
Identification of Mycoviruses in the Pathogens of Fragrant Pear Valsa Canker from Xinjiang in China

Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation and Identification of Pathogens
2.2. DNA and RNA Extraction
2.3. Metatranscriptomic Sequencing and Bioinformatics Analysis
2.4. Putative Mycovirus Sequence Confirmation
2.5. Phylogenetic Analysis
2.6. Growth Rate and Virulence Assay
2.7. Horizontal Transmission of Hypovirulence Traits
3. Results
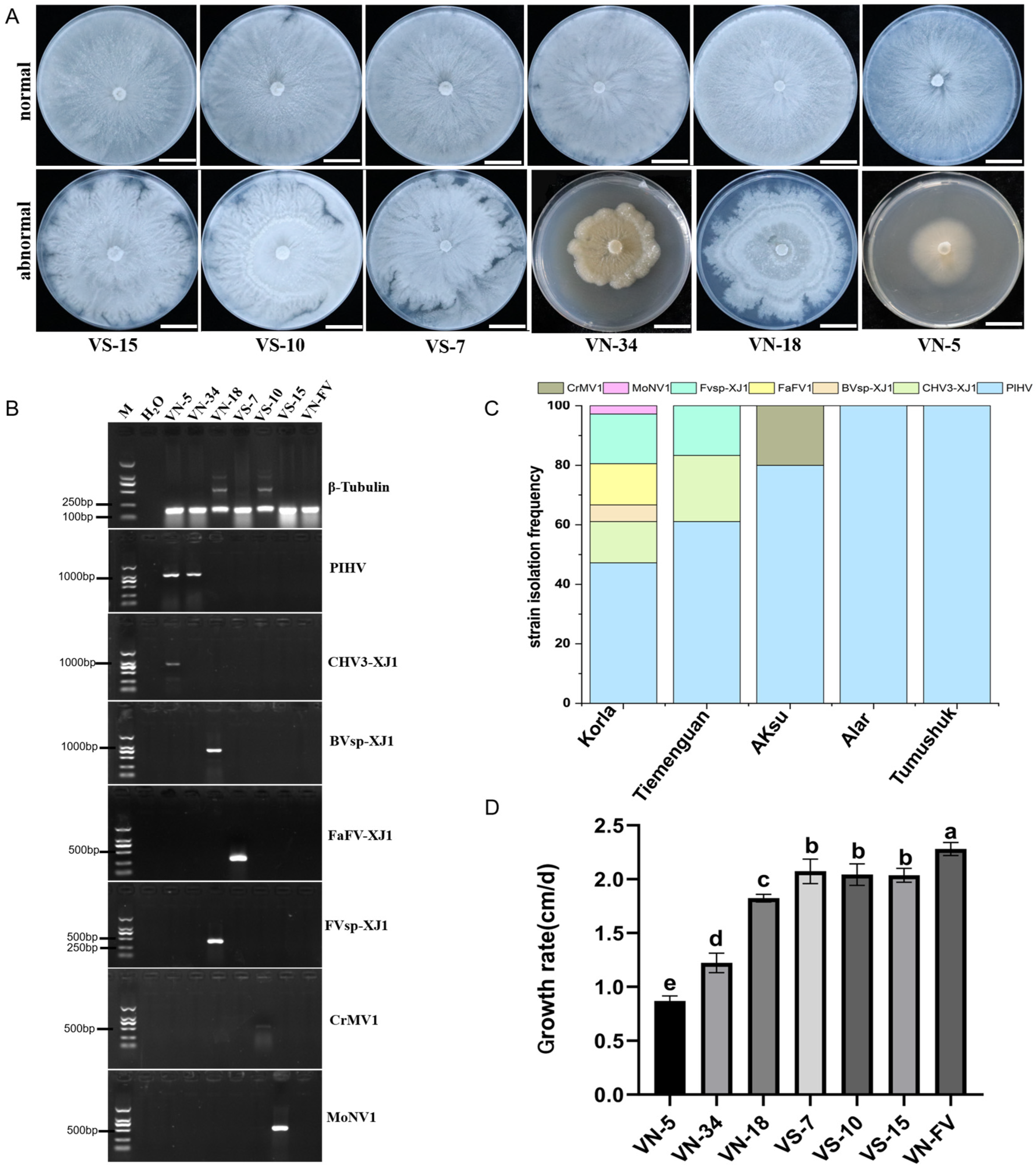
3.1. Isolation and Identification of Fragrant Pear Canker Disease Pathogens
3.2. Metatranscriptomic Identification of Mycoviruses Infecting the Tested Strains
3.3. Mycovirus Detection and Its Effect on the Growth of Valsa
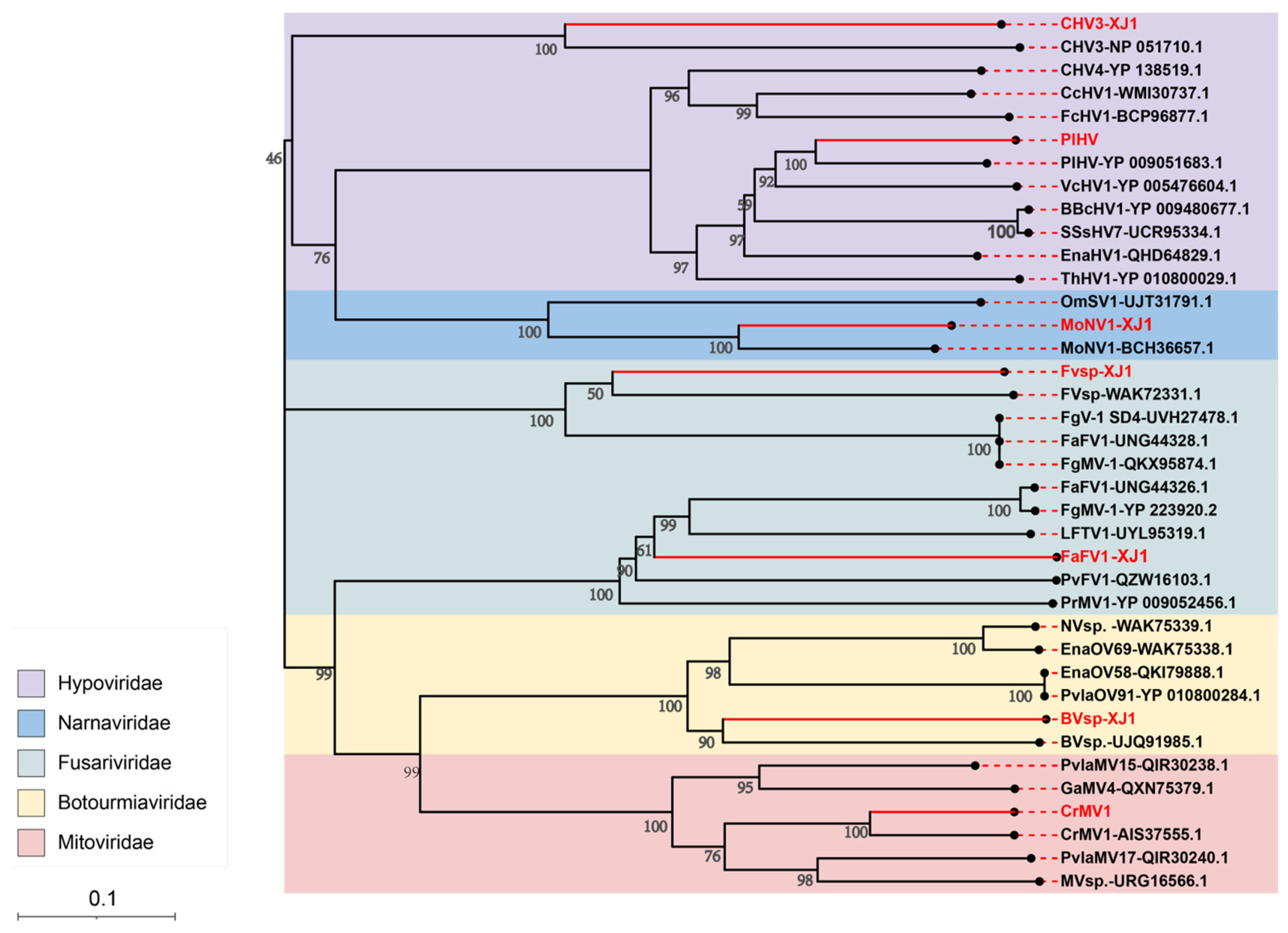
3.4. Viral Sequence Phylogenetic Tree Analysis
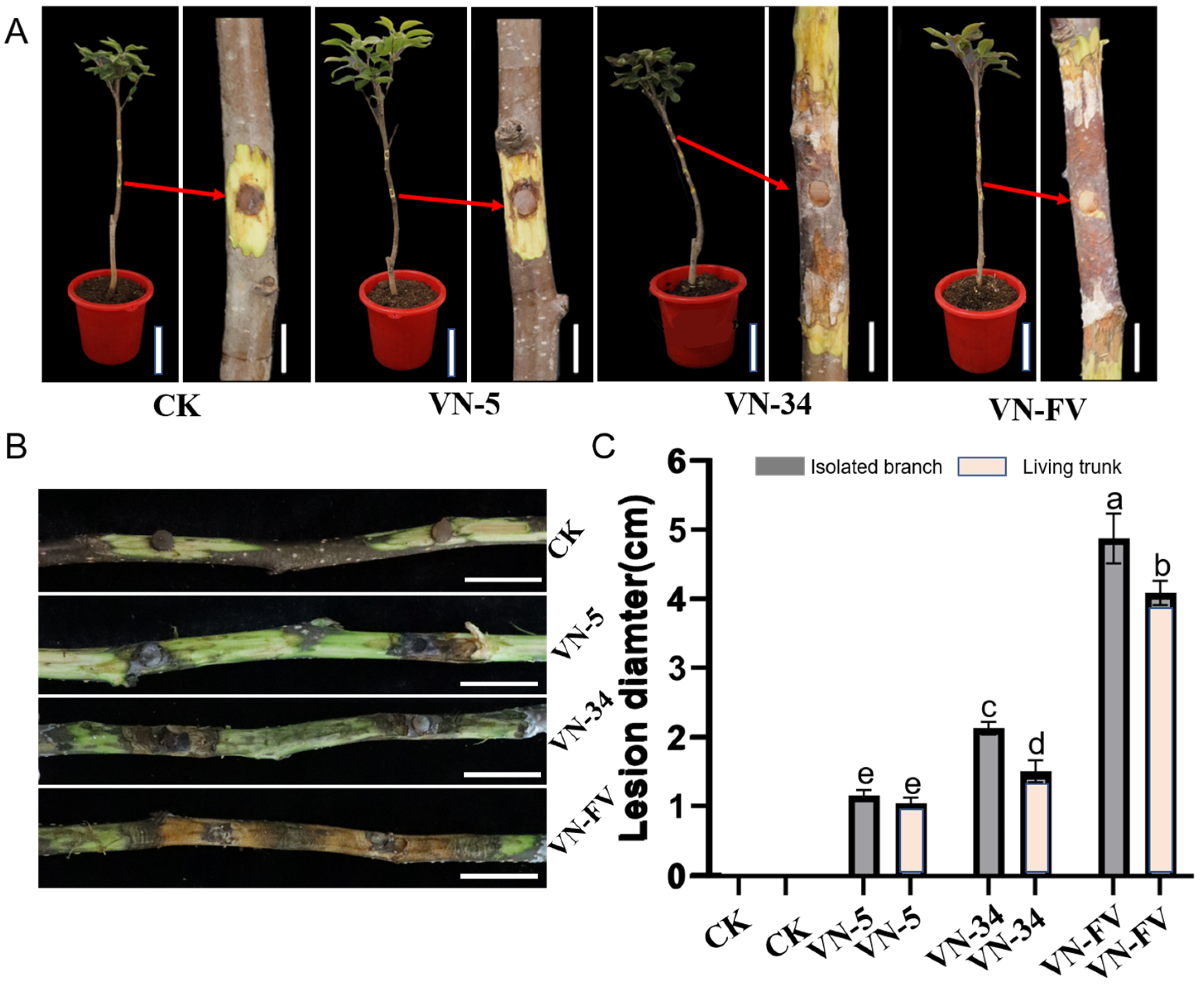
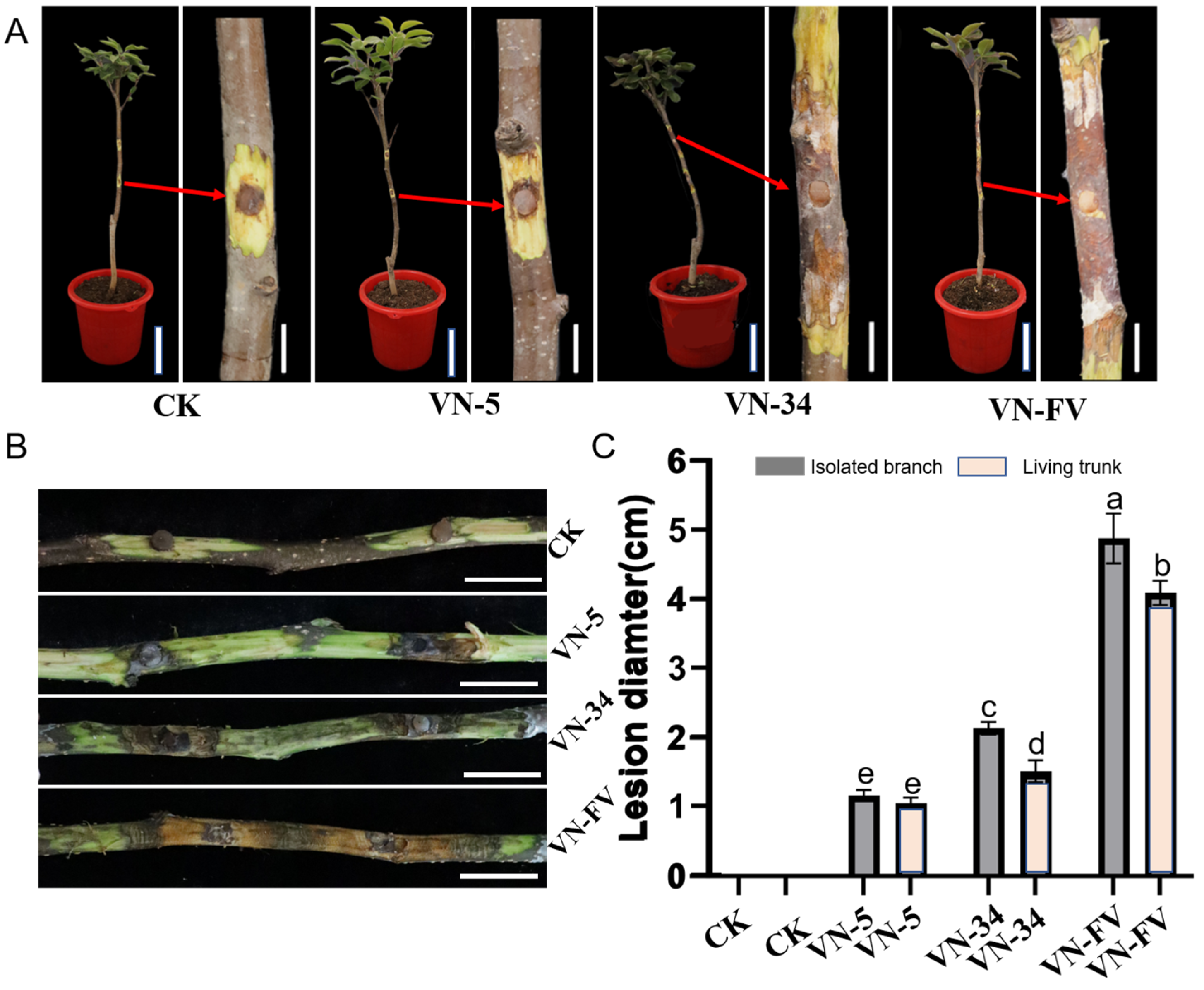
3.5. Hypovirulence Associated with PlHV and CHV3-XJ1
3.6. Horizontal Transmission of PlHV and CHV3-XJ1
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Niu, Y.; Chen, X.; Zhou, W.; Li, W.; Zhao, S.; Nasir, M.; Dong, S.; Zhang, S.; Liao, K. Genetic relationship between the ‘Korla fragrant pear’ and local pear varieties in Xinjiang based on floral organ characteristics. Sci. Hortic. 2019, 257, 108621. [Google Scholar] [CrossRef]
- Li, Z.; Yin, Z.; Fan, Y.; Xu, M.; Kang, Z.; Huang, L. Candidate effector proteins of the necrotrophic apple canker pathogen Valsa mali can suppress BAX-induced PCD. Front. Plant Sci. 2015, 6, 579. [Google Scholar] [CrossRef] [PubMed]
- Yin, Z.; Liu, H.; Li, Z.; Ke, X.; Dou, D.; Gao, X.; Song, N.; Dai, Q.; Wu, Y.; Xu, J.; et al. Genome sequence of Valsa canker pathogens uncovers a potential adaptation of colonization of woody bark. New Phytol. 2015, 208, 1202–1216. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Wang, W.; Li, G.; Du, M.; Bi, S.; Zhao, M.; Cao, G.; Li, H. Selection of field control agents and comprehensive control effect of pear Valsa canker. J. Fruit Sci. 2018, 35, 143–147. [Google Scholar] [CrossRef]
- Xu, L.; Meng, Y.; Liu, R.; Xiao, Y.; Wang, Y.; Huang, L. Inhibitory effects of Bacillus vallismortis T27 against apple Valsa canker caused by Valsa mali. Pestic. Biochem. Physiol. 2023, 195, 105564. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Shi, B.; Wang, Z.; Qin, G.; Hou, H.; Tu, H.; Wang, L. Exploration of the biocontrol activity of Bacillus atrophaeus strain HF1 against pear Valsa canker caused by Valsa pyri. Int. J. Mol. Sci. 2023, 24, 15477. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Jiang, D. New insights into mycoviruses and exploration for the biological control of crop fungal diseases. Annu. Rev. Phytopathol. 2014, 52, 45–68. [Google Scholar] [CrossRef]
- Ghabrial, S.A.; Castón, J.R.; Jiang, D.; Nibert, M.L.; Suzuki, N. 50-plus years of fungal viruses. Virology 2015, 479–480, 356–368. [Google Scholar] [CrossRef]
- Jiang, D.; Fu, Y.; Li, G.; Ghabrial, S.A. Viruses of the plant pathogenic fungus Sclerotinia sclerotiorum. Adv. Virus Res. 2013, 86, 215–248. [Google Scholar] [CrossRef]
- Rodríguez-García, C.; Medina, V.; Alonso, A.; Ayllón, M.A. Mycoviruses of Botrytis cinerea isolates from different hosts: Mycovirus and plant pathogenic fungi. Ann. Appl. Biol. 2014, 164, 46–61. [Google Scholar] [CrossRef]
- Zhong, J.; Chen, D.; Lei, X.; Zhu, H.; Zhu, J.; Da Gao, B. Detection and characterization of a novel gammapartitivirus in the phytopathogenic fungus Colletotrichum acutatum strain HNZJ001. Virus Res. 2014, 190, 104–109. [Google Scholar] [CrossRef]
- Ai, Y.; Zhong, J.; Chen, C.; Zhu, H.; Gao, B. A novel single-stranded RNA virus isolated from the rice-pathogenic fungus Magnaporthe oryzae with similarity to members of the family tombusviridae. Arch. Virol. 2016, 161, 725–729. [Google Scholar] [CrossRef]
- Kondo, H.; Hisano, S.; Chiba, S.; Maruyama, K.; Andika, I.B.; Toyoda, K.; Fujimori, F.; Suzuki, N. Sequence and phylogenetic analyses of novel totivirus-like double-stranded RNAs from field-collected powdery mildew fungi. Virus Res. 2016, 213, 353–364. [Google Scholar] [CrossRef]
- Gupta, T.; Kumari, C.; Vanshika; Kulshrestha, S. Biology and mycovirus-assisted biological control of Sclerotinia sclerotiorum infecting vegetable and oilseed crops. Arch. Phytopathol. Pflanzenschutz. 2019, 52, 1049–1067. [Google Scholar] [CrossRef]
- Zhu, H.; Chen, D.; Zhong, J.; Zhang, S.; Gao, B. A novel mycovirus identified from the rice false smut fungus Ustilaginoidea virens. Virus Genes 2015, 51, 159–162. [Google Scholar] [CrossRef] [PubMed]
- Pearson, M.N.; Beever, R.E.; Boine, B.; Arthur, K. Mycoviruses of filamentous fungi and their relevance to plant pathology. Mol. Plant Pathol. 2009, 10, 115–128. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, Q.; Iritani, A.; Ohkita, S.; Vu, B.V.; Yokoya, K.; Matsubara, A.; Ikeda, K.-I.; Suzuki, N.; Nakayashiki, H. A fungal argonaute interferes with RNA interference. Nucleic Acids Res. 2018, 46, 2495–2508. [Google Scholar] [CrossRef] [PubMed]
- Lau, S.K.P.; Lo, G.C.S.; Chow, F.W.N.; Fan, R.Y.Y.; Cai, J.J.; Yuen, K.Y.; Woo, P.C.Y. Novel partitivirus enhances virulence of and causes aberrant gene expression in Talaromyces marneffei. mBio 2018, 9, e00947-18. [Google Scholar] [CrossRef] [PubMed]
- Meyer, J.B.; Chalmandrier, L.; Fässler, F.; Schefer, C.; Rigling, D.; Prospero, S. Role of fresh dead wood in the epidemiology and the biological control of the chestnut blight fungus. Plant Dis. 2019, 103, 430–438. [Google Scholar] [CrossRef] [PubMed]
- Chiba, S.; Salaipeth, L.; Lin, Y.; Sasaki, A.; Kanematsu, S.; Suzuki, N. A novel bipartite double-stranded RNA mycovirus from the white root rot fungus Rosellinia necatrix: Molecular and biological characterization, taxonomic considerations, and potential for biological control. J. Virol. 2009, 83, 12801–12812. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Li, B.; Fu, Y.; Jiang, D.; Ghabrial, S.A.; Li, G.; Peng, Y.; Xie, J.; Cheng, J.; Huang, J.; et al. A geminivirus-related DNA mycovirus that confers hypovirulence to a plant pathogenic fungus. Proc. Natl. Acad. Sci. USA 2010, 107, 8387–8392. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Li, B.; Fu, Y.; Xie, J.; Cheng, J.; Ghabrial, S.A.; Li, G.; Yi, X.; Jiang, D. Extracellular transmission of a DNA mycovirus and its use as a natural fungicide. Proc. Natl. Acad. Sci. USA 2013, 110, 1452–1457. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Xie, J.; Fu, Y.; Cheng, J.; Qu, Z.; Zhao, Z.; Cheng, S.; Chen, T.; Li, B.; Wang, Q.; et al. A 2-kb mycovirus converts a pathogenic fungus into a beneficial endophyte for brassica protection and yield enhancement. Mol. Plant. 2020, 13, 1420–1433. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Li, X.; Kotta-Loizou, I.; Dong, K.; Li, S.; Ni, D.; Hong, N.; Wang, G.; Xu, W. A mycovirus modulates the endophytic and pathogenic traits of a plant associated fungus. ISME J. 2021, 15, 1893–1906. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Shi, M.; Tian, J.; Lin, X.; Kang, Y.; Chen, L.; Qin, X.; Xu, J.; Holmes, E.C.; Zhang, Y. Unprecedented genomic diversity of RNA viruses in arthropods reveals the ancestry of negative-sense RNA viruses. eLife 2015, 4, e05378. [Google Scholar] [CrossRef] [PubMed]
- Marzano, S.L.; Domier, L.L. Novel mycoviruses discovered from metatranscriptomics survey of soybean phyllosphere phytobiomes. Virus Res. 2016, 213, 332–342. [Google Scholar] [CrossRef] [PubMed]
- Arjona-Lopez, J.M.; Telengech, P.; Jamal, A.; Hisano, S.; Kondo, H.; Yelin, M.D.; Arjona-Girona, I.; Kanematsu, S.; Lopez-Herrera, C.J.; Suzuki, N. Novel, diverse RNA viruses from mediterranean isolates of the phytopathogenic fungus, Rosellinia necatrix: Insights into evolutionary biology of fungal viruses. Environ. Microbiol. 2018, 20, 1464–1483. [Google Scholar] [CrossRef] [PubMed]
- Chiapello, M.; Rodríguez-Romero, J.; Ayllón, M.A.; Turina, M. Analysis of the virome associated to grapevine downy mildew lesions reveals new mycovirus lineages. Virus Evol. 2020, 6, veaa058. [Google Scholar] [CrossRef]
- Yang, S.; Dai, R.; Salaipeth, L.; Huang, L.; Liu, J.; Andika-Ida, B.; Sun, L. Infection of two heterologous mycoviruses reduces the virulence of Valsa mali, a fungal agent of apple Valsa canker disease. Front. Microbiol. 2021, 12, 659210. [Google Scholar] [CrossRef]
- Schenk, J.J.; Becklund, L.E.; Carey, S.J.; Fabre, P.P. What is the modified CTAB protocol? Characterizing modifications to the CTAB DNA extraction protocol. Appl. Plant Sci. 2023, 11, e11517. [Google Scholar] [CrossRef]
- Bartholomäus, A.; Wibberg, D.; Winkler, A.; Pühler, A.; Varrelmann, M. Deep sequencing analysis reveals the mycoviral diversity of the virome of an avirulent isolate of Rhizoctonia solani AG-2-2 IV. PLoS ONE 2016, 11, e0165965. [Google Scholar] [CrossRef] [PubMed]
- Marzano, S.L.; Nelson, B.D.; Ajayi-Oyetunde, O.; Bradley, C.A.; Hughes, T.J.; Hartman, G.L.; Eastburn, D.M.; Domier, L.L. Identification of diverse mycoviruses through metatranscriptomics characterization of the viromes of five major fungal plant pathogens. J. Virol. 2016, 90, 6846–6863. [Google Scholar] [CrossRef] [PubMed]
- Mu, F.; Xie, J.; Cheng, S.; You, M.; Barbetti, M.J.; Jia, J.; Wang, Q.; Cheng, J.; Fu, Y.; Chen, T.; et al. Virome characterization of a collection of S. sclerotiorum from Australia. Front. Microbiol. 2017, 8, 2540. [Google Scholar] [CrossRef]
- Hideki, K.; Satoko, K.; Nobuhiro, S. Viruses of the white root rot fungus, Rosellinia necatrix. Adv. Virus Res. 2013, 86, 177–214. [Google Scholar] [CrossRef]
- Qu, Z.; Zhao, H.; Zhang, H.; Wang, Q.; Yao, Y.; Cheng, J.; Lin, Y.; Xie, J.; Fu, Y.; Jiang, D. Bio-priming with a hypovirulent phytopathogenic fungus enhances the connection and strength of microbial interaction network in rapeseed. NPJ Bioflms Microbiomes 2020, 6, 45. [Google Scholar] [CrossRef] [PubMed]
- Kondo, H.; Botella, L.; Suzuki, N. Mycovirus diversity and evolution revealed/inferred from recent studies. Annu. Rev. Phytopathol. 2022, 60, 307–336. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Suzuki, N. Continued mycovirus discovery expanding our understanding of virus lifestyles, symptom expression, and host defense. Curr. Opin. Microbiol. 2023, 75, 102337. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Cheng, S.; Xiao, X.; Cheng, J.; Fu, Y.; Chen, T.; Jiang, D.; Xie, J. Discovery of two mycoviruses by high-throughput sequencing and assembly of mycovirus-derived small silencing RNAs from a hypovirulent strain of Sclerotinia sclerotiorum. Front. Microbiol. 2019, 10, 1415. [Google Scholar] [CrossRef]
- Ye, T.; Lu, Z.; Li, H.; Duan, J.; Hai, D.; Lin, Y.; Xie, J.; Cheng, J.; Li, B.; Chen, T.; et al. Characterization of a fungal virus rep-resenting a novel genus in the family alphaflexiviridae. Viruses 2023, 15, 339. [Google Scholar] [CrossRef]
- Koloniuk, I.; El-Habbak, M.H.; Petrzik, K.; Ghabrial, S.A. Complete genome sequence of a novel hypovirus infecting phomopsis longicolla. Arch. Virol. 2014, 159, 1861–1863. [Google Scholar] [CrossRef]
- Wu, S.; Cheng, J.; Fu, Y.; Chen, T.; Jiang, D.; Ghabrial, S.A.; Xie, J. Virus-mediated suppression of host non-self recognition facilitates horizontal transmission of heterologous viruses. PLoS Pathog. 2017, 3, e1006234. [Google Scholar] [CrossRef]
- Zhang, R.; Hisano, S.; Tani, A.; Kondo, H.; Kanematsu, S.; Suzuki, N. A capsidless ssRNA virus hosted by an unrelated dsRNA virus. Nat. Microbiol. 2016, 1, 15001. [Google Scholar] [CrossRef]
- Ayllón, M.A.; Turina, M.; Xie, J.; Nerva, L.; Marzano, S.L.; Donaire, L.; Jiang, D. ICTV virus taxonomy profile: Botourmiaviridae. J. Gen. Virol. 2020, 101, 454–455. [Google Scholar] [CrossRef]
- Rui, Z.; Liu, S.; Sotaro, C.; Hideki, K.; Satoko, K.; Nobuhiro, S. A novel single-stranded RNA virus isolated from a phytopathogenic filamentous fungus, Rosellinia necatrix, with similarity to hypo-like viruses. Front. Microbiol. 2014, 5, 360. [Google Scholar] [CrossRef]
- Livia, D.; Julio, R.; María, A.A. Molecular characterization of Botrytis ourmia-like virus, a mycovirus close to the plant pathogenic genus Ourmiavirus. Virology 2016, 489, 158–164. [Google Scholar] [CrossRef]
- Zhong, J.; Zhao, S.; Li, G.; Pang, X.; Deng, X.; Zhu, H.; Zhou, Q. A novel fusarivirus isolated from the phytopathogenic fungus Nigrospora oryzae. Virus Genes 2016, 52, 891–895. [Google Scholar] [CrossRef]
- Marais, A.; Nivault, A.; Faure, C.; Comont, G.; Theil, S.; Candresse, T.; Corio-Costet, M. Molecular characterization of a novel fusarivirus infecting the plant-pathogenic fungus Neofusicoccum luteum. Arch. Virol. 2018, 163, 559–562. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Hai, D.; Mu, F.; Yu, X.; Zhao, Y.; He, B.; Xie, J.; Jiang, D.; Liu, H. Molecular characterization of a novel fusarivirus infecting the plant-pathogenic fungus Botryosphaeria dothidea. Arch Virol. 2020, 165, 1033–1037. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Cai, L.; Liu, M.; Wang, X.; Yang, J.; An, H.; Deng, Q.; Zhang, S.; Fang, S. A novel previously undescribed fusarivirus from the phytopathogenic fungus Setosphaeria turcica. Arch Virol. 2021, 166, 665–669. [Google Scholar] [CrossRef] [PubMed]
- Gong, W.; Liu, H.; Zhu, X.; Zhao, S.; Cheng, J.; Zhu, H.; Zhong, J.; Zhou, Q. Molecular characterization of a novel fusarivirus infecting the plant-pathogenic fungus Alternaria solani. Arch Virol. 2021, 1166, 2063–2067. [Google Scholar] [CrossRef] [PubMed]
- Zhong, J.; Shang, H.; Zhu, X.; Zhu, J.; Hu, Y.; Gao, B. Characterization of a novel single-stranded RNA virus, closely related to fusariviruses, infecting the plant pathogenic fungus Alternaria brassicicola. Virus Res. 2016, 217, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Ghabrial, S.A.; Suzuki, N. Viruses of plant pathogenic fungi. Annu. Rev. Phytopathol. 2009, 47, 353–384. [Google Scholar] [CrossRef] [PubMed]
- Ghabrial, S.A.; Dunn, S.E.; Li, H.; Xie, J.; Baker, T.S. Viruses of helminthosporium (cochlioblus) victoriae. Adv. Virus Res. 2013, 86, 289–325. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Contig Number | Contig Length | Name of Putative Viruses | Best Match | aa Identity | Genome Type | Family/Genus |
|---|---|---|---|---|---|---|---|
| 1 | Bcontig1 | 9810 | Phomopsis longicolla hypovirus (PlHV) | Phomopsis longicolla hypovirus YP_009051683.1 | 99% | +SSRNA | Hypoviridae |
| 2 | Acontig5 | 2969 | Cryphonectria hypovirus 3-XJ1(CHV3-XJ1) | Cryphonectria hypovirus 3 NP_051710.1 | 81% | +SSRNA | Hypoviridae |
| 3 | Bcontig11 | 2519 | Botourmiaviridae sp.-XJ1(BVsp-XJ1) | Botourmiaviridae sp. UJQ91985.1 | 83% | +SSRNA | Botourmiaviridae |
| 4 | Ccontig6 | 6046 | Fusarium asiaticum fusarivirus 1-XJ1(FaFV1-XJ1) | Fusarium asiaticum fusarivirus 1UNG44326.1 | 98% | +SSRNA | Fusariviridae |
| 5 | Ccontig35 | 653 | Fusariviridae sp.-XJ1(Fvsp-XJ1) | Fusariviridae sp. WAK72331.1 | 57% | +SSRNA | Fusariviridae |
| 6 | Acontig4 | 3046 | Cytospora ribis mitovirus 1 (CrMV1) | Cytospora ribis mitovirus 1 AIS37555.1 | 100% | +SSRNA | Mitoviridae |
| 7 | Ccontig33 | 665 | Magnaporthe oryzae narnavirus 1-XJ1(MoNV1-XJ1) | Magnaporthe oryzae narnavirus 1BCH36657.1 | 98% | +SSRNA | Narnaviridae |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, C.; Zheng, X.; Tie, Z.; Xi, H.; Shi, M.; Ma, Y.; Chen, W.; Mi, Y.; Yang, R.; Zhao, S.; et al. Identification of Mycoviruses in the Pathogens of Fragrant Pear Valsa Canker from Xinjiang in China. Viruses 2024, 16, 355. https://doi.org/10.3390/v16030355
Zhang C, Zheng X, Tie Z, Xi H, Shi M, Ma Y, Chen W, Mi Y, Yang R, Zhao S, et al. Identification of Mycoviruses in the Pathogens of Fragrant Pear Valsa Canker from Xinjiang in China. Viruses. 2024; 16(3):355. https://doi.org/10.3390/v16030355
Chicago/Turabian StyleZhang, Chenguang, Xiaoya Zheng, Zhanjiang Tie, Hui Xi, Mai Shi, Yanjun Ma, Wenbin Chen, Yingjie Mi, Rui Yang, Sifeng Zhao, and et al. 2024. "Identification of Mycoviruses in the Pathogens of Fragrant Pear Valsa Canker from Xinjiang in China" Viruses 16, no. 3: 355. https://doi.org/10.3390/v16030355
APA StyleZhang, C., Zheng, X., Tie, Z., Xi, H., Shi, M., Ma, Y., Chen, W., Mi, Y., Yang, R., Zhao, S., & Zhang, X. (2024). Identification of Mycoviruses in the Pathogens of Fragrant Pear Valsa Canker from Xinjiang in China. Viruses, 16(3), 355. https://doi.org/10.3390/v16030355