
Development of Chimera AMP–Endolysin with Wider Spectra Against Gram-Negative Bacteria Using High-Throughput Assay

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Media, and Growth Conditions
2.2. DNA Extraction and Genome Sequencing
2.3. Identification of Endolysin Genes from Microbial Genome Data
2.4. Construction of the Endolysin Library
2.5. Endolysin Expression and Lysate Preparation
2.6. Plate Lysis Assay
2.7. Growth Inhibition Assay
2.8. Expression and Purification of Endolysins
2.9. Endolysin Activity Assay
2.10. Cytotoxicity Assay
2.11. Biofilm Assay
3. Results
3.1. Identification of Endolysin Genes from Genomes of Gram-Negative Bacteria
3.2. Screening of Chimeric Endolysin Libraries
3.3. Identification of Active Chimeric Endolysin Against A. baumannii
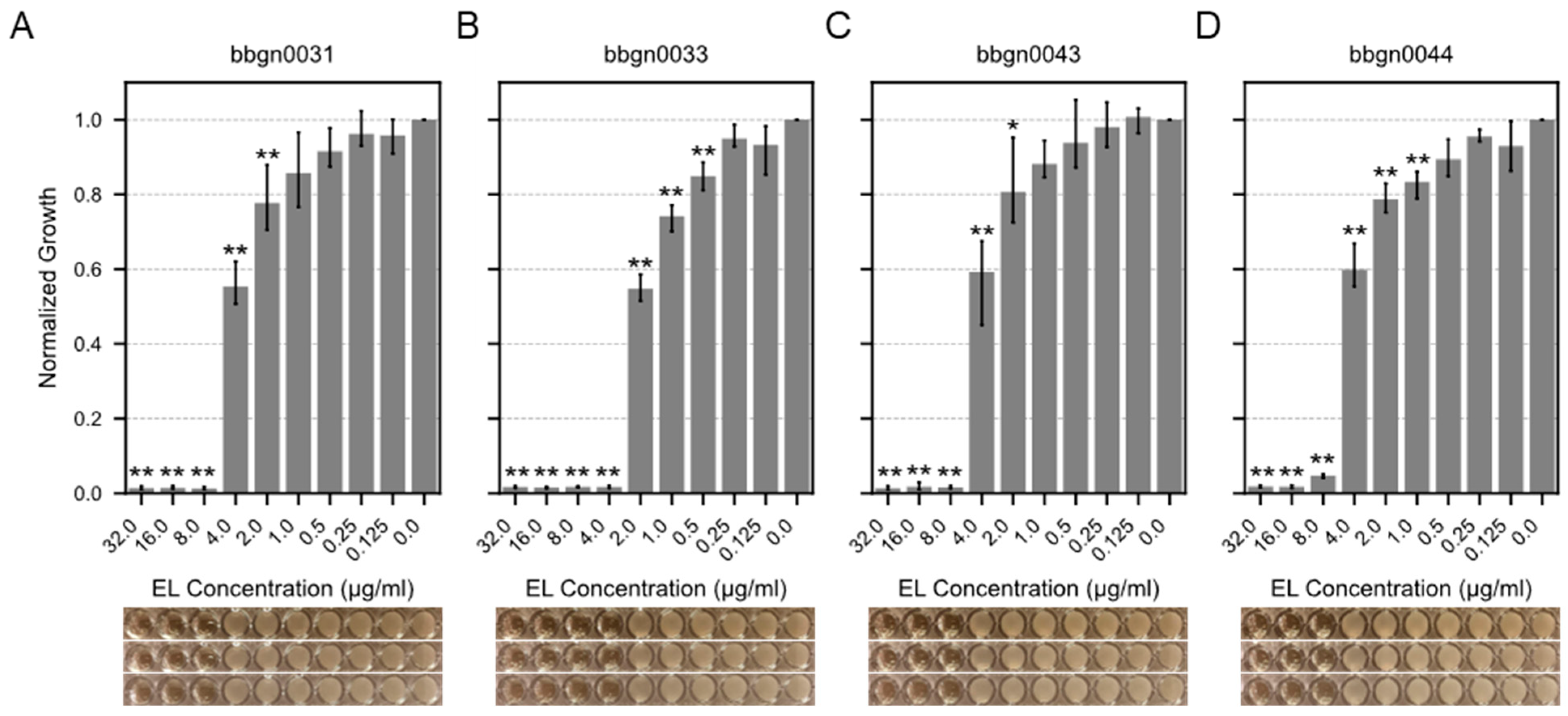
3.4. Antibacterial Spectrum of Chimera AMP–Endolysins
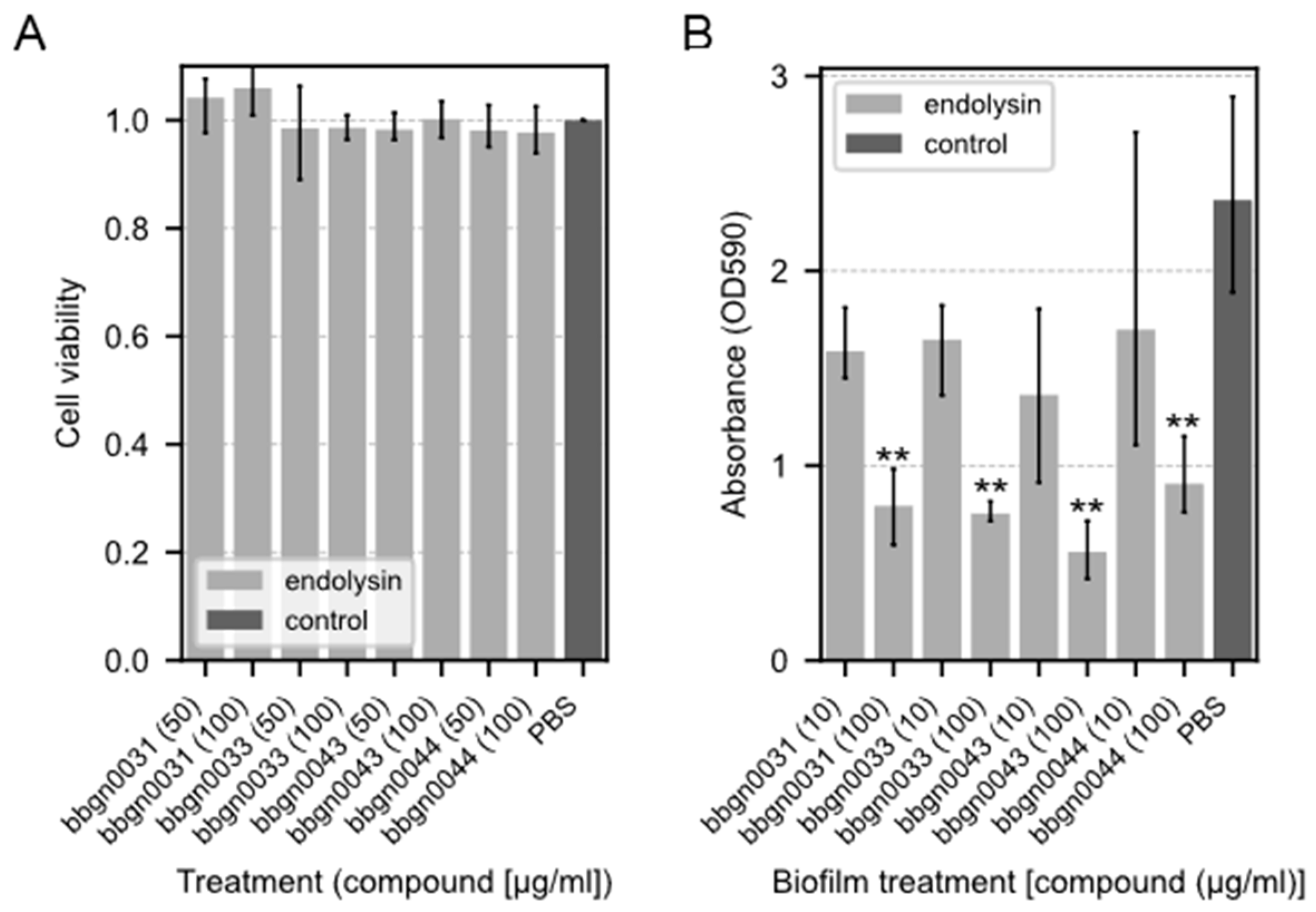
3.5. Cytotoxicity Assay of Chimera AMP–Endolysins
3.6. Biofilm Degradation Assay of Chimera AMP–Endolysins
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nikaido, H. Multidrug Resistance in Bacteria. Annu. Rev. Biochem. 2009, 78, 119. [Google Scholar] [CrossRef]
- Prestinaci, F.; Pezzotti, P.; Pantosti, A. Antimicrobial resistance: A global multifaceted phenomenon. Pathog. Glob. Health 2015, 109, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Breijyeh, Z.; Jubeh, B.; Karaman, R. Resistance of Gram-Negative Bacteria to Current Antibacterial Agents and Approaches to Resolve It. Molecules 2020, 25, 1340. [Google Scholar] [CrossRef]
- De Oliveira, D.M.P.; Forde, B.M.; Kidd, T.J.; Harris, P.N.A.; Schembri, M.A.; Beatson, S.A.; Paterson, D.L.; Walker, M.J. Antimicrobial Resistance in ESKAPE Pathogens. Clin. Microbiol. Rev. 2020, 33, e00181-19. [Google Scholar] [CrossRef] [PubMed]
- Imran, M.; Ahmad, M.N.; Dasgupta, A.; Rana, P.; Srinivas, N.; Chopra, S. Novel Approaches for the Treatment of Infections Due to Multidrug-Resistant Bacterial Pathogens. Future Med. Chem. 2022, 14, 1133–1148. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Rubio, L.; Gutiérrez, D.; Donovan, D.M.; Martínez, B.; Rodríguez, A.; García, P. Phage lytic proteins: Biotechnological applications beyond clinical antimicrobials. Crit. Rev. Biotechnol. 2016, 36, 542–552. [Google Scholar] [CrossRef] [PubMed]
- Schmelcher, M.; Donovan, D.M.; Loessner, M.J. Bacteriophage Endolysins as Novel Antimicrobials. Future Microbiol. 2012, 7, 1147–1171. [Google Scholar] [CrossRef] [PubMed]
- Sui, B.; Wang, X.; Zhao, T.; Zhen, J.; Ren, H.; Liu, W.; Zhang, X.; Zhang, C. Design, Screening, and Characterization of Engineered Phage Endolysins with Extracellular Antibacterial Activity against Gram-Negative Bacteria. Appl. Environ. Microbiol. 2023, 89, e00581-23. [Google Scholar] [CrossRef]
- Schmelcher, M.; Loessner, M.J. Bacteriophage endolysins: Applications for food safety. Curr. Opin. Biotechnol. 2016, 37, 76–87. [Google Scholar] [CrossRef]
- Aslam, B.; Arshad, M.I.; Aslam, M.A.; Muzammil, S.; Siddique, A.B.; Yasmeen, N.; Khurshid, M.; Rasool, M.; Ahmad, M.; Rasool, M.H.; et al. Bacteriophage Proteome: Insights and Potentials of an Alternate to Antibiotics. Infect. Dis. Ther. 2021, 10, 1171–1193. [Google Scholar] [CrossRef]
- Abdelrahman, F.; Easwaran, M.; Daramola, O.I.; Ragab, S.; Lynch, S.; Oduselu, T.J.; Khan, F.M.; Ayobami, A.; Adnan, F.; Torrents, E.; et al. Phage-Encoded Endolysins. Antibiotics 2021, 10, 124. [Google Scholar] [CrossRef]
- Abdurahman, M.A.; Durukan, İ.; Dinçer, T.; Pektaş, S.; Karataş, E.; Kiliç, A.O. Staphylococcus aureus Bacteriophage 52 Endolysin Exhibits Anti-Biofilm and Broad Antibacterial Activity Against Gram-Positive Bacteria. Protein J. 2023, 42, 596–606. [Google Scholar] [CrossRef]
- Kim, S.-H.; Lee, H.; Park, M.-K. Isolation, characterization, and application of a novel, lytic phage vB_SalA_KFSST3 with depolymerase for the control of Salmonella and its biofilm on cantaloupe under cold temperature. Food Res. Int. 2023, 172, 113062. [Google Scholar] [CrossRef]
- São-José, C. Engineering of Phage-Derived Lytic Enzymes: Improving Their Potential as Antimicrobials. Antibiotics 2018, 7, 29. [Google Scholar] [CrossRef] [PubMed]
- Heselpoth, R.D.; Yin, Y.; Moult, J.; Nelson, D.C. Increasing the stability of the bacteriophage endolysin PlyC using rationale-based FoldX computational modeling. Protein Eng. Des. Sel. 2015, 28, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.M.; Kim, D.; Kim, K.; Park, S.-J.; Akter, S.; Kim, J.; Bang, S.; Kim, S.; Kim, J.; Lee, J.C.; et al. Engineering of lysin by fusion of antimicrobial peptide (cecropin A) enhances its antibacterial properties against multidrug-resistant Acinetobacter baumannii. Front. Microbiol. 2022, 13, 988522. [Google Scholar] [CrossRef]
- Vander Elst, N.; Bert, J.; Favoreel, H.; Lavigne, R.; Meyer, E.; Briers, Y. Development of engineered endolysins with in vitro intracellular activity against streptococcal bovine mastitis-causing pathogens. Microb. Biotechnol. 2023, 16, 2367–2386. [Google Scholar] [CrossRef]
- Gontijo, M.T.P.; Jorge, G.P.; Brocchi, M. Current Status of Endolysin-Based Treatments against Gram-Negative Bacteria. Antibiotics 2021, 10, 1143. [Google Scholar] [CrossRef] [PubMed]
- Love, M.J.; Abeysekera, G.S.; Muscroft-Taylor, A.C.; Billington, C.; Dobson, R.C.J. On the catalytic mechanism of bacteriophage endolysins: Opportunities for engineering. Biochim. Biophys. Acta BBA-Proteins Proteom. 2020, 1868, 140302. [Google Scholar] [CrossRef]
- Briers, Y.; Walmagh, M.; Grymonprez, B.; Biebl, M.; Pirnay, J.-P.; Defraine, V.; Michiels, J.; Cenens, W.; Aertsen, A.; Miller, S.; et al. Art-175 Is a Highly Efficient Antibacterial against Multidrug-Resistant Strains and Persisters of Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2014, 58, 3774–3784. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Zheng, Y.; Dai, J.; Zhou, J.; Yu, R.; Zhang, C. Design SMAP29-LysPA26 as a Highly Efficient Artilysin against Pseudomonas aeruginosa with Bactericidal and Antibiofilm Activity. Microbiol. Spectr. 2021, 9, e00546-21. [Google Scholar] [CrossRef]
- Xu, D.; Zhao, S.; Dou, J.; Xu, X.; Zhi, Y.; Wen, L. Engineered endolysin-based “artilysins” for controlling the gram-negative pathogen Helicobacter pylori. AMB Express 2021, 11, 63. [Google Scholar] [CrossRef]
- Gouveia, A.; Pinto, D.; Veiga, H.; Antunes, W.; Pinho, M.G.; São-José, C. Synthetic antimicrobial peptides as enhancers of the bacteriolytic action of staphylococcal phage endolysins. Sci. Rep. 2022, 12, 1245. [Google Scholar] [CrossRef] [PubMed]
- Schleifer, K.H.; Kandler, O. Peptidoglycan types of bacterial cell walls and their taxonomic implications. Bacteriol. Rev. 1972, 36, 407–477. [Google Scholar] [CrossRef]
- Kogawa, M.; Nishikawa, Y.; Saeki, T.; Yoda, T.; Arikawa, K.; Takeyama, H.; Hosokawa, M. Revealing within-species diversity in uncultured human gut bacteria with single-cell long-read sequencing. Front. Microbiol. 2023, 14, 1133917. [Google Scholar] [CrossRef] [PubMed]
- Ide, K.; Saeki, T.; Arikawa, K.; Yoda, T.; Endoh, T.; Matsuhashi, A.; Takeyama, H.; Hosokawa, M. Exploring strain diversity of dominant human skin bacterial species using single-cell genome sequencing. Front. Microbiol. 2022, 13, 955404. [Google Scholar] [CrossRef] [PubMed]
- Sirén, K.; Millard, A.; Petersen, B.; Gilbert, M.T.P.; Clokie, M.R.J.; Sicheritz-Pontén, T. Rapid discovery of novel prophages using biological feature engineering and machine learning. NAR Genom. Bioinforma. 2021, 3, lqaa109. [Google Scholar] [CrossRef]
- McNair, K.; Zhou, C.; Dinsdale, E.A.; Souza, B.; Edwards, R.A. PHANOTATE: A novel approach to gene identification in phage genomes. Bioinformatics 2019, 35, 4537–4542. [Google Scholar] [CrossRef] [PubMed]
- Buchfink, B.; Reuter, K.; Drost, H.-G. Sensitive protein alignments at tree-of-life scale using DIAMOND. Nat. Methods 2021, 18, 366–368. [Google Scholar] [CrossRef] [PubMed]
- Quevillon, E.; Silventoinen, V.; Pillai, S.; Harte, N.; Mulder, N.; Apweiler, R.; Lopez, R. InterProScan: Protein domains identifier. Nucleic Acids Res. 2005, 33, W116. [Google Scholar] [CrossRef]
- Oliveira, H.; Melo, L.D.R.; Santos, S.B.; Nóbrega, F.L.; Ferreira, E.C.; Cerca, N.; Azeredo, J.; Kluskens, L.D. Molecular Aspects and Comparative Genomics of Bacteriophage Endolysins. J. Virol. 2013, 87, 4558–4570. [Google Scholar] [CrossRef] [PubMed]
- Yoda, T.; Matsuhashi, A.; Matsushita, A.; Shibagaki, S.; Sasakura, Y.; Aoki, K.; Hosokawa, M.; Tsuda, S. Uncovering novel endolysins against methicillin-resistant Staphylococcus aureus using microbial single-cell genome sequencing. bioRxiv 2023. [Google Scholar] [CrossRef]
- Sitthisak, S.; Manrueang, S.; Khongfak, S.; Leungtongkam, U.; Thummeepak, R.; Thanwisai, A.; Burton, N.; Dhanoa, G.K.; Tsapras, P.; Sagona, A.P. Antibacterial activity of vB_AbaM_PhT2 phage hydrophobic amino acid fusion endolysin, combined with colistin against Acinetobacter baumannii. Sci. Rep. 2023, 13, 7470. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Gajdács, M.; Baráth, Z.; Kárpáti, K.; Szabó, D.; Usai, D.; Zanetti, S.; Donadu, M.G. No Correlation between Biofilm Formation, Virulence Factors, and Antibiotic Resistance in Pseudomonas aeruginosa: Results from a Laboratory-Based In Vitro Study. Antibiotics 2021, 10, 1134. [Google Scholar] [CrossRef]
- Yang, H.; Wang, M.; Yu, J.; Wei, H. Antibacterial Activity of a Novel Peptide-Modified Lysin Against Acinetobacter baumannii and Pseudomonas aeruginosa. Front. Microbiol. 2015, 6, 1471. [Google Scholar] [CrossRef] [PubMed]
- Jung, C.-J.; Liao, Y.-D.; Hsu, C.-C.; Huang, T.-Y.; Chuang, Y.-C.; Chen, J.-W.; Kuo, Y.-M.; Chia, J.-S. Identification of potential therapeutic antimicrobial peptides against Acinetobacter baumannii in a mouse model of pneumonia. Sci. Rep. 2021, 11, 7318. [Google Scholar] [CrossRef] [PubMed]
- Peng, S.-Y.; You, R.-I.; Lai, M.-J.; Lin, N.-T.; Chen, L.-K.; Chang, K.-C. Highly potent antimicrobial modified peptides derived from the Acinetobacter baumannii phage endolysin LysAB2. Sci. Rep. 2017, 7, 11477. [Google Scholar] [CrossRef]
- Defraine, V.; Schuermans, J.; Grymonprez, B.; Govers, S.K.; Aertsen, A.; Fauvart, M.; Michiels, J.; Lavigne, R.; Briers, Y. Efficacy of Artilysin Art-175 against Resistant and Persistent Acinetobacter baumannii. Antimicrob. Agents Chemother. 2016, 60, 3480–3488. [Google Scholar] [CrossRef]
- Khan, F.M.; Gondil, V.S.; Li, C.; Jiang, M.; Li, J.; Yu, J.; Wei, H.; Yang, H. A Novel Acinetobacter baumannii Bacteriophage Endolysin LysAB54 With High Antibacterial Activity Against Multiple Gram-Negative Microbes. Front. Cell. Infect. Microbiol. 2021, 11, 637313. [Google Scholar] [CrossRef]
- Chu, D.; Lan, J.; Liang, L.; Xia, K.; Li, L.; Yang, L.; Liu, H.; Zhang, T. The antibacterial activity of a novel highly thermostable endolysin, LysKP213, against Gram-negative pathogens is enhanced when combined with outer membrane permeabilizing agents. Front. Microbiol. 2024, 15, 1454618. [Google Scholar] [CrossRef]
- Xiang, Y.; Wang, S.; Huang, H.; Li, X.; Wei, Y.; Li, H.; Ji, X. A novel endolysin from an Enterococcus faecalis phage and application. Microb. Pathog. 2024, 192, 106689. [Google Scholar] [CrossRef]
- Strange, J.E.S.; Leekitcharoenphon, P.; Møller, F.D.; Aarestrup, F.M. Metagenomics analysis of bacteriophages and antimicrobial resistance from global urban sewage. Sci. Rep. 2021, 11, 1600. [Google Scholar] [CrossRef] [PubMed]
- Howell, A.A.; Versoza, C.J.; Pfeifer, S.P. Computational host range prediction—The good, the bad, and the ugly. Virus Evol. 2024, 10, vead083. [Google Scholar] [CrossRef]
- Gerstmans, H.; Grimon, D.; Gutiérrez, D.; Lood, C.; Rodríguez, A.; van Noort, V.; Lammertyn, J.; Lavigne, R.; Briers, Y. A VersaTile-driven platform for rapid hit-to-lead development of engineered lysins. Sci. Adv. 2020, 6, eaaz1136. [Google Scholar] [CrossRef]
- Anderson, E.M.; Sychantha, D.; Brewer, D.; Clarke, A.J.; Geddes-McAlister, J.; Khursigara, C.M. Peptidoglycomics reveals compositional changes in peptidoglycan between biofilm- and planktonic-derived Pseudomonas aeruginosa. J. Biol. Chem. 2020, 295, 504–516. [Google Scholar] [CrossRef] [PubMed]
- Gantz, M.; Aleku, G.A.; Hollfelder, F. Ultrahigh-throughput screening in microfluidic droplets: A faster route to new enzymes. Trends Biochem. Sci. 2022, 47, 451–452. [Google Scholar] [CrossRef] [PubMed]
- van Kempen, M.; Kim, S.S.; Tumescheit, C.; Mirdita, M.; Lee, J.; Gilchrist, C.L.M.; Söding, J.; Steinegger, M. Fast and accurate protein structure search with Foldseek. Nat. Biotechnol. 2023, 42, 243–246. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tested Bacteria | bbgn0031 | bbgn0033 | bbgn0043 | bbgn0044 | |
| * chmrAMP | * P81417 | * Q963A8 | * Q963A8 | ||
| ** SGM003-00043_05170 | ** SGM003-00043_05170 | ** SGM003-00043_05170 | ** SGM007-00005_02751 | ||
| Gram-positive | Staphylococcus aureus | >32 | >32 | >32 | >32 |
| Streptococcus gallolyticus | >32 | >32 | >32 | >32 | |
| Bacillus subtilis | >32 | >32 | >32 | >32 | |
| Enterococcus faecalis | >32 | >32 | >32 | >32 | |
| Gram-negative | Escherichia coli | 16 | 32 | >32 | >32 |
| Acinetobacter baumannii | 8 | 4 | 8 | 8 | |
| Pseudomonas aeruginosa | 16 | 8 | 16 | >32 | |
| Enterobacter cloacae | >32 | >32 | >32 | >32 | |
| Klebsiella aerogenes | >32 | >32 | >32 | >32 | |
| Klebsiella pneumoniae | >32 | >32 | >32 | >32 | |
| Fusobacterium nucleatum | >32 | >32 | >32 | >32 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kogawa, M.; Yoda, T.; Matsuhashi, A.; Matsushita, A.; Otsuka, Y.; Shibagaki, S.; Hosokawa, M.; Tsuda, S. Development of Chimera AMP–Endolysin with Wider Spectra Against Gram-Negative Bacteria Using High-Throughput Assay. Viruses 2025, 17, 200. https://doi.org/10.3390/v17020200
Kogawa M, Yoda T, Matsuhashi A, Matsushita A, Otsuka Y, Shibagaki S, Hosokawa M, Tsuda S. Development of Chimera AMP–Endolysin with Wider Spectra Against Gram-Negative Bacteria Using High-Throughput Assay. Viruses. 2025; 17(2):200. https://doi.org/10.3390/v17020200
Chicago/Turabian StyleKogawa, Masato, Takuya Yoda, Ayumi Matsuhashi, Ai Matsushita, Yoshiki Otsuka, Shohei Shibagaki, Masahito Hosokawa, and Soichiro Tsuda. 2025. "Development of Chimera AMP–Endolysin with Wider Spectra Against Gram-Negative Bacteria Using High-Throughput Assay" Viruses 17, no. 2: 200. https://doi.org/10.3390/v17020200
APA StyleKogawa, M., Yoda, T., Matsuhashi, A., Matsushita, A., Otsuka, Y., Shibagaki, S., Hosokawa, M., & Tsuda, S. (2025). Development of Chimera AMP–Endolysin with Wider Spectra Against Gram-Negative Bacteria Using High-Throughput Assay. Viruses, 17(2), 200. https://doi.org/10.3390/v17020200