

Tegument Protein pUL47 Is Important but Not Essential for Horizontal Transmission of Vaccinal Strain SB-1 of Gallid Alphaherpesvirus 3
 and
and
Abstract
1. Introduction
2. Methods
2.1. Ethics Statement
2.2. pUL47 Sequence Alignment
2.3. Cells
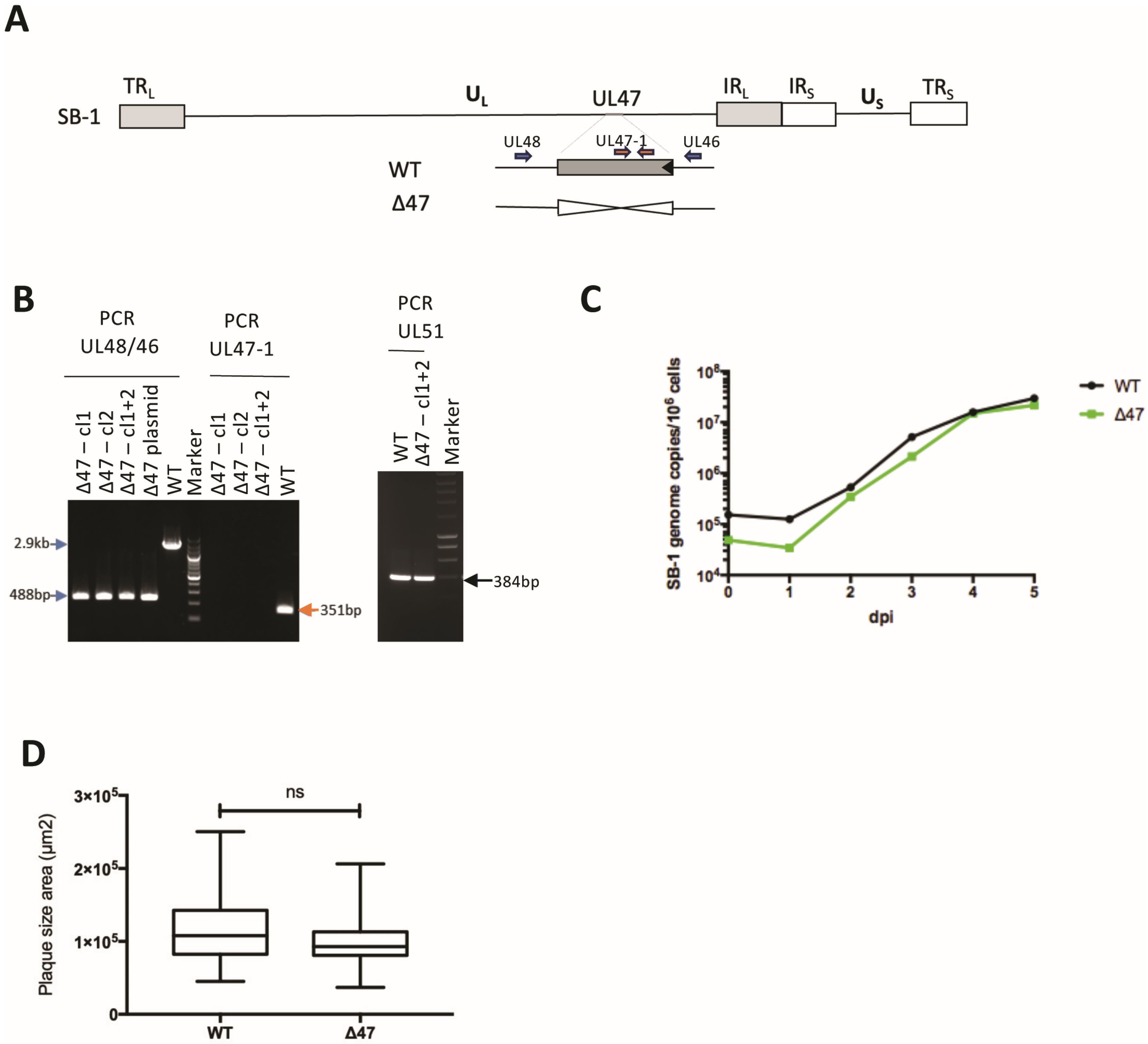
2.4. Generation of SB-1 Mutant and Virus Propagation
2.5. Multi-Step Growth Kinetics
2.6. Plaque Size Assays
2.7. Animal Experiment
2.8. DNA Extraction from PBMCs, Splenocytes, Feather Samples, and Dust
2.9. Virus Quantification Through Real-Time PCR in PBMCs, Splenocytes, Feather Samples, and Dust
2.10. Viral PCR Analyses
2.11. Statistical Analyses
3. Results
3.1. UL47 Is Dispensable for Vaccine SB-1 Replication In Vitro
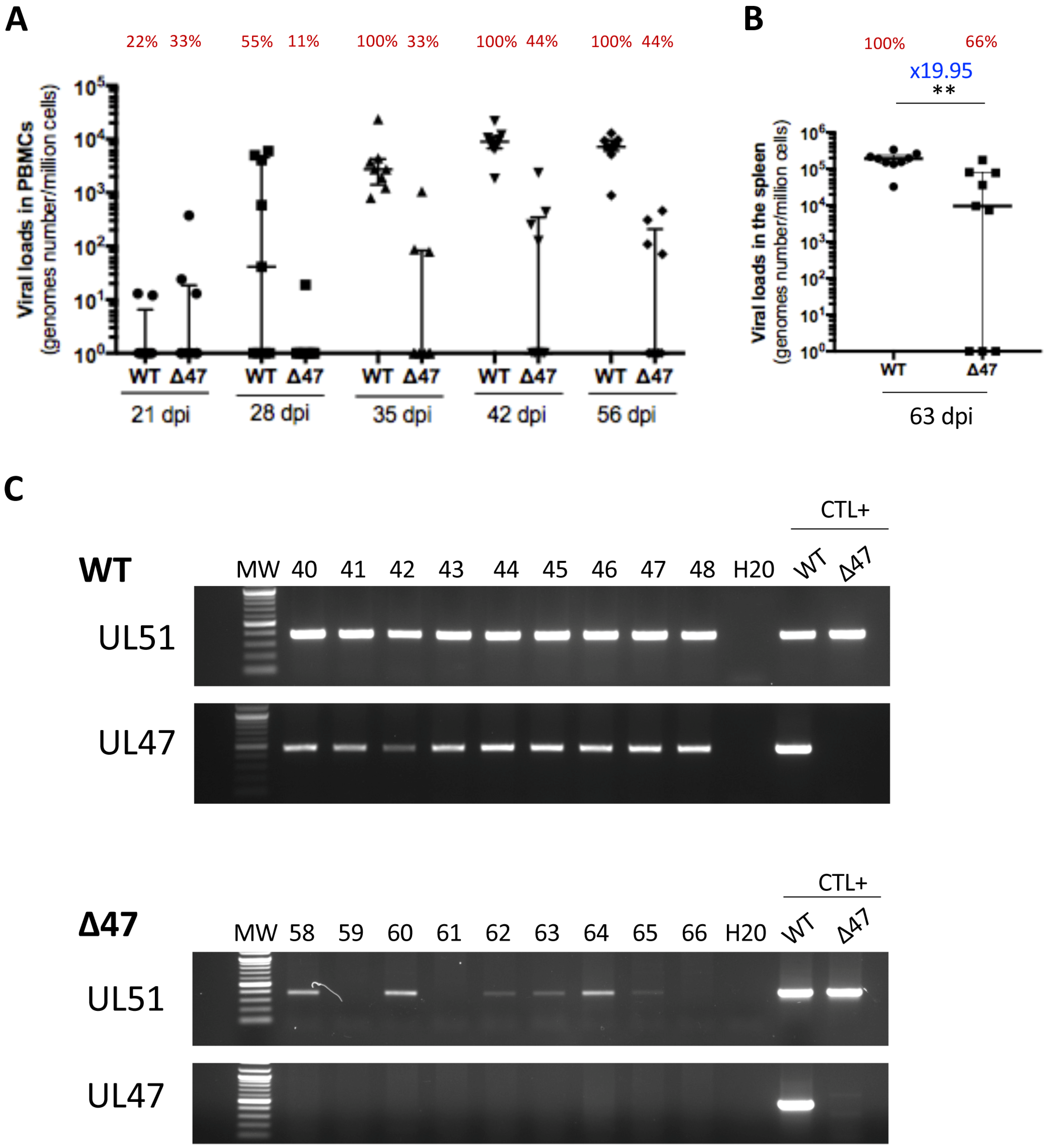
3.2. The Absence of UL47 Reduces SB-1 Viral Load in the PBMCs and the Spleen of Inoculated Chickens
3.3. The Absence of UL47 Delays Virus Transport to the Skin of Inoculated Chickens but Does Not Compromise Replication and Persistence in Feathers
3.4. Tegument Protein pUL47 Is Important but Not Essential for Horizontal Transmission Between Chickens
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Schat, K.A.; Calnek, B.W. Characterization of an apparently nononcogenic Marek’s disease virus. J. Natl. Cancer Inst. 1978, 60, 1075–1082. [Google Scholar] [CrossRef]
- Schat, K.A. History of the First-Generation Marek’s Disease Vaccines: The Science and Little-Known Facts. Avian Dis. 2016, 60, 715–724. [Google Scholar] [CrossRef]
- Nair, V.; Gimeno, I.M.; Dunn, J.R. Marek’s Disease. In Diseases of Poultry (14th edition); Swayne, D.E., Boulianne, M., Logue, C.M., McDougald, L.R., Nair, V., Suarez, D.L., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2018; pp. 550–586. [Google Scholar]
- Spatz, S.J.; Schat, K.A. Comparative genomic sequence analysis of the Marek’s disease vaccine strain SB-1. Virus Genes 2011, 42, 331–338. [Google Scholar] [CrossRef]
- Cho, B.R. Dual virus maturation of both pathogenic and apathogenic Marek’s disease herpesvirus (MDHV) in the feather follicles of dually infected chickens. Avian Dis. 1977, 21, 501–507. [Google Scholar] [CrossRef]
- Renz, K.G.; Islam, A.; Cheetham, B.F.; Walkden-Brown, S.W. Absolute quantification using real-time polymerase chain reaction of Marek’s disease virus serotype 2 in field dust samples, feather tips and spleens. J. Virol. Methods 2006, 135, 186–191. [Google Scholar] [CrossRef]
- Cortes, A.L.; Montiel, E.R.; Lemiere, S.; Gimeno, I.M. Comparison of blood and feather pulp samples for the diagnosis of Marek’s disease and for monitoring Marek’s disease vaccination by real time-PCR. Avian Dis. 2011, 55, 302–310. [Google Scholar] [CrossRef]
- McPherson, M.C.; Cheng, H.H.; Delany, M.E. Marek’s disease herpesvirus vaccines integrate into chicken host chromosomes yet lack a virus-host phenotype associated with oncogenic transformation. Vaccine 2016, 34, 5554–5561. [Google Scholar] [CrossRef]
- Krieter, A.; Xu, H.; Akbar, H.; Kim, T.; Jarosinski, K.W. The Conserved Herpesviridae Protein Kinase (CHPK) of Gallid alphaherpesvirus 3 (GaHV3) Is Required for Horizontal Spread and Natural Infection in Chickens. Viruses 2022, 14, 586. [Google Scholar] [CrossRef]
- Hein, R.; Koopman, R.; Garcia, M.; Armour, N.; Dunn, J.R.; Barbosa, T.; Martinez, A. Review of Poultry Recombinant Vector Vaccines. Avian Dis. 2021, 65, 438–452. [Google Scholar] [CrossRef]
- Darteil, R.; Bublot, M.; Laplace, E.; Bouquet, J.F.; Audonnet, J.C.; Riviere, M. Herpesvirus of turkey recombinant viruses expressing infectious bursal disease virus (IBDV) VP2 immunogen induce protection against an IBDV virulent challenge in chickens. Virology 1995, 211, 481–490. [Google Scholar] [CrossRef]
- Le Gros, F.X.; Dancer, A.; Giacomini, C.; Pizzoni, L.; Bublot, M.; Graziani, M.; Prandini, F. Field efficacy trial of a novel HVT-IBD vector vaccine for 1-day-old broilers. Vaccine 2009, 27, 592–596. [Google Scholar] [CrossRef]
- Rauw, F.; Palya, V.; Gardin, Y.; Tatar-Kis, T.; Dorsey, K.M.; Lambrecht, B.; van den Berg, T. Efficacy of rHVT-AI vector vaccine in broilers with passive immunity against challenge with two antigenically divergent Egyptian clade 2.2.1 HPAI H5N1 strains. Avian Dis. 2012, 56, 913–922. [Google Scholar] [CrossRef]
- Esaki, M.; Godoy, A.; Rosenberger, J.K.; Rosenberger, S.C.; Gardin, Y.; Yasuda, A.; Dorsey, K.M. Protection and antibody response caused by turkey herpesvirus vector Newcastle disease vaccine. Avian Dis. 2013, 57, 750–755. [Google Scholar] [CrossRef]
- Gergen, L.; Cook, S.; Ledesma, B.; Cress, W.; Higuchi, D.; Counts, D.; Cruz-Coy, J.; Crouch, C.; Davis, P.; Tarpey, I.; et al. A double recombinant herpes virus of turkeys for the protection of chickens against Newcastle, infectious laryngotracheitis and Marek’s diseases. Avian Pathol. 2019, 48, 45–56. [Google Scholar] [CrossRef]
- Tang, N.; Zhang, Y.; Sadigh, Y.; Moffat, K.; Shen, Z.; Nair, V.; Yao, Y. Generation of A Triple Insert Live Avian Herpesvirus Vectored Vaccine Using CRISPR/Cas9-Based Gene Editing. Vaccines 2020, 8, 97. [Google Scholar] [CrossRef]
- Criado, M.F.; Kassa, A.; Bertran, K.; Kwon, J.H.; Sa, E.S.M.; Killmaster, L.; Ross, T.M.; Mebatsion, T.; Swayne, D.E. Efficacy of multivalent recombinant herpesvirus of turkey vaccines against high pathogenicity avian influenza, infectious bursal disease, and Newcastle disease viruses. Vaccine 2023, 41, 2893–2904. [Google Scholar] [CrossRef]
- Sadigh, Y.; Powers, C.; Spiro, S.; Pedrera, M.; Broadbent, A.; Nair, V. Gallid herpesvirus 3 SB-1 strain as a recombinant viral vector for poultry vaccination. NPJ Vaccines 2018, 3, 21. [Google Scholar] [CrossRef]
- Calnek, B.W.; Schat, K.A.; Peckham, M.C.; Fabricant, J. Field trials with a bivalent vaccine (HVT and SB-1) against Marek’s disease. Avian Dis. 1983, 27, 844–849. [Google Scholar] [CrossRef]
- Witter, R.L. New serotype 2 and attenuated serotype 1 Marek’s disease vaccine viruses: Comparative efficacy. Avian Dis. 1987, 31, 752–765. [Google Scholar] [CrossRef]
- Islam, A.; Walkden-Brown, S.W. Quantitative profiling of the shedding rate of the three Marek’s disease virus (MDV) serotypes reveals that challenge with virulent MDV markedly increases shedding of vaccinal viruses. J. Gen. Virol. 2007, 88, 2121–2128. [Google Scholar] [CrossRef]
- Hirai, K.; Yamada, M.; Arao, Y.; Kato, S.; Nii, S. Replicating Marek’s disease virus (MDV) serotype 2 DNA with inserted MDV serotype 1 DNA sequences in a Marek’s disease lymphoblastoid cell line MSB1-41C. Arch. Virol. 1990, 114, 153–165. [Google Scholar] [CrossRef]
- Denesvre, C.; Dumarest, M.; Rémy, S.; Gourichon, D.; Eloit, M. Chicken skin virome analyzed by high-throughput sequencing shows a composition highly different from human skin. Virus Genes. 2015, 5, 209–216. [Google Scholar] [CrossRef]
- Chakravarti, S.; Mescolini, G.; Parker, D.; Prins, C.; Baigent, S.; Nair, V.; Yao, Y. Effects of Co-infections on Marek’s Disease in UK Poultry Farms and Development of Novel Rapid Diagnostic Strategies. In Proceedings of the 14th International Symposium on Marek’s Disease and Avian Herpesviruses, St Louis, MO, USA, 12–14 July 2024; p. 40. [Google Scholar]
- Chuard, A.; Courvoisier-Guyader, K.; Remy, S.; Spatz, S.; Denesvre, C.; Pasdeloup, D. The tegument protein pUL47 of Marek’s Disease Virus is necessary for horizontal transmission and is important for expression of glycoprotein gC. J. Virol. 2020, 95, e01645-20. [Google Scholar] [CrossRef]
- Hernandez, R.; Brown, D.T. Growth and maintenance of chick embryo fibroblasts (CEF). Curr. Protoc. Microbiol. 2010, 17, A-4I. [Google Scholar] [CrossRef]
- Dorange, F.; El Mehdaoui, S.; Pichon, C.; Coursaget, P.; Vautherot, J.F. Marek’s disease virus (MDV) homologues of herpes simplex virus type 1 UL49 (VP22) and UL48 (VP16) genes: High-level expression and characterization of MDV-1 VP22 and VP16. J. Gen. Virol. 2000, 81, 2219–2230. [Google Scholar] [CrossRef]
- Morgan, R.W.; Cantello, J.L.; McDermott, C.H. Transfection of chicken embryo fibroblasts with Marek’s disease virus DNA. Avian Dis. 1990, 34, 345–351. [Google Scholar] [CrossRef]
- Sinzger, C.; Knapp, J.; Schmidt, K.; Kahl, M.; Jahn, G. A simple and rapid method for preparation of viral DNA from cell associated cytomegalovirus. J. Virol. Methods 1999, 81, 115–122. [Google Scholar] [CrossRef]
- Denesvre, C.; Blondeau, C.; Lemesle, M.; Le Vern, Y.; Vautherot, D.; Roingeard, P.; Vautherot, J.F. Morphogenesis of a highly replicative EGFPVP22 recombinant Marek’s disease virus (MDV) in cell culture. J. Virol. 2007, 81, 12348–12359. [Google Scholar] [CrossRef]
- Richerioux, N.; Blondeau, C.; Wiedemann, A.; Remy, S.; Vautherot, J.F.; Denesvre, C. Rho-ROCK and Rac-PAK signaling pathways have opposing effects on the cell-to-cell spread of Marek’s Disease Virus. PLoS ONE 2012, 7, e44072. [Google Scholar] [CrossRef]
- Berthault, C.; Larcher, T.; Hartle, S.; Vautherot, J.F.; Trapp-Fragnet, L.; Denesvre, C. Atrophy of primary lymphoid organs induced by Marek’s disease virus during early infection is associated with increased apoptosis, inhibition of cell proliferation and a severe B-lymphopenia. Vet. Res. 2018, 49, 31. [Google Scholar] [CrossRef]
- Rémy, S.; Le Pape, G.; Gourichon, D.; Gardin, Y.; Denesvre, C. Chickens can durably clear herpesvirus vaccine infection in feathers while still carrying vaccine-induced antibodies. Vet. Res. 2020, 51, 24. [Google Scholar] [CrossRef]
- Jarosinski, K.W.; Yunis, R.; O’Connell, P.H.; Markowski-Grimsrud, C.J.; Schat, K.A. Influence of genetic resistance of the chicken and virulence of Marek’s disease virus (MDV) on nitric oxide responses after MDV infection. Avian Dis. 2002, 46, 636–649. [Google Scholar] [CrossRef]
- Noguchi, K.; Gel, Y.R.; Brunner, E.; Konietschke, F. nparLD: An R Software Package for the Nonparametric Analysis of Longitudinal Data in Factorial Experiments. J. Stat. Softw. 2012, 50, 1–23. [Google Scholar] [CrossRef]
- Brunner, E.; Domhof, S.; Langer, F. Nonparametric Analysis of Longitudinal Data in Factorial Experiments; Wiley: New York, NY, USA, 2002. [Google Scholar]
- Singh, S.M.; Baigent, S.J.; Petherbridge, L.J.; Smith, L.P.; Nair, V.K. Comparative efficacy of BAC-derived recombinant SB-1 vaccine and the parent wild type strain in preventing replication, shedding and disease induced by virulent Marek’s disease virus. Res. Vet. Sci. 2010, 89, 140–145. [Google Scholar] [CrossRef]
- Che, X.; Reichelt, M.; Sommer, M.H.; Rajamani, J.; Zerboni, L.; Arvin, A.M. Functions of the ORF9-to-ORF12 gene cluster in varicella-zoster virus replication and in the pathogenesis of skin infection. J. Virol. 2008, 82, 5825–5834. [Google Scholar] [CrossRef]
- Helferich, D.; Veits, J.; Teifke, J.P.; Mettenleiter, T.C.; Fuchs, W. The UL47 gene of avian infectious laryngotracheitis virus is not essential for in vitro replication but is relevant for virulence in chickens. J. Gen. Virol. 2007, 88, 732–742. [Google Scholar] [CrossRef]
- Lobanov, V.A.; Maher-Sturgess, S.L.; Snider, M.G.; Lawman, Z.; Babiuk, L.A.; van Drunen Littel-van den Hurk, S. A UL47 gene deletion mutant of bovine herpesvirus type 1 exhibits impaired growth in cell culture and lack of virulence in cattle. J. Virol. 2010, 84, 445–458. [Google Scholar] [CrossRef]
- Kopp, M.; Klupp, B.G.; Granzow, H.; Fuchs, W.; Mettenleiter, T.C. Identification and characterization of the pseudorabies virus tegument proteins UL46 and UL47: Role for UL47 in virion morphogenesis in the cytoplasm. J. Virol. 2002, 76, 8820–8833. [Google Scholar] [CrossRef]
- Liu, Z.; Kato, A.; Shindo, K.; Noda, T.; Sagara, H.; Kawaoka, Y.; Arii, J.; Kawaguchi, Y. Herpes simplex virus 1 UL47 interacts with viral nuclear egress factors UL31, UL34, and Us3 and regulates viral nuclear egress. J. Virol. 2014, 88, 4657–4667. [Google Scholar] [CrossRef]
- Donnelly, M.; Verhagen, J.; Elliott, G. RNA binding by the herpes simplex virus type 1 nucleocytoplasmic shuttling protein UL47 is mediated by an N-terminal arginine-rich domain that also functions as its nuclear localization signal. J. Virol. 2007, 81, 2283–2296. [Google Scholar] [CrossRef]
- Islam, A.; Schulz, S.; Afroz, S.; Babiuk, L.A.; van Drunen Littel-van den Hurk, S. Interaction of VP8 with mRNAs of bovine herpesvirus-1. Virus Res. 2015, 197, 116–126. [Google Scholar] [CrossRef]
- Denesvre, C.; You, Y.; Remy, S.; Vychodil, T.; Courvoisier, K.; Penzes, Z.; Bertzbach, L.D.; Kheimar, A.; Kaufer, B.B. Impact of viral telomeric repeat sequences on herpesvirus vector vaccine integration and persistence. PLoS Pathog. 2024, 20, e1012261. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Target Gene | Sequence (5′ to 3′) | Amplicon Size | |
|---|---|---|---|
| SB1UL48-F2 | for | CTGCTGCTCGCACAATGTAA | 2894 kbp with UL47 488 bp wo UL47 |
| SB1UL46-R2 | rev | GCTAGCGCAAGCATATCG | |
| SB1UL47-F1 | for | GAAAGTATTGCCCCGGGTAT | 351 bp |
| SB1UL47-R1 | rev | CTCATTTAATCGCGACAGCA | |
| SB1 qPCR | probe | FAM-CCCGGGTCGCCTCATCTGGA-TAMRA | 67 bp |
| for | AGCATGCGGGAAGAAAAGAG | ||
| rev | GCGCCGAAAAGCTAGAAAAG | ||
| iNos qPCR | probe | FAM-CTCTGCCTGCTGTTGCCAACATGC-TAMRA | 81 bp |
| for | GAGTGGTTTAAGGAGTTGGATCTGA | ||
| rev | TTCCAGACCTCCCACCTCAA | ||
| SB1UL51-F | for | CTGTACATTCTAGCCAACGACTG | 384 bp |
| SB1UL51-R | rev | GCCCGGATCCCAGTGGCTGATAAC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Esaki, M.; Chollot, M.; Rémy, S.; Courvoisier-Guyader, K.; Penzes, Z.; Pasdeloup, D.; Denesvre, C. Tegument Protein pUL47 Is Important but Not Essential for Horizontal Transmission of Vaccinal Strain SB-1 of Gallid Alphaherpesvirus 3. Viruses 2025, 17, 431. https://doi.org/10.3390/v17030431
Esaki M, Chollot M, Rémy S, Courvoisier-Guyader K, Penzes Z, Pasdeloup D, Denesvre C. Tegument Protein pUL47 Is Important but Not Essential for Horizontal Transmission of Vaccinal Strain SB-1 of Gallid Alphaherpesvirus 3. Viruses. 2025; 17(3):431. https://doi.org/10.3390/v17030431
Chicago/Turabian StyleEsaki, Motoyuki, Mélanie Chollot, Sylvie Rémy, Katia Courvoisier-Guyader, Zoltan Penzes, David Pasdeloup, and Caroline Denesvre. 2025. "Tegument Protein pUL47 Is Important but Not Essential for Horizontal Transmission of Vaccinal Strain SB-1 of Gallid Alphaherpesvirus 3" Viruses 17, no. 3: 431. https://doi.org/10.3390/v17030431
APA StyleEsaki, M., Chollot, M., Rémy, S., Courvoisier-Guyader, K., Penzes, Z., Pasdeloup, D., & Denesvre, C. (2025). Tegument Protein pUL47 Is Important but Not Essential for Horizontal Transmission of Vaccinal Strain SB-1 of Gallid Alphaherpesvirus 3. Viruses, 17(3), 431. https://doi.org/10.3390/v17030431