

Detection and Characterization of Influenza A Virus Endemic Circulation in Suckling and Nursery Pigs Originating from Vaccinated Farms in the Same Production System
 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design and Sample Collection
2.2. Influenza A Virus RNA Detection and Subtyping
2.3. Detection of Pandemic or North American TRIG Matrix Genes
2.4. Influenza A Virus Isolation
2.5. Influenza A Virus Complete Genome Sequencing
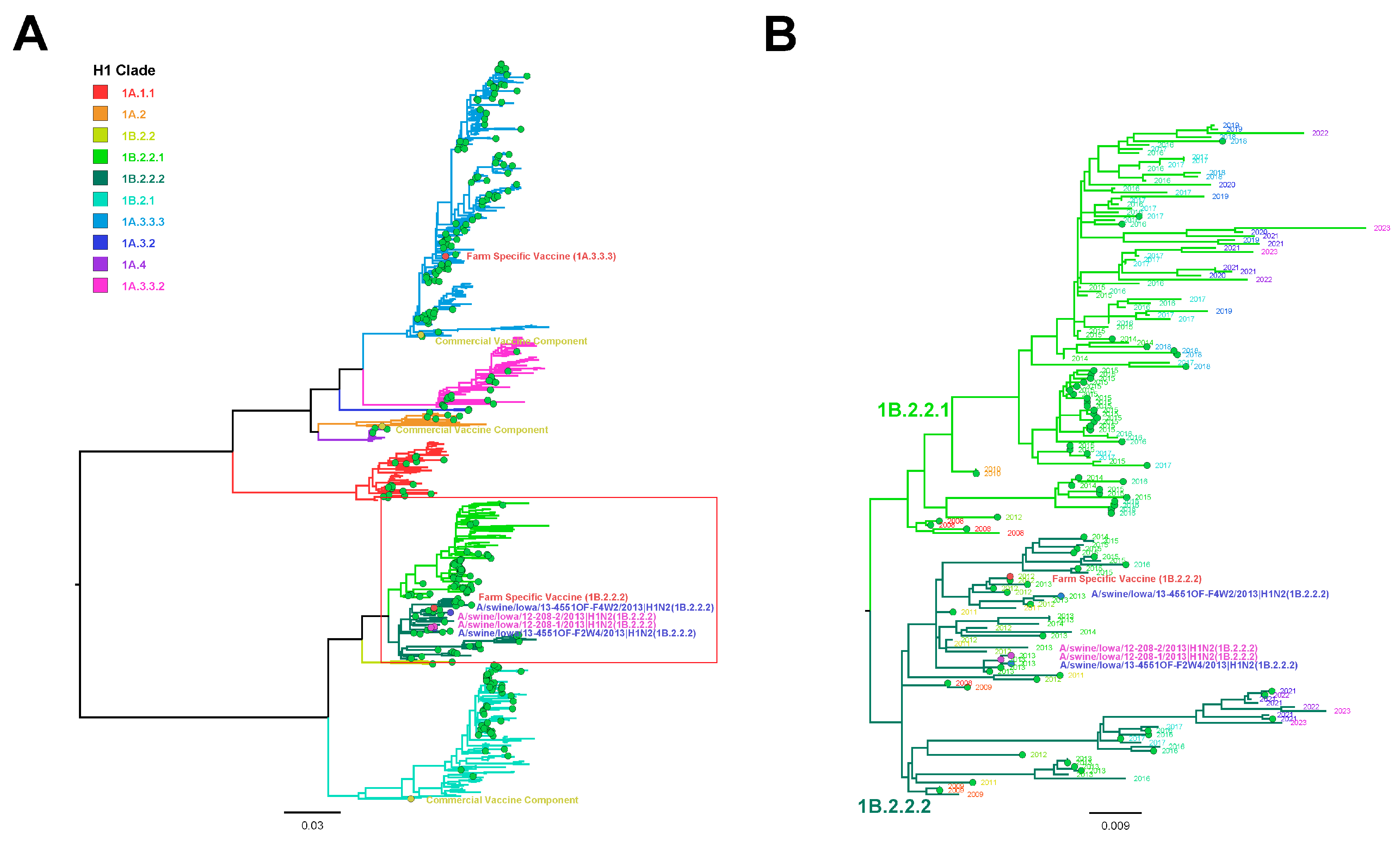
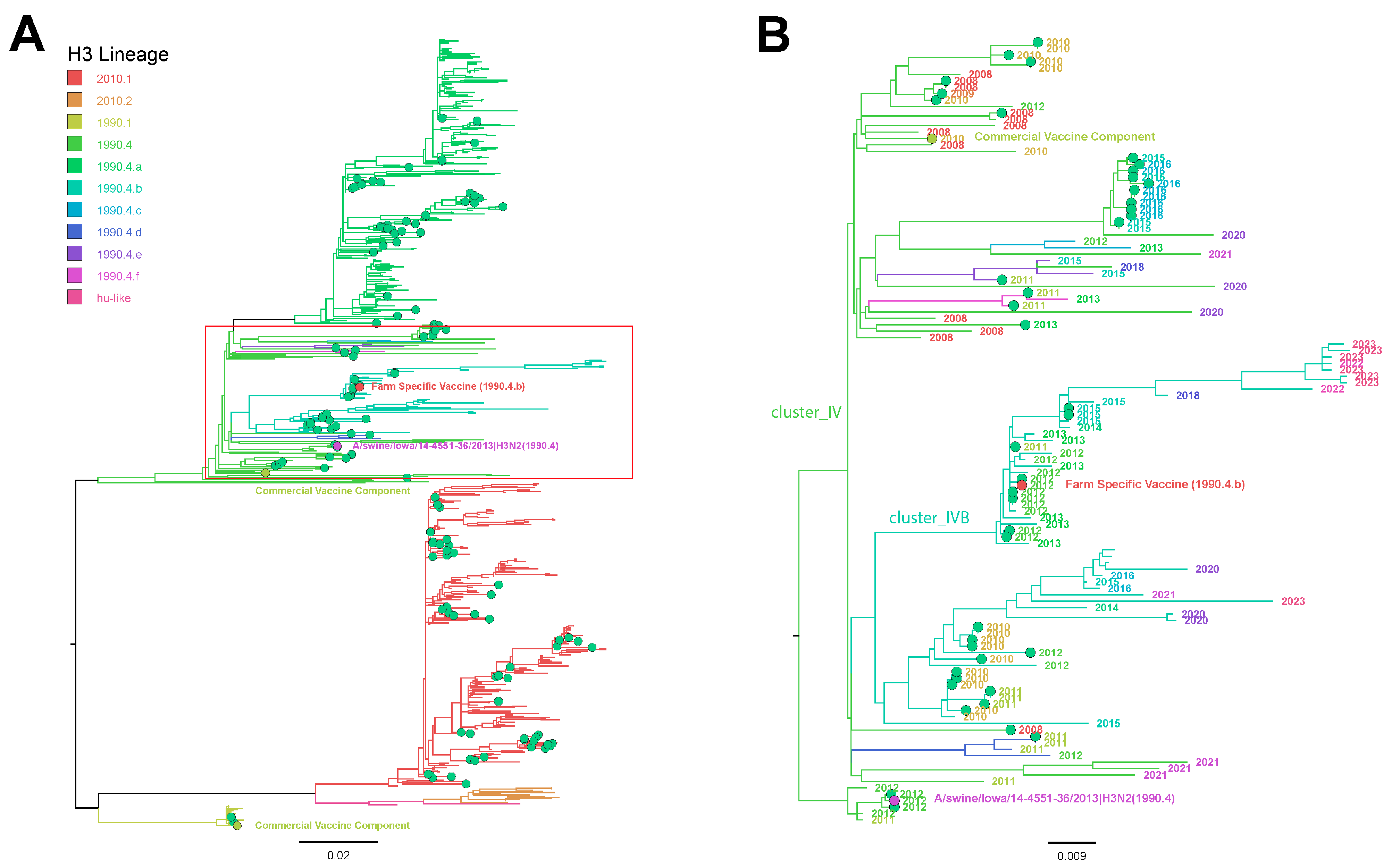
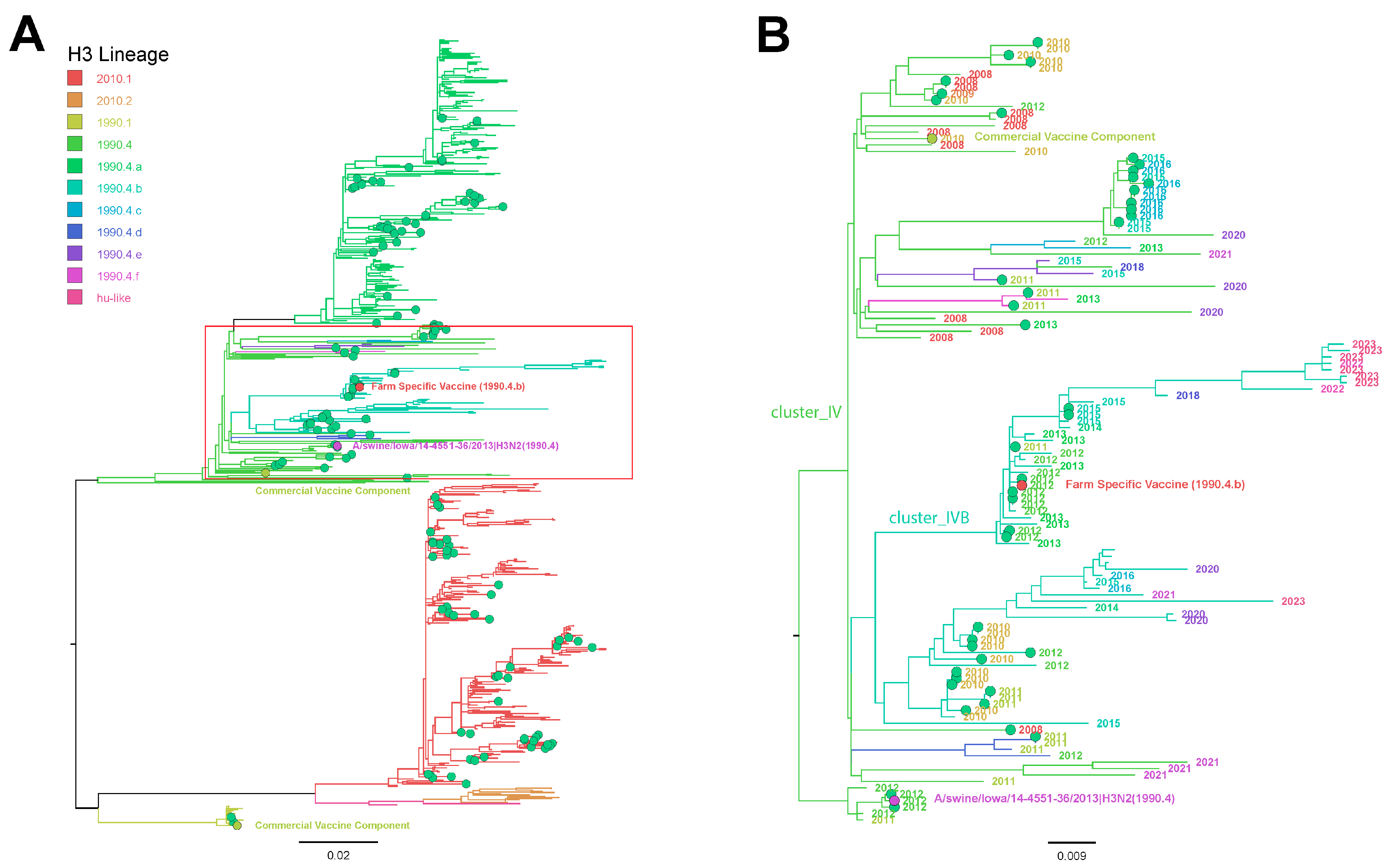
2.6. Influenza A Virus Phylogenetic Analysis
3. Results
3.1. Influenza A Virus Respiratory Clinical Signs
3.2. Detection of Influenza A Virus RNA in Suckling Pig Nasal Swabs
3.3. Detection of Influenza A Virus RNA in Nursery Pig Oral Fluids
3.4. Detection of Pandemic or North American TRIG Matrix
3.5. Influenza A Virus Isolation
3.6. Influenza A Virus Gene Sequencing
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ellis, J.; Clark, E.; Haines, D.; West, K.; Krakowka, S.; Kennedy, S.; Allan, G.M. Porcine circovirus-2 and concurrent infections in the field. Vet. Microbiol. 2004, 98, 159–163. [Google Scholar] [CrossRef] [PubMed]
- Alexander, D.J.; Brown, I.H. Recent zoonoses caused by influenza A viruses. Rev. Sci. Tech. 2000, 19, 197–225. [Google Scholar] [CrossRef]
- Richt, J.A.; Lager, K.M.; Janke, B.H.; Woods, R.D.; Webster, R.G.; Webby, R.J. Pathogenic and antigenic properties of phylogenetically distinct reassortant H3N2 swine influenza viruses cocirculating in the United States. J. Clin. Microbiol. 2003, 41, 3198–3205. [Google Scholar] [CrossRef] [PubMed]
- Vincent, A.L.; Ma, W.; Lager, K.M.; Janke, B.H.; Richt, J.A. Swine influenza viruses a North American perspective. Adv. Virus Res. 2008, 72, 127–154. [Google Scholar] [CrossRef] [PubMed]
- Zhou, N.N.; Senne, D.A.; Landgraf, J.S.; Swenson, S.L.; Erickson, G.; Rossow, K.; Liu, L.; Yoon, K.; Krauss, S.; Webster, R.G. Genetic reassortment of avian, swine, and human influenza A viruses in American pigs. J. Virol. 1999, 73, 8851–8856. [Google Scholar] [CrossRef]
- Vincent, A.L.; Ma, W.; Lager, K.M.; Gramer, M.R.; Richt, J.A.; Janke, B.H. Characterization of a newly emerged genetic cluster of H1N1 and H1N2 swine influenza virus in the United States. Virus Genes. 2009, 39, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Anderson, T.K.; Chang, J.; Arendsee, Z.W.; Venkatesh, D.; Souza, C.K.; Kimble, J.B.; Lewis, N.S.; Davis, C.T.; Vincent, A.L. Swine Influenza A Viruses and the Tangled Relationship with Humans. Cold Spring Harb. Perspect. Med. 2021, 11, a038737. [Google Scholar] [CrossRef] [PubMed]
- Kitikoon, P.; Nelson, M.I.; Killian, M.L.; Anderson, T.K.; Koster, L.; Culhane, M.R.; Vincent, A.L. Genotype patterns of contemporary reassorted H3N2 virus in US swine. J. Gen. Virol. 2013, 94, 1236–1241. [Google Scholar] [CrossRef] [PubMed]
- Rajao, D.S.; Vincent, A.L.; Perez, D.R. Adaptation of Human Influenza Viruses to Swine. Front. Vet. Sci. 2018, 5, 347. [Google Scholar] [CrossRef]
- Vincent, A.L.; Anderson, T.K.; Lager, K.M. A Brief Introduction to Influenza A Virus in Swine. Methods Mol. Biol. 2020, 2123, 249–271. [Google Scholar] [CrossRef]
- Walia, R.R.; Anderson, T.K.; Vincent, A.L. Regional patterns of genetic diversity in swine influenza A viruses in the United States from 2010 to 2016. Influenza Other Respir Viruses 2019, 13, 262–273. [Google Scholar] [CrossRef] [PubMed]
- Chamba Pardo, F.O.; Schelkopf, A.; Allerson, M.; Morrison, R.; Culhane, M.; Perez, A.; Torremorell, M. Breed-to-wean farm factors associated with influenza A virus infection in piglets at weaning. Prev. Vet. Med. 2018, 161, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Vincent, A.L.; Perez, D.R.; Rajao, D.; Anderson, T.K.; Abente, E.J.; Walia, R.R.; Lewis, N.S. Influenza A virus vaccines for swine. Vet. Microbiol. 2017, 206, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Wymore Brand, M.; Anderson, T.K.; Kitikoon, P.; Brian Kimble, J.; Otis, N.; Gauger, P.C.; Souza, C.K.; Kaplan, B.; Mogler, M.; Strait, E.; et al. Bivalent hemagglutinin and neuraminidase influenza replicon particle vaccines protect pigs against influenza a virus without causing vaccine associated enhanced respiratory disease. Vaccine 2022, 40, 5569–5578. [Google Scholar] [CrossRef] [PubMed]
- Vander Veen, R.L.; Loynachan, A.T.; Mogler, M.A.; Russell, B.J.; Harris, D.L.; Kamrud, K.I. Safety, immunogenicity, and efficacy of an alphavirus replicon-based swine influenza virus hemagglutinin vaccine. Vaccine 2012, 30, 1944–1950. [Google Scholar] [CrossRef] [PubMed]
- Vander Veen, R.L.; Harris, D.L.; Kamrud, K.I. Alphavirus replicon vaccines. Anim. Health Res. Rev. 2012, 13, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Chase, C.C. Autogenous vaccines: Current use in the field in the U.S. cattle and hog industry. Dev. Biol. 2004, 117, 69–71. [Google Scholar]
- Allerson, M.; Deen, J.; Detmer, S.E.; Gramer, M.R.; Joo, H.S.; Romagosa, A.; Torremorell, M. The impact of maternally derived immunity on influenza A virus transmission in neonatal pig populations. Vaccine 2013, 31, 500–505. [Google Scholar] [CrossRef] [PubMed]
- Kitikoon, P.; Nilubol, D.; Erickson, B.J.; Janke, B.H.; Hoover, T.C.; Sornsen, S.A.; Thacker, E.L. The immune response and maternal antibody interference to a heterologous H1N1 swine influenza virus infection following vaccination. Vet. Immunol. Immunopathol. 2006, 112, 117–128. [Google Scholar] [CrossRef]
- Loeffen, W.L.; Heinen, P.P.; Bianchi, A.T.; Hunneman, W.A.; Verheijden, J.H. Effect of maternally derived antibodies on the clinical signs and immune response in pigs after primary and secondary infection with an influenza H1N1 virus. Vet. Immunol. Immunopathol. 2003, 92, 23–35. [Google Scholar] [CrossRef]
- Rajao, D.S.; Sandbulte, M.R.; Gauger, P.C.; Kitikoon, P.; Platt, R.; Roth, J.A.; Perez, D.R.; Loving, C.L.; Vincent, A.L. Heterologous challenge in the presence of maternally-derived antibodies results in vaccine-associated enhanced respiratory disease in weaned piglets. Virology 2016, 491, 79–88. [Google Scholar] [CrossRef]
- White, L.A.; Torremorell, M.; Craft, M.E. Influenza A virus in swine breeding herds: Combination of vaccination and biosecurity practices can reduce likelihood of endemic piglet reservoir. Prev. Vet. Med. 2017, 138, 55–69. [Google Scholar] [CrossRef] [PubMed]
- Corzo, C.A.; Culhane, M.; Dee, S.; Morrison, R.B.; Torremorell, M. Airborne detection and quantification of swine influenza a virus in air samples collected inside, outside and downwind from swine barns. PLoS ONE 2013, 8, e71444. [Google Scholar] [CrossRef]
- Diaz, A.; Perez, A.; Sreevatsan, S.; Davies, P.; Culhane, M.; Torremorell, M. Association between Influenza A Virus Infection and Pigs Subpopulations in Endemically Infected Breeding Herds. PLoS ONE 2015, 10, e0129213. [Google Scholar] [CrossRef]
- Allerson, M.W.; Davies, P.R.; Gramer, M.R.; Torremorell, M. Infection dynamics of pandemic 2009 H1N1 influenza virus in a two-site swine herd. Transbound. Emerg. Dis. 2014, 61, 490–499. [Google Scholar] [CrossRef] [PubMed]
- Diaz, A.; Marthaler, D.; Culhane, M.; Sreevatsan, S.; Alkhamis, M.; Torremorell, M. Complete Genome Sequencing of Influenza A Viruses within Swine Farrow-to-Wean Farms Reveals the Emergence, Persistence, and Subsidence of Diverse Viral Genotypes. J. Virol. 2017, 91, e00745-17. [Google Scholar] [CrossRef]
- Nelson, M.I.; Viboud, C.; Vincent, A.L.; Culhane, M.R.; Detmer, S.E.; Wentworth, D.E.; Rambaut, A.; Suchard, M.A.; Holmes, E.C.; Lemey, P. Global migration of influenza A viruses in swine. Nat. Commun. 2015, 6, 6696. [Google Scholar] [CrossRef] [PubMed]
- Brown, I.H. The epidemiology and evolution of influenza viruses in pigs. Vet. Microbiol. 2000, 74, 29–46. [Google Scholar] [CrossRef]
- Chamba Pardo, F.O.; Wayne, S.; Culhane, M.R.; Perez, A.; Allerson, M.; Torremorell, M. Effect of strain-specific maternally-derived antibodies on influenza A virus infection dynamics in nursery pigs. PLoS ONE 2019, 14, e0210700. [Google Scholar] [CrossRef]
- CPardo, F.O.C.; Allerson, M.W.; Culhane, M.R.; Morrison, R.B.; Davies, P.R.; Perez, A.; Torremorell, M. Effect of influenza A virus sow vaccination on infection in pigs at weaning: A prospective longitudinal study. Transbound. Emerg. Dis. 2021, 68, 183–193. [Google Scholar] [CrossRef]
- Prickett, J.R.; Zimmerman, J.J. The development of oral fluid-based diagnostics and applications in veterinary medicine. Anim. Health Res. Rev. 2010, 11, 207–216. [Google Scholar] [CrossRef]
- Harmon, K.; Bower, L.; Kim, W.I.; Pentella, M.; Yoon, K.J. A matrix gene-based multiplex real-time RT-PCR for detection and differentiation of 2009 pandemic H1N1 and other influenza A viruses in North America. Influenza Other Respir Viruses 2010, 4, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Gauger, P.C. Isolation of swine influenza virus in cell cultures and embryonated chicken eggs. Methods Mol. Biol. 2014, 1161, 265–276. [Google Scholar] [CrossRef]
- Bowman, A.S.; Sreevatsan, S.; Killian, M.L.; Page, S.L.; Nelson, S.W.; Nolting, J.M.; Cardona, C.; Slemons, R.D. Molecular evidence for interspecies transmission of H3N2pM/H3N2v influenza A viruses at an Ohio agricultural fair, July 2012. Emerg. Microbes Infect. 2012, 1, e33. [Google Scholar] [CrossRef]
- Rothberg, J.M.; Hinz, W.; Rearick, T.M.; Schultz, J.; Mileski, W.; Davey, M.; Leamon, J.H.; Johnson, K.; Milgrew, M.J.; Edwards, M.; et al. An integrated semiconductor device enabling non-optical genome sequencing. Nature 2011, 475, 348–352. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Anderson, T.K.; Nelson, M.I.; Kitikoon, P.; Swenson, S.L.; Korslund, J.A.; Vincent, A.L. Population dynamics of cocirculating swine influenza A viruses in the United States from 2009 to 2012. Influenza Other Respir Viruses 2013, 7 (Suppl. S4), 42–51. [Google Scholar] [CrossRef]
- Beaudoin, A.; Johnson, S.; Davies, P.; Bender, J.; Gramer, M. Characterization of influenza a outbreaks in Minnesota swine herds and measures taken to reduce the risk of zoonotic transmission. Zoonoses Public. Health 2012, 59, 96–106. [Google Scholar] [CrossRef]
- Lopez-Moreno, G.; Culhane, M.R.; Davies, P.; Corzo, C.; Allerson, M.W.; Torremorell, M. Farm management practices associated with influenza A virus contamination of people working in Midwestern United States swine farms. Porc. Health Manag. 2023, 9, 13. [Google Scholar] [CrossRef]
- Pitzer, V.E.; Aguas, R.; Riley, S.; Loeffen, W.L.; Wood, J.L.; Grenfell, B.T. High turnover drives prolonged persistence of influenza in managed pig herds. J. R. Soc. Interface 2016, 13, 20160138. [Google Scholar] [CrossRef]
- Reynolds, J.J.; Torremorell, M.; Craft, M.E. Mathematical modeling of influenza A virus dynamics within swine farms and the effects of vaccination. PLoS ONE 2014, 9, e106177. [Google Scholar] [CrossRef]
- Simon-Grifé, M.; Martín-Valls, G.E.; Vilar, M.J.; Busquets, N.; Mora-Salvatierra, M.; Bestebroer, T.M.; Fouchier, R.A.; Martín, M.; Mateu, E.; Casal, J. Swine influenza virus infection dynamics in two pig farms; results of a longitudinal assessment. Vet. Res. 2012, 43, 24. [Google Scholar] [CrossRef] [PubMed]
- Almeida, M.N.; Zhang, M.; Zimmerman, J.J.; Holtkamp, D.J.; Linhares, D.C.L. Finding PRRSV in sow herds: Family oral fluids vs. serum samples from due-to-wean pigs. Prev. Vet. Med. 2021, 193, 105397. [Google Scholar] [CrossRef]
- Garrido-Mantilla, J.; Alvarez, J.; Culhane, M.; Nirmala, J.; Cano, J.P.; Torremorell, M. Comparison of individual, group and environmental sampling strategies to conduct influenza surveillance in pigs. BMC Vet. Res. 2019, 15, 61. [Google Scholar] [CrossRef]
- Rose, N.; Herve, S.; Eveno, E.; Barbier, N.; Eono, F.; Dorenlor, V.; Andraud, M.; Camsusou, C.; Madec, F.; Simon, G. Dynamics of influenza A virus infections in permanently infected pig farms: Evidence of recurrent infections, circulation of several swine influenza viruses and reassortment events. Vet. Res. 2013, 44, 72. [Google Scholar] [CrossRef]
- Sandbulte, M.R.; Spickler, A.R.; Zaabel, P.K.; Roth, J.A. Optimal Use of Vaccines for Control of Influenza A Virus in Swine. Vaccines 2015, 3, 22–73. [Google Scholar] [CrossRef] [PubMed]
- Meiners, C.; Loesken, S.; Doehring, S.; Starick, E.; Pesch, S.; Maas, A.; Noe, T.; Beer, M.; Harder, T.; Grosse Beilage, E. Field study on swine influenza virus (SIV) infection in weaner pigs and sows. Tierarztl. Prax. Ausg. G. Grosstiere Nutztiere 2014, 42, 351–359. [Google Scholar] [CrossRef]
- Janke, B.H. Clinicopathological features of Swine influenza. Curr. Top. Microbiol. Immunol. 2013, 370, 69–83. [Google Scholar] [CrossRef]
- Prickett, J.R.; Kim, W.; Simer, R.; Yoon, K.J.; Zimmerman, J. Oral-fluid samples for surveillance of commercial growing pigs for porcine reproductive and respiratory syndrome virus and porcine circovirus type 2 infections. J. Swine Health Prod. 2008, 16, 6. [Google Scholar]
- Wills, R.W.; Zimmerman, J.J.; Yoon, K.J.; Swenson, S.L.; Hoffman, L.J.; McGinley, M.J.; Hill, H.T.; Platt, K.B. Porcine reproductive and respiratory syndrome virus: Routes of excretion. Vet. Microbiol. 1997, 57, 69–81. [Google Scholar] [CrossRef]
- Detmer, S.E.; Patnayak, D.P.; Jiang, Y.; Gramer, M.R.; Goyal, S.M. Detection of Influenza A virus in porcine oral fluid samples. J. Vet. Diagn. Invest. 2011, 23, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Vander Veen, R.; Kamrud, K.; Mogler, M.; Loynachan, A.T.; McVicker, J.; Berglund, P.; Owens, G.; Timberlake, S.; Lewis, W.; Smith, J.; et al. Rapid development of an efficacious swine vaccine for novel H1N1. PLoS Curr. 2009, 1, Rrn1123. [Google Scholar] [CrossRef]
- Vander Veen, R.L.; Mogler, M.A.; Russell, B.J.; Loynachan, A.T.; Harris, D.L.; Kamrud, K.I. Haemagglutinin and nucleoprotein replicon particle vaccination of swine protects against the pandemic H1N1 2009 virus. Vet. Rec. 2013, 173, 344. [Google Scholar] [CrossRef] [PubMed]
- Ducatez, M.F.; Hause, B.; Stigger-Rosser, E.; Darnell, D.; Corzo, C.; Juleen, K.; Simonson, R.; Brockwell-Staats, C.; Rubrum, A.; Wang, D.; et al. Multiple reassortment between pandemic (H1N1) 2009 and endemic influenza viruses in pigs, United States. Emerg. Infect. Dis. 2011, 17, 1624–1629. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Anderson, T.K.; Walia, R.R.; Dorman, K.S.; Janas-Martindale, A.; Vincent, A.L. The genomic evolution of H1 influenza A viruses from swine detected in the United States between 2009 and 2016. J Gen Virol 2017, 98, 2001–2010. [Google Scholar] [CrossRef]
- Nelson, M.I.; Gramer, M.R.; Vincent, A.L.; Holmes, E.C. Global transmission of influenza viruses from humans to swine. J. Gen. Virol. 2012, 93, 2195–2203. [Google Scholar] [CrossRef]
- Nelson, M.I.; Vincent, A.L. Reverse zoonosis of influenza to swine: New perspectives on the human-animal interface. Trends Microbiol. 2015, 23, 142–153. [Google Scholar] [CrossRef] [PubMed]
- Epperson, S.; Jhung, M.; Richards, S.; Quinlisk, P.; Ball, L.; Moll, M.; Boulton, R.; Haddy, L.; Biggerstaff, M.; Brammer, L.; et al. Human infections with influenza A(H3N2) variant virus in the United States, 2011–2012. Clin. Infect. Dis. 2013, 57 (Suppl. S1), S4–S11. [Google Scholar] [CrossRef]
- Schicker, R.S.; Rossow, J.; Eckel, S.; Fisher, N.; Bidol, S.; Tatham, L.; Matthews-Greer, J.; Sohner, K.; Bowman, A.S.; Avrill, J.; et al. Outbreak of Influenza A(H3N2) Variant Virus Infections Among Persons Attending Agricultural Fairs Housing Infected Swine—Michigan and Ohio, July–August 2016. Morb. Mortal. Wkly. Rep. 2016, 65, 1157–1160. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Sample Collection | ||||||||
|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | |
| Collection Date | 11–15 March | 25–29 March | 8–12 April | 22–26 April | 6–9 May | 8–12 July | 22–26 July | 5–9 August |
| Vaccine Dose | Prime dose | Prime dose | Boost dose | Boost dose | ||||
| Administration Date | March 4 | March 25 | April 1 | April 15 | ||||
| Farms Vaccinated | F1, F2, F3 | F4 * | F1, F3 | F2 | ||||
| Group | Location | Age (Days) | Sample Type | Samples/Farm | # of Farms | # of Samplings | Total Samples |
|---|---|---|---|---|---|---|---|
| Suckling Pigs | Farrowing | 12–17 | Nasal Swabs | 135 | 4 | 8 | 4320 |
| Nursery Pigs | Nursery | 28–35 | Oral Fluids | 3–8 | 4 | 8 | 158 |
| Farm 1 | Farm 2 | Farm 3 | Farm 4 | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| H1 | H3 | H1/H3 | H1 | H3 | H1/H3 | H1 | H3 | H1/H3 | H1 | H3 | H1/H3 | |||||||||
| Time | #/# Pos. (%) | Mean Ct [SEM] | #/# Pos. (%) | Mean Ct [SEM] | #/# Pos. (%) | #/# Pos. (%) | Mean Ct [SEM] | #/# Pos. (%) | Mean Ct [SEM] | #/# Pos. (%) | #/# Pos. (%) | Mean Ct [SEM] | #/# Pos. (%) | Mean Ct [SEM] | #/# Pos. (%) | #/# Pos. (%) | Mean Ct [SEM] | #/# Pos. (%) | Mean Ct [SEM] | #/# Pos. (%) |
| 1 | - | - | - | - | - | 38/135 (28.1) 5 * | 29.6 [0.6] | 3/135 (2.2) 29 * | 34.4 [0.4] | 11/135 (8.1) 2 * | 1/135 (0.7) 1 * | 31.7 NA | - | - | - | 2/135 (1.5) | 33.8 [1.1] | - | - | - |
| 2 | 6/135 (4.4) | 32.6 [0.6] | - | - | - | 8/135 (6.0) 6 * | 33.3 [0.6] | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 3 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 4 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 5 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 6 | 2/135 (1.5) 1 * | 33.3 [0.1] | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 * | NA | - |
| 7 | 2 * | NA | 1 * | NA | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 8 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Pos/ Farm | 8/1080 (0.7) | 32.8 [0.4] | - | - | - | 46/1080 (4.2) | 30.2 [0.5] | 3/1080 (0.3) | 34.4 [0.8] | 11/1080 (1.0) | 1/1080 (0.1) | 31.7 NA | - | - | - | 2/1080 (0.2) | 33.8 [1.1] | - | - | - |
| Farm 1 | Farm 2 | Farm 3 | Farm 4 | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N1 | N2 | N1/N2 | N1 | N2 | N1/N2 | N1 | N2 | N1/N2 | N1 | N2 | N1/N2 | |||||||||
| Time | #/# Pos. (%) | Mean Ct [SEM] | #/# Pos. (%) | Mean Ct [SEM] | #/# Pos. (%) | #/# Pos. (%) | Mean Ct [SEM] | #/# Pos. (%) | Mean Ct [SEM] | #/# Pos. (%) | #/# Pos. (%) | Mean Ct [SEM] | #/# Pos. (%) | Mean Ct [SEM] | #/# Pos. (%) | #/# Pos. (%) | Mean Ct [SEM] | #/# Pos. (%) | Mean Ct [SEM] | #/# Pos. (%) |
| 1 | - | - | - | - | - | 15 * | NA | 54/135 (28.1) | 29.7 [0.5] | - | - | - | 2/135 (1.5) | 32.8 [1.9] | - | - | - | 2/135 (1.5) | 33.4 [0.9] | - |
| 2 | - | - | 6/135 (4.4) | 32.0 [0.5] | - | 1 * | NA | 10/135 (7.4) 6 * | 32.9 [0.4] | - | - | - | - | - | - | - | - | - | - | - |
| 3 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 4 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 5 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 6 | - | - | 2/135 (1.5) 1 * | 32.9 [0.5] | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 7 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 8 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Pos/ Farm | - | - | 8/1080 (0.7) | 30.2 [0.4] | - | - | - | 64/1080 (5.9) | 30.2 [0.4] | - | - | - | 2/1080 (0.2) | 32.8 [1.9] | - | - | - | 2/1080 (0.2) | 33.4 [0.9] | - |
| Farm 1 | Farm 2 | Farm 3 | Farm 4 | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| H1 | H3 | H1/H3 | H1 | H3 | H1/H3 | H1 | H3 | H1/H3 | H1 | H3 | H1/H3 | |||||||||
| Time | #/# Pos. (%) | Mean Ct (SEM) | #/# Pos. (%) | Mean Ct (SEM) | #/# Pos. (%) | #/# Pos. (%) | Mean Ct (SEM) | #/# Pos. (%) | Mean Ct (SEM) | #/# Pos. (%) | #/# Pos. (%) | Mean Ct (SEM) | #/# Pos. (%) | Mean Ct (SEM) | #/# Pos. (%) | #/# Pos. (%) | Mean Ct (SEM) | #/# Pos. (%) | Mean Ct (SEM) | #/# Pos. (%) |
| 1 | - | - | 1 * | NA | - | 4/8 (50.0) 1 * | 34.0 [0.3] | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 2 | - | - | - | - | - | 1/8 (12.5) 3 * | 33.5 [0.0] | - | - | - | 1 * | NA | - | - | 1 * | 1/8 (12.5) 1 * | 31.9 [0.0] | - | - | - |
| 3 | 1 * | NA | - | - | - | 2/7 (28.6) 2 * | 32.5 [0.3] | - | - | - | - | - | - | - | 1 * | - | - | 3/49 (6.1) | 29.6 [1.2] | - |
| 4 | - | - | - | - | - | 6/8 (75.0) 1 * | 32.2 [0.8] | - | - | - | NT | NT | NT | NT | NT | - | - | - | - | 1 * |
| 5 | - | - | - | - | - | NT | NT | NT | NT | NT | NT | NT | NT | NT | NT | NT | NT | NT | NT | NT |
| 6 | 1 * | NA | 1 * | NA | - | - | - | - | - | - | - | - | - | - | - | 1 * | NA | - | - | - |
| 7 | NT | NT | NT | NT | NT | - | - | - | - | - | NT | NT | NT | NT | NT | - | - | 1 * | NA | - |
| 8 | NT | NT | NT | NT | NT | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Farm Total | - | - | - | - | - | 13/49 (26.5) | 32.9 [0.4] | - | - | - | - | - | - | - | - | 1/49 (2.0) | 31.9 [0.0] | 3/49 (6.1) | 30.1 [1.8] | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dias, A.S.; Baker, A.L.V.; Baker, R.B.; Zhang, J.; Zeller, M.A.; Kitikoon, P.; Gauger, P.C. Detection and Characterization of Influenza A Virus Endemic Circulation in Suckling and Nursery Pigs Originating from Vaccinated Farms in the Same Production System. Viruses 2024, 16, 626. https://doi.org/10.3390/v16040626
Dias AS, Baker ALV, Baker RB, Zhang J, Zeller MA, Kitikoon P, Gauger PC. Detection and Characterization of Influenza A Virus Endemic Circulation in Suckling and Nursery Pigs Originating from Vaccinated Farms in the Same Production System. Viruses. 2024; 16(4):626. https://doi.org/10.3390/v16040626
Chicago/Turabian StyleDias, Alessandra Silva, Amy L. Vincent Baker, Rodney B. Baker, Jianqiang Zhang, Michael A. Zeller, Pravina Kitikoon, and Phillip C. Gauger. 2024. "Detection and Characterization of Influenza A Virus Endemic Circulation in Suckling and Nursery Pigs Originating from Vaccinated Farms in the Same Production System" Viruses 16, no. 4: 626. https://doi.org/10.3390/v16040626
APA StyleDias, A. S., Baker, A. L. V., Baker, R. B., Zhang, J., Zeller, M. A., Kitikoon, P., & Gauger, P. C. (2024). Detection and Characterization of Influenza A Virus Endemic Circulation in Suckling and Nursery Pigs Originating from Vaccinated Farms in the Same Production System. Viruses, 16(4), 626. https://doi.org/10.3390/v16040626