/EPAR/fatrovax-rhd (accessed on 24 March 2025).
- Mukhin, A.N.; Alekseev, K.P.; Yuzhakov, A.G.; Selezneva, E.V.; Moskvinsmall, A.C.A.; Verkhovsky, O.A.; Aliper, T.I. Antigenic and immunogenic activity of virus-like particles based on rabbit hemorrhagic disease virus (Caliciviridae: Lagovirus) genotypes GI1 and GI2 recombinant major capsid proteins. Vopr. Virusol. 2023, 68, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Banadyga, L.; Zhao, Y.; Zhao, Z.; Schiffman, Z.; Huang, P.; Li, E.; Wang, C.; Gao, Y.; Feng, N.; et al. Peste des Petits Ruminants Virus-Like Particles Induce a Potent Humoral and Cellular Immune Response in Goats. Viruses 2019, 11, 918. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Most Likely Donor Strain | Methods and Average p-Values | |||||||
|---|---|---|---|---|---|---|---|---|---|
| NSP 1 | SP 2 | RDP | GENECONV | BootScan | MaxChi | Chimaera | SiScan | 3Seq | |
| HN2024/0221-01 (PP539001) | JS-NATF2 (OM451150) | AZ1 (MT506237) | 2.145 × 10−46 | 3.002 × 10−109 | 1.671 × 10−112 | 2.545 × 10−37 | 2.381 × 10−39 | 4.786 × 10−43 | 1.912 × 10−174 |
| HN2024/0221-03 (PP539002) | |||||||||
| HN2024/0221-04 (PP539003) | |||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, B.; Dong, W.; Song, Y.; Fan, Z.; Cavadini, P.; Wang, F. Detection of a New Recombinant Rabbit Hemorrhagic Disease Virus 2 in China and Development of Virus-like Particle-Based Vaccine. Viruses 2025, 17, 710. https://doi.org/10.3390/v17050710
Hu B, Dong W, Song Y, Fan Z, Cavadini P, Wang F. Detection of a New Recombinant Rabbit Hemorrhagic Disease Virus 2 in China and Development of Virus-like Particle-Based Vaccine. Viruses. 2025; 17(5):710. https://doi.org/10.3390/v17050710
Chicago/Turabian StyleHu, Bo, Wenyu Dong, Yanhua Song, Zhiyu Fan, Patrizia Cavadini, and Fang Wang. 2025. "Detection of a New Recombinant Rabbit Hemorrhagic Disease Virus 2 in China and Development of Virus-like Particle-Based Vaccine" Viruses 17, no. 5: 710. https://doi.org/10.3390/v17050710
APA StyleHu, B., Dong, W., Song, Y., Fan, Z., Cavadini, P., & Wang, F. (2025). Detection of a New Recombinant Rabbit Hemorrhagic Disease Virus 2 in China and Development of Virus-like Particle-Based Vaccine. Viruses, 17(5), 710. https://doi.org/10.3390/v17050710