
Testing the Tenacity of Small Ruminant Lentiviruses In Vitro to Assess the Potential Risk of Indirect Fomites’ Transmission



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Virus
2.2. Nutrient Medium
2.3. Drying Medium
2.4. Cells
2.5. Virus Stored in Liquid
2.6. Virus Dried, Stored, and Resuspended
2.7. Statistical Analysis
3. Results
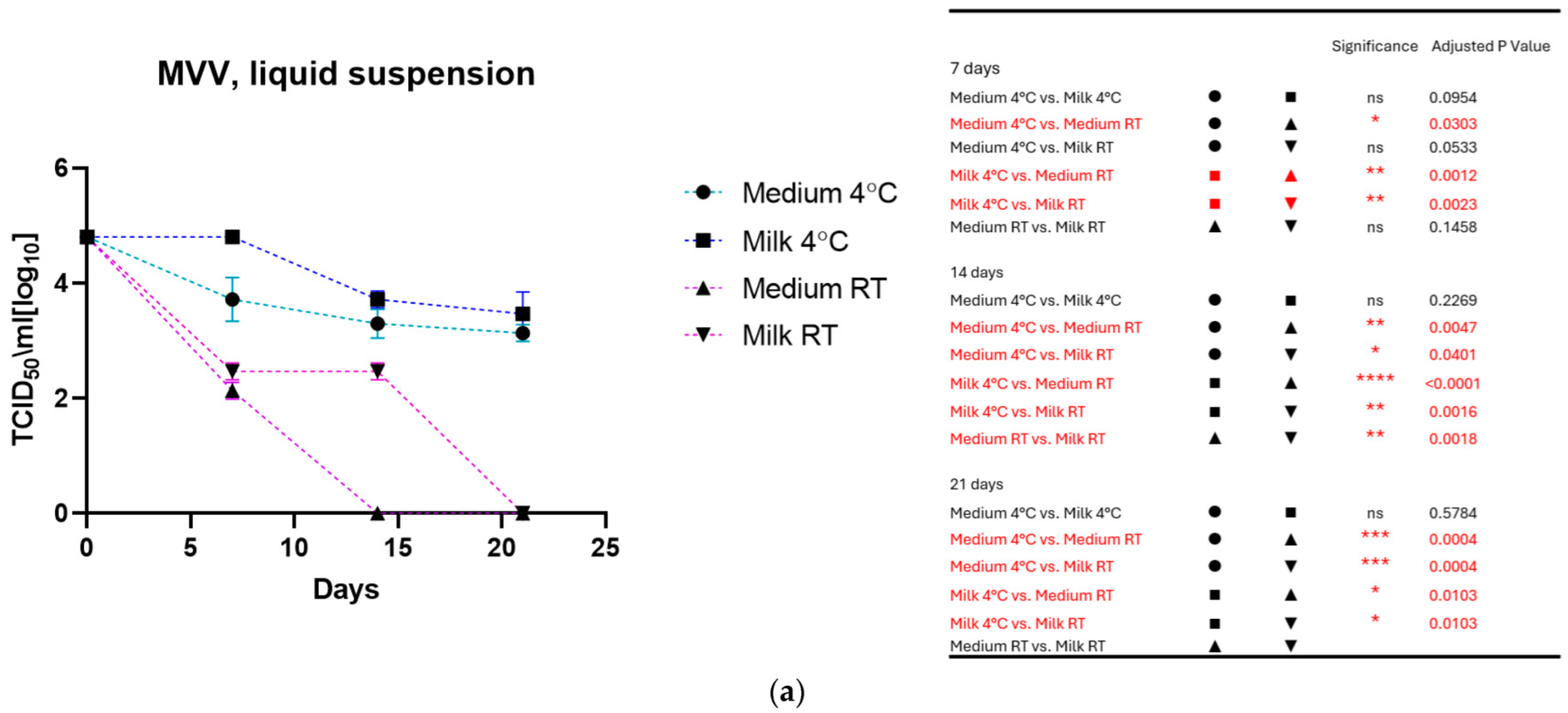
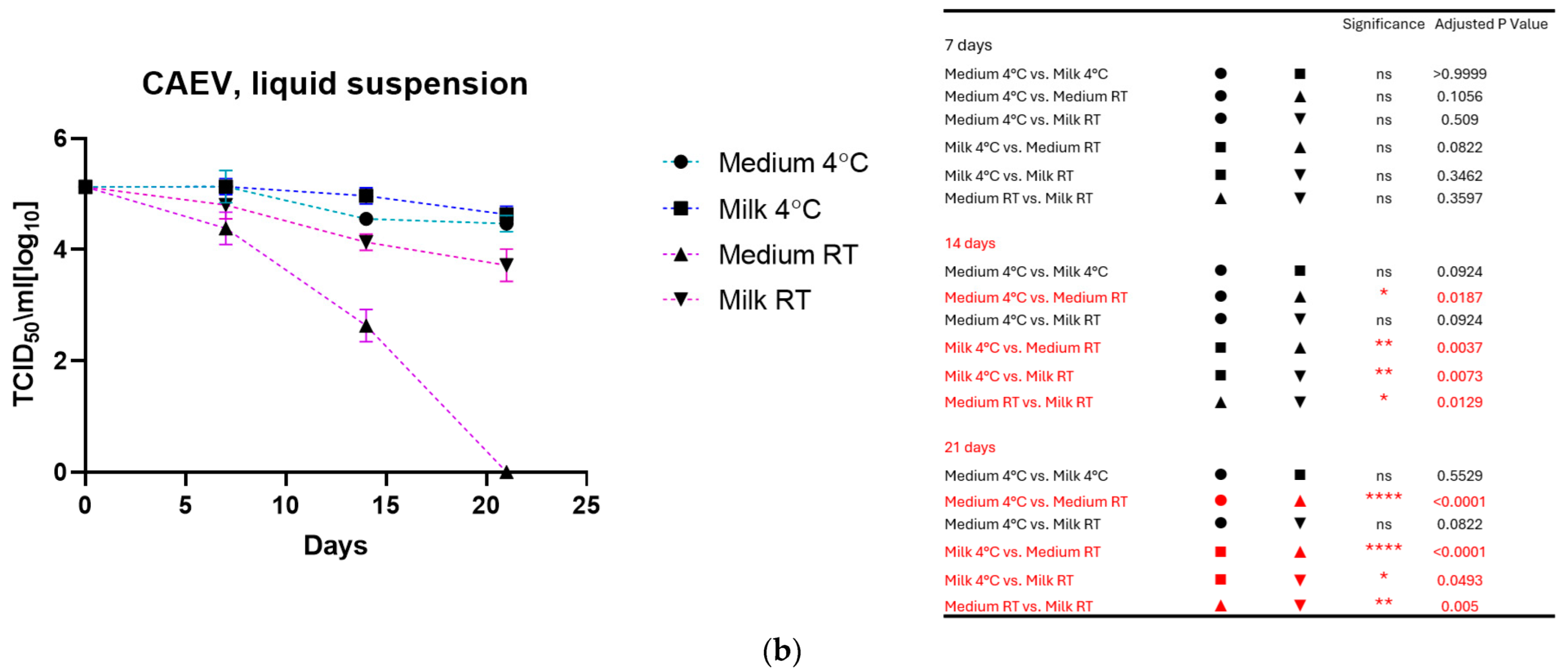
3.1. Virus Stored in Liquid
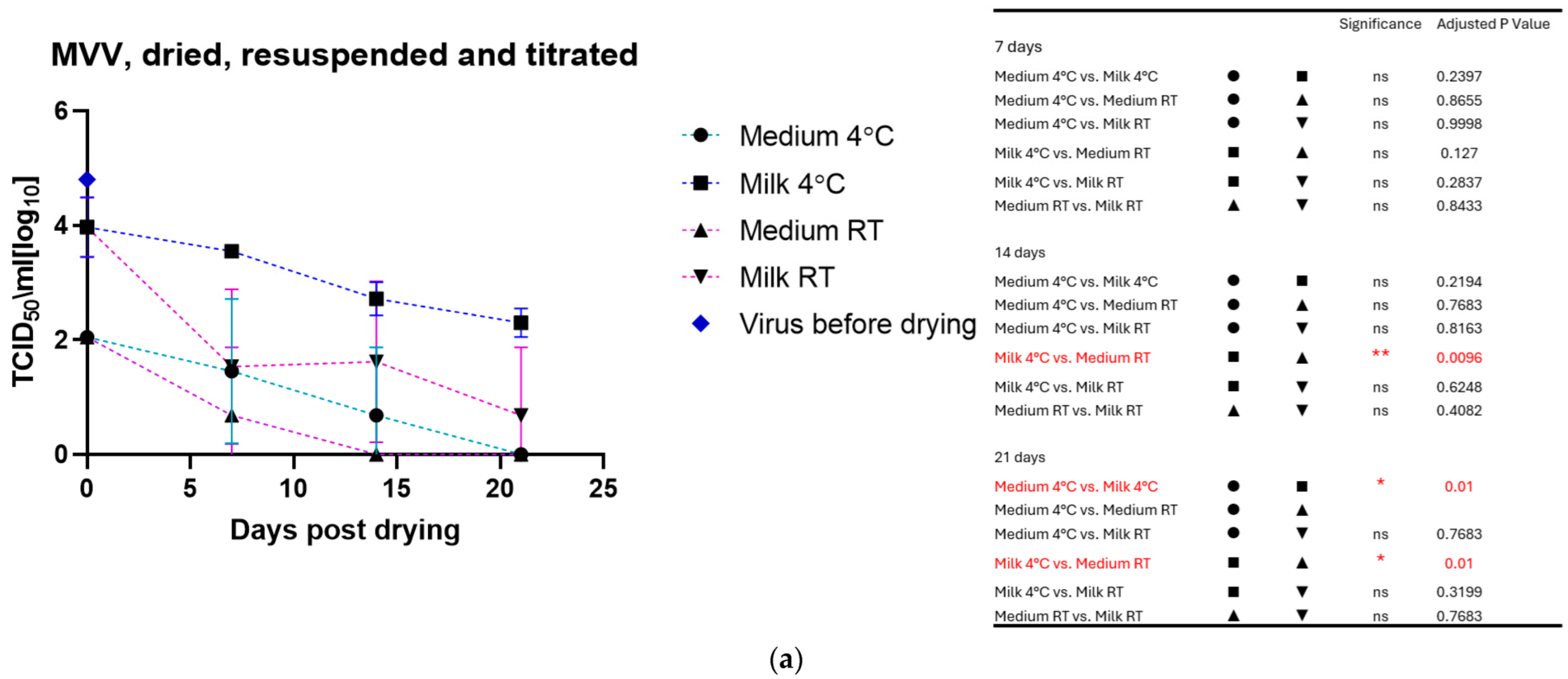
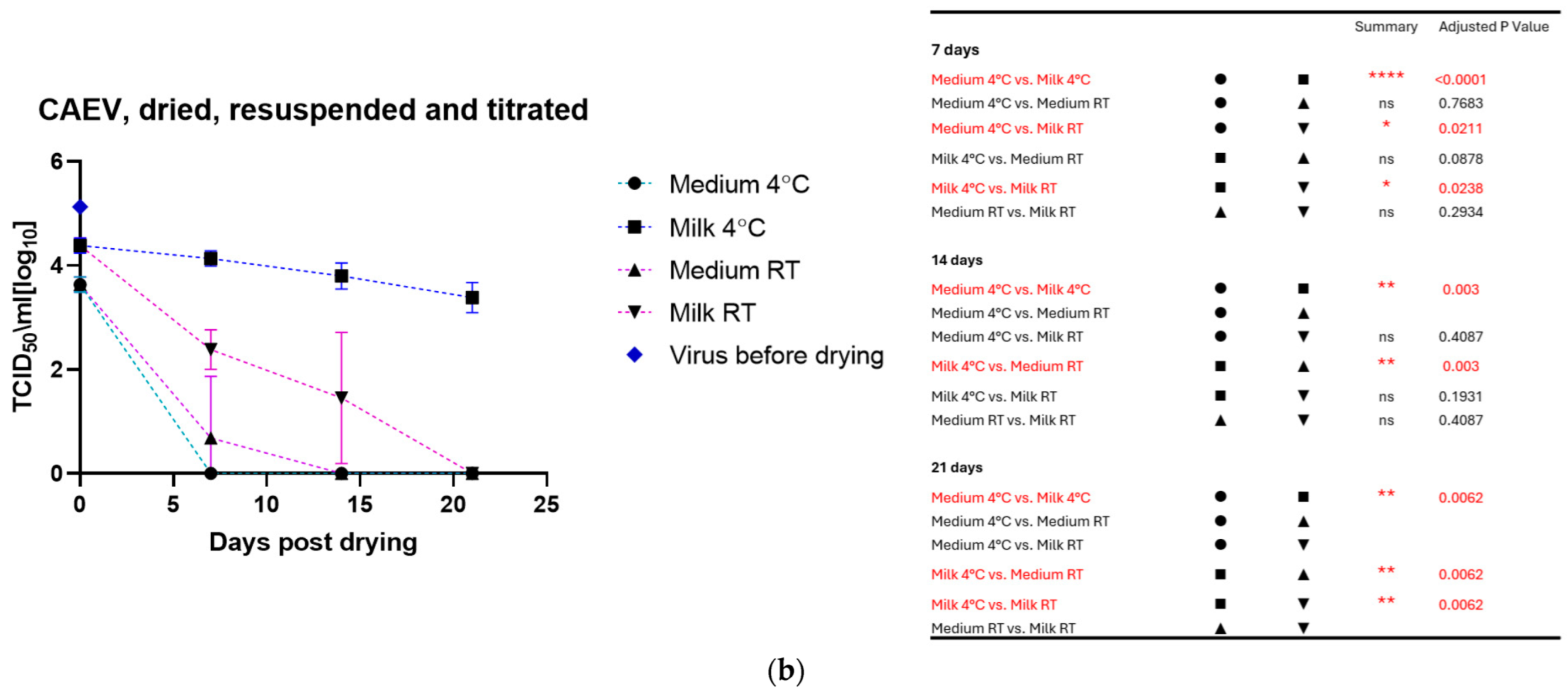
3.2. Virus: Dried, Stored, and Resuspended
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| SRLV | Small ruminant lentiviruses |
| MVV | Maedi-visna virus |
| CAEV | Caprine arthritis encephalitis virus |
| LSM | Lamb synovial membrane cells |
| GSM | Goat synovial membrane cells |
| FBS | Fetal bovine serum |
| RT | Room temperature |
References
- Olech, M.; Kuzmak, J.; Kycko, A.; Junkuszew, A. Phylogenetic analysis of small ruminant lentiviruses originating from naturally infected sheep and goats from poland based on the long terminal repeat sequences. J. Vet. Res. 2022, 66, 497–510. [Google Scholar] [CrossRef]
- Michiels, R.; Adjadj, N.R.; De Regge, N. Phylogenetic analysis of belgian small ruminant lentiviruses supports cross species virus transmission and identifies new subtype b5 strains. Pathogens 2020, 9, 183. [Google Scholar] [CrossRef] [PubMed]
- Kuhar, U.; Barlic-Maganja, D.; Zadnik, T.; Grom, J. Molecular and genetic characteristics of small ruminant lentiviruses in slovenia. Acta Vet. Hung. 2013, 61, 135–146. [Google Scholar] [CrossRef] [PubMed]
- L’Homme, Y.; Ouardani, M.; Levesque, V.; Bertoni, G.; Simard, C.; Pisoni, G. Molecular characterization and phylogenetic analysis of small ruminant lentiviruses isolated from Canadian sheep and goats. Virol. J. 2011, 8, 271. [Google Scholar] [CrossRef]
- Leroux, C.; Cruz, J.C.; Mornex, J.F. Srlvs: A genetic continuum of lentiviral species in sheep and goats with cumulative evidence of cross species transmission. Curr. HIV Res. 2010, 8, 94–100. [Google Scholar]
- Reina, R.; Mora, M.I.; Glaria, I.; Garcia, I.; Solano, C.; Lujan, L.; Badiola, J.J.; Contreras, A.; Berriatua, E.; Juste, R.; et al. Molecular characterization and phylogenetic study of Maedi visna and caprine arthritis encephalitis viral sequences in sheep and goats from spain. Virus Res. 2006, 121, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Gjerset, B.; Storset, A.K.; Rimstad, E. Genetic diversity of small-ruminant lentiviruses: Characterization of norwegian isolates of caprine arthritis encephalitis virus. J. Gen. Virol. 2006, 87, 573–580. [Google Scholar] [CrossRef]
- Zanoni, R.G. Phylogenetic analysis of small ruminant lentiviruses. J. Gen. Virol. 1998, 79, 1951–1961. [Google Scholar] [CrossRef]
- Valas, S.; Benoit, C.; Guionaud, C.; Perrin, G.; Mamoun, R.Z. North american and french caprine arthritis-encephalitis viruses emerge from ovine maedi-visna viruses. Virology 1997, 237, 307–318. [Google Scholar] [CrossRef]
- Gazit, A.; Yaniv, A.; Dvir, M.; Perk, K.; Irving, S.G.; Dahlberg, J.E. The caprine arthritis-encephalitis virus is a distinct virus within the lentivirus group. Virology 1983, 124, 192–195. [Google Scholar] [CrossRef]
- Shah, C.; Boni, J.; Huder, J.B.; Vogt, H.R.; Muhlherr, J.; Zanoni, R.; Miserez, R.; Lutz, H.; Schupbach, J. Phylogenetic analysis and reclassification of caprine and ovine lentiviruses based on 104 new isolates: Evidence for regular sheep-to-goat transmission and worldwide propagation through livestock trade. Virology 2004, 319, 12–26. [Google Scholar] [CrossRef]
- Kalogianni, A.I.; Bossis, I.; Ekateriniadou, L.V.; Gelasakis, A.I. Etiology, epizootiology and control of maedi-visna in dairy sheep: A review. Animals 2020, 10, 616. [Google Scholar] [CrossRef] [PubMed]
- de Miguel, R.; Arrieta, M.; Rodriguez-Largo, A.; Echeverria, I.; Resendiz, R.; Perez, E.; Ruiz, H.; Perez, M.; de Andres, D.; Reina, R.; et al. Worldwide prevalence of small ruminant lentiviruses in sheep: A systematic review and meta-analysis. Animals 2021, 11, 784. [Google Scholar] [CrossRef]
- Sigurdsson, B.; Grimsson, H.; Palsson, P.A. Maedi, a chronic, progressive infection of sheep’s lungs. J. Infect. Dis. 1952, 90, 233–241. [Google Scholar] [CrossRef]
- Carrozza, M.L.; Niewiadomska, A.M.; Mazzei, M.; Abi-Said, M.R.; Hue, S.; Hughes, J.; Gatseva, A.; Gifford, R.J. Emergence and pandemic spread of small ruminant lentiviruses. Virus Evol. 2023, 9, vead005. [Google Scholar] [CrossRef]
- Molaee, V.; Bazzucchi, M.; De Mia, G.M.; Otarod, V.; Abdollahi, D.; Rosati, S.; Luhken, G. Phylogenetic analysis of small ruminant lentiviruses in Germany and Iran suggests their expansion with domestic sheep. Sci. Rep. 2020, 10, 2243. [Google Scholar] [CrossRef] [PubMed]
- Pétursson, G. Experience with visna virus in Iceland. Ann. N. Y. Acad. Sci. 1994, 724, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Palsson, P.A. Maedi-visna. J. Clin. Pathol. Suppl. (R Coll Pathol.) 1972, 6, 115–120. [Google Scholar] [CrossRef]
- Zink, M.C.; Johnson, L.K. Pathobiology of lentivirus infections of sheep and goats. Virus Res. 1994, 32, 139–154. [Google Scholar] [CrossRef]
- Blacklaws, B.A. Small ruminant lentiviruses: Immunopathogenesis of visna-maedi and caprine arthritis and encephalitis virus. Comp. Immunol. Microbiol. Infect. Dis. 2012, 35, 259–269. [Google Scholar] [CrossRef]
- Minguijon, E.; Reina, R.; Perez, M.; Polledo, L.; Villoria, M.; Ramirez, H.; Leginagoikoa, I.; Badiola, J.J.; Garcia-Marin, J.F.; de Andres, D.; et al. Small ruminant lentivirus infections and diseases. Vet. Microbiol. 2015, 181, 75–89. [Google Scholar] [CrossRef] [PubMed]
- Nagel-Alne, G.E.; Krontveit, R.; Bohlin, J.; Valle, P.S.; Skjerve, E.; Solverod, L.S. The norwegian healthier goats program—Modeling lactation curves using a multilevel cubic spline regression model. J. Dairy Sci. 2014, 97, 4166–4173. [Google Scholar] [CrossRef] [PubMed]
- Nowicka, D.; Czopowicz, M.; Bagnicka, E.; Rzewuska, M.; Strzalkowska, N.; Kaba, J. Influence of small ruminant lentivirus infection on cheese yield in goats. J. Dairy Res. 2015, 82, 102–106. [Google Scholar] [CrossRef] [PubMed]
- Kalogianni, A.I.; Gelasakis, A.I. The impact of small ruminant lentiviruses infections on milk yield and milk quality traits in intensively reared dairy sheep. Small Rumin. Res. 2025, 243, 107424. [Google Scholar] [CrossRef]
- Blacklaws, B.A.; Berriatua, E.; Torsteinsdottir, S.; Watt, N.J.; de Andres, D.; Klein, D.; Harkiss, G.D. Transmission of small ruminant lentiviruses. Vet. Microbiol. 2004, 101, 199–208. [Google Scholar] [CrossRef]
- Alvarez, V.; Arranz, J.; Daltabuit-Test, M.; Leginagoikoa, I.; Juste, R.A.; Amorena, B.; de Andres, D.; Lujan, L.L.; Badiola, J.J.; Berriatua, E. Relative contribution of colostrum from maedi-visna virus (mvv) infected ewes to mvv-seroprevalence in lambs. Res. Vet. Sci. 2005, 78, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Illius, A.W.; Savill, N.J. Maternal transmission of small ruminant lentivirus has no epidemiological importance. Prev. Vet. Med. 2024, 230, 106297. [Google Scholar] [CrossRef]
- Thormar, H. Maedi-visna virus and its relationship to human immunodeficiency virus. AIDS Rev. 2005, 7, 233–245. [Google Scholar]
- Villoria, M.; Leginagoikoa, I.; Luján, L.; Pérez, M.; Salazar, E.; Berriatua, E.; Juste, R.A.; Minguijón, E. Detection of small ruminant lentivirus in environmental samples of air and water. Small Rumin. Res. 2013, 110, 155–160. [Google Scholar] [CrossRef]
- Cardinaux, L.; Zahno, M.L.; Deubelbeiss, M.; Zanoni, R.; Vogt, H.R.; Bertoni, G. Virological and phylogenetic characterization of attenuated small ruminant lentivirus isolates eluding efficient serological detection. Vet. Microbiol. 2013, 162, 572–581. [Google Scholar] [CrossRef] [PubMed]
- Herrmann-Hoesing, L.M.; White, S.N.; Broughton-Neiswanger, L.E.; Johnson, W.C.; Noh, S.M.; Schneider, D.A.; Li, H.; Taus, N.S.; Reynolds, J.; Truscott, T.; et al. Ovine progressive pneumonia virus is transmitted more effectively via aerosol nebulization than oral administration. Open J. Vet. Med. 2012, 2, 113–119. [Google Scholar] [CrossRef]
- Ravazzolo, A.P.; Nenci, C.; Vogt, H.-R.; Waldvogel, A.; Obexer-Ruff, G.; Peterhans, E.; Bertoni, G. Viral load, organ distribution, histopathological lesions, and cytokine mrna expression in goats infected with a molecular clone of the caprine arthritis encephalitis virus. Virology 2006, 350, 116–127. [Google Scholar] [CrossRef] [PubMed]
- Adams, D.S.; Klevjer-Anderson, P.; Carlson, J.L.; McGuire, T.C. Transmission and control of caprine arthritis-encephalitis virus. Am. J. Vet. Res. 1983, 44, 1670–1675. [Google Scholar] [CrossRef]
- Vandamme, A.M.; Van Laethem, K.; Schmit, J.C.; Van Wijngaerden, E.; Reynders, M.; Debyser, Z.; Witvrouw, M.; Van Ranst, M.; De Clercq, E.; Desmyter, J. Long-term stability of human immunodeficiency virus viral load and infectivity in whole blood. Eur. J. Clin. Investig. 1999, 29, 445–452. [Google Scholar] [CrossRef]
- van Bueren, J.; Simpson, R.A.; Jacobs, P.; Cookson, B.D. Survival of human immunodeficiency virus in suspension and dried onto surfaces. J. Clin. Microbiol. 1994, 32, 571–574. [Google Scholar] [CrossRef] [PubMed]
- Pyper, J.M.; Clements, J.E.; Gonda, M.A.; Narayan, O. Sequence homology between cloned caprine arthritis encephalitis virus and visna virus, two neurotropic lentiviruses. J. Virol. 1986, 58, 665–700. [Google Scholar] [CrossRef]
- Turelli, P.; Guiguen, F.; Mornex, J.F.; Vigne, R.; Querat, G. Dutpase-minus caprine arthritis-encephalitis virus is attenuated for pathogenesis and accumulates g-to-a substitutions. J. Virol. 1997, 71, 4522–4530. [Google Scholar] [CrossRef] [PubMed]
- Saltarelli, M.; Querat, G.; Konings, D.A.; Vigne, R.; Clements, J.E. Nucleotide sequence and transcriptional analysis of molecular clones of caev which generate infectious virus. Virology 1990, 179, 347–364. [Google Scholar] [CrossRef]
- Staskus, K.A.; Retzel, E.F.; Lewis, E.D.; Silsby, J.L.; St Cyr, S.; Rank, J.M.; Wietgrefe, S.W.; Haase, A.T.; Cook, R.; Fast, D.; et al. Isolation of replication-competent molecular clones of visna virus. Virology 1991, 181, 228–240. [Google Scholar] [CrossRef]
- De Martin, E.; Golomingi, A.; Zahno, M.; Cachim, J.; Di Labio, E.; Perler, L.; Abril, C.; Zanoni, R.; Bertoni, G. Diagnostic response to a cross-border challenge for the swiss caprine arthritis encephalitis virus eradication program. Schweiz. Arch. Tierheilkd. 2019, 161, 93–104. [Google Scholar] [CrossRef]
- Ramakrishnan, M.A. Determination of 50% endpoint titer using a simple formula. World J. Virol. 2016, 5, 85–86. [Google Scholar] [CrossRef] [PubMed]
- Kramer, A.; Lexow, F.; Bludau, A.; Koster, A.M.; Misailovski, M.; Seifert, U.; Eggers, M.; Rutala, W.; Dancer, S.J.; Scheithauer, S. How long do bacteria, fungi, protozoa, and viruses retain their replication capacity on inanimate surfaces? A systematic review examining environmental resilience versus healthcare-associated infection risk by “fomite-borne risk assessment”. Clin. Microbiol. Rev. 2024, 37, e0018623. [Google Scholar] [CrossRef] [PubMed]
- Blatti-Cardinaux, L.; Pisoni, G.; Stoffel, M.H.; Zanoni, R.; Zahno, M.L.; Bertoni, G. Generation of a molecular clone of an attenuated lentivirus, a first step in understanding cytopathogenicity and virulence. Virology 2016, 487, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Minardi da Cruz, J.C.; Singh, D.K.; Lamara, A.; Chebloune, Y. Small ruminant lentiviruses (srlvs) break the species barrier to acquire new host range. Viruses 2013, 5, 1867–1884. [Google Scholar] [CrossRef]
- Torsteinsdottir, S.; Matthiasdottir, S.; Vidarsdottir, N.; Svansson, V.; Petursson, G. Intratracheal inoculation as an efficient route of experimental infection with maedi-visna virus. Res. Vet. Sci. 2003, 75, 245–247. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Samoilenko, M.; Nedosekov, V.; Bertoni, G. Testing the Tenacity of Small Ruminant Lentiviruses In Vitro to Assess the Potential Risk of Indirect Fomites’ Transmission. Viruses 2025, 17, 419. https://doi.org/10.3390/v17030419
Samoilenko M, Nedosekov V, Bertoni G. Testing the Tenacity of Small Ruminant Lentiviruses In Vitro to Assess the Potential Risk of Indirect Fomites’ Transmission. Viruses. 2025; 17(3):419. https://doi.org/10.3390/v17030419
Chicago/Turabian StyleSamoilenko, Maksym, Vitalii Nedosekov, and Giuseppe Bertoni. 2025. "Testing the Tenacity of Small Ruminant Lentiviruses In Vitro to Assess the Potential Risk of Indirect Fomites’ Transmission" Viruses 17, no. 3: 419. https://doi.org/10.3390/v17030419
APA StyleSamoilenko, M., Nedosekov, V., & Bertoni, G. (2025). Testing the Tenacity of Small Ruminant Lentiviruses In Vitro to Assess the Potential Risk of Indirect Fomites’ Transmission. Viruses, 17(3), 419. https://doi.org/10.3390/v17030419