
Fast-Track Discovery of SARS-CoV-2-Neutralizing Antibodies from Human B Cells by Direct Functional Screening
 , ,
, ,  ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Donors and Ethics
2.2. Expression, Purification and Fluorescent-Labelling of Coronavirus Spike RBD and huACE-2
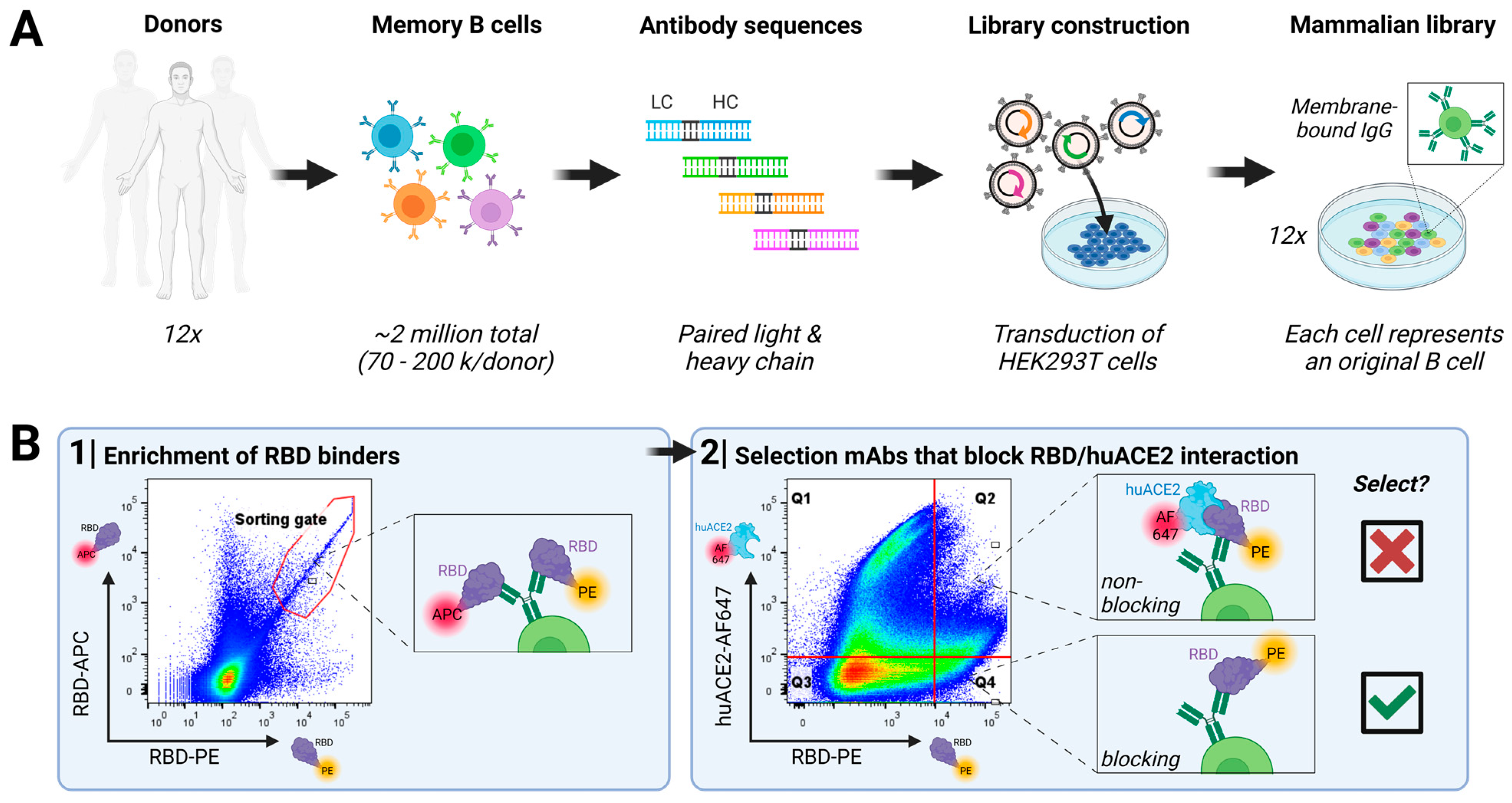
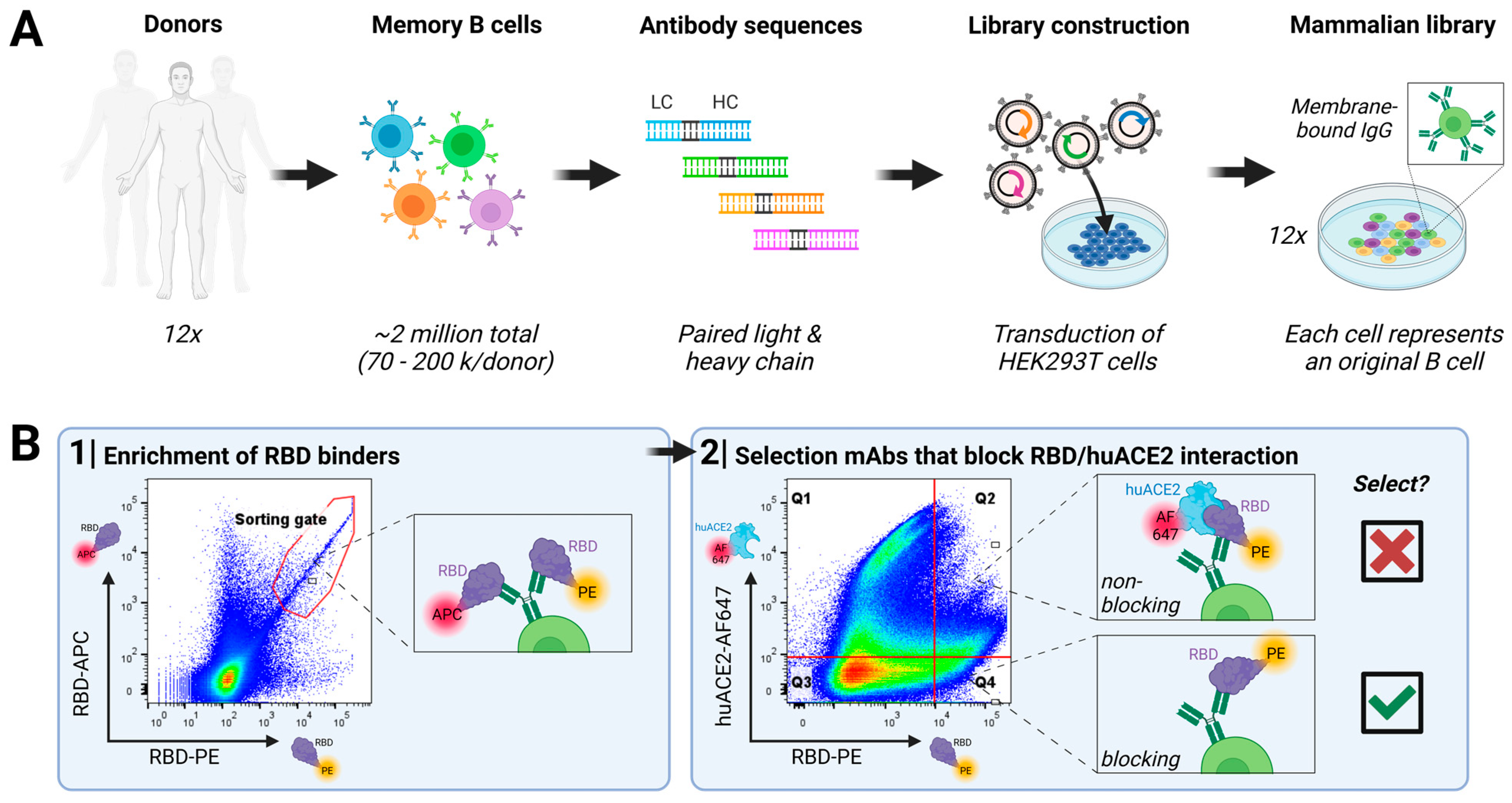
2.3. Antibody Discovery
2.4. Production of Human Monoclonal Antibodies
2.5. Preparation of Pseudo-Typed SARS-CoV-2 (SARS-CoPsV-2)
2.6. Neutralization of SARS-CoPsV-2
2.7. Binding to Variants of SARS-CoV-2 Spike
2.8. Neutralization of Wild-Type SARS-CoV-2
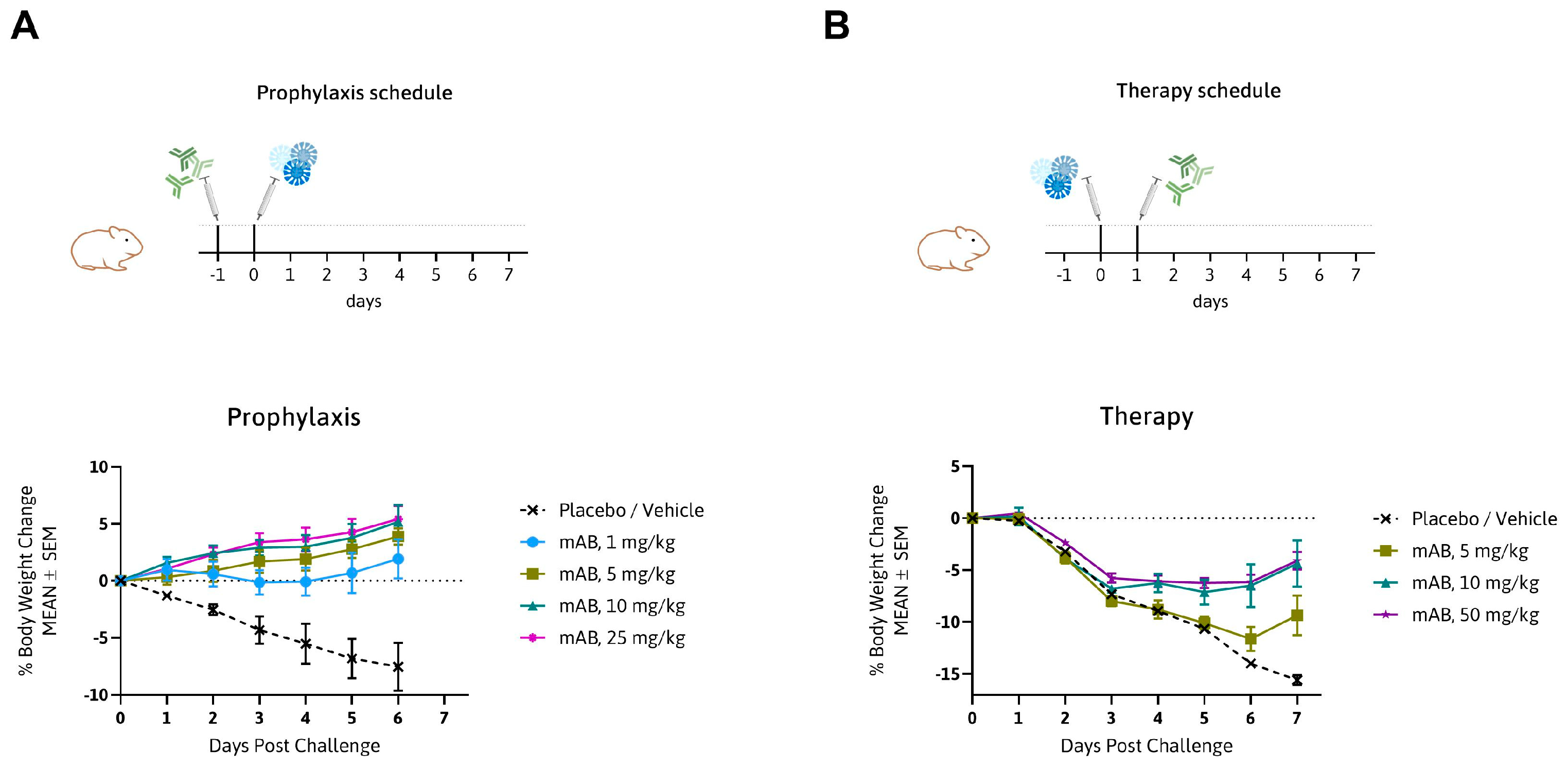
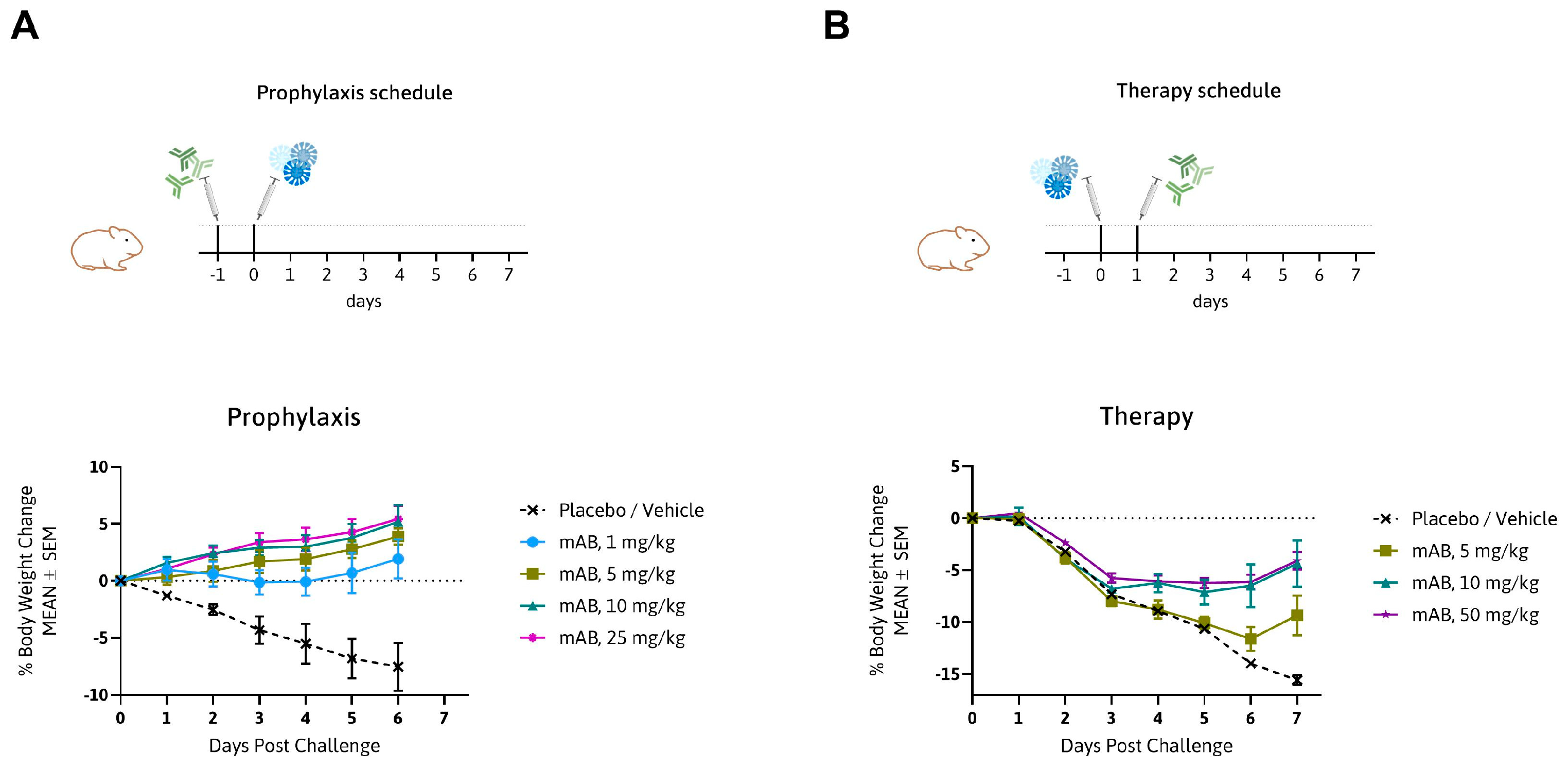
2.9. In Vivo Experiments (Hamster)
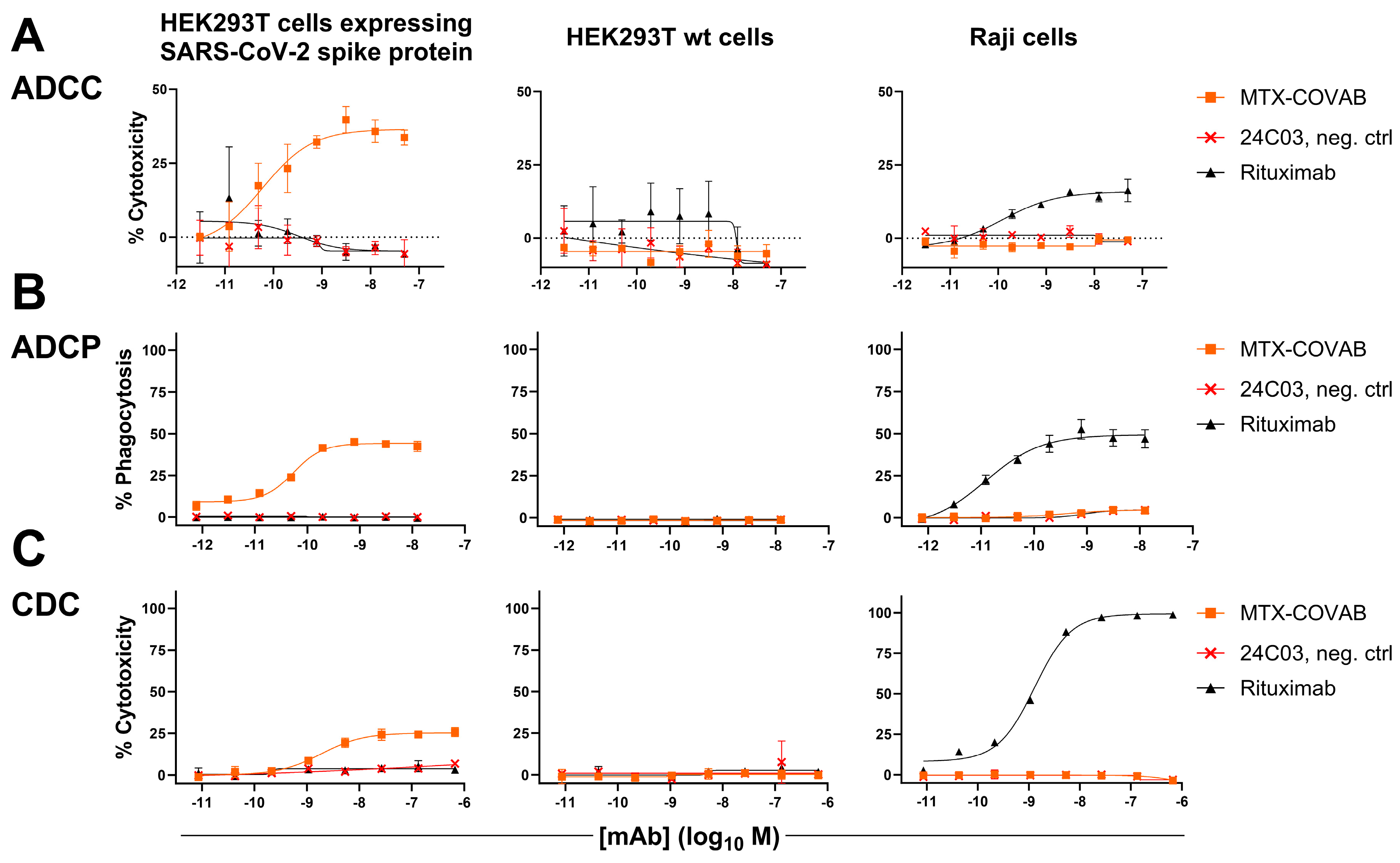
2.10. Antibody-Dependent Cellular Cytotoxicity (ADCC)
2.11. Antibody-Dependent Cellular Phagocytosis (ADCP)
2.12. Complement-Dependent Cytotoxicity (CDC)
3. Results
3.1. Direct Discovery of SARS-CoV-2-Neutralizing Antibodies
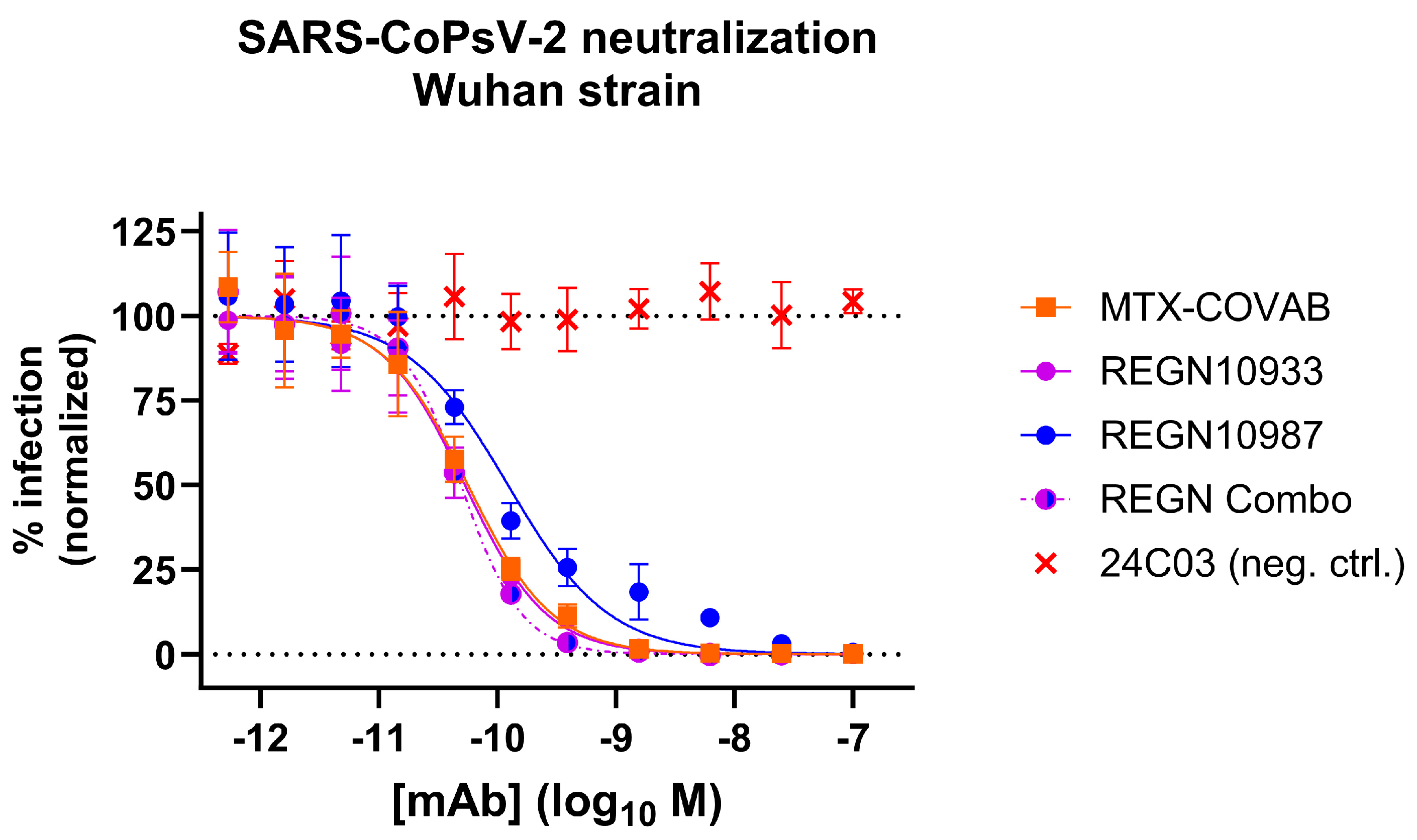
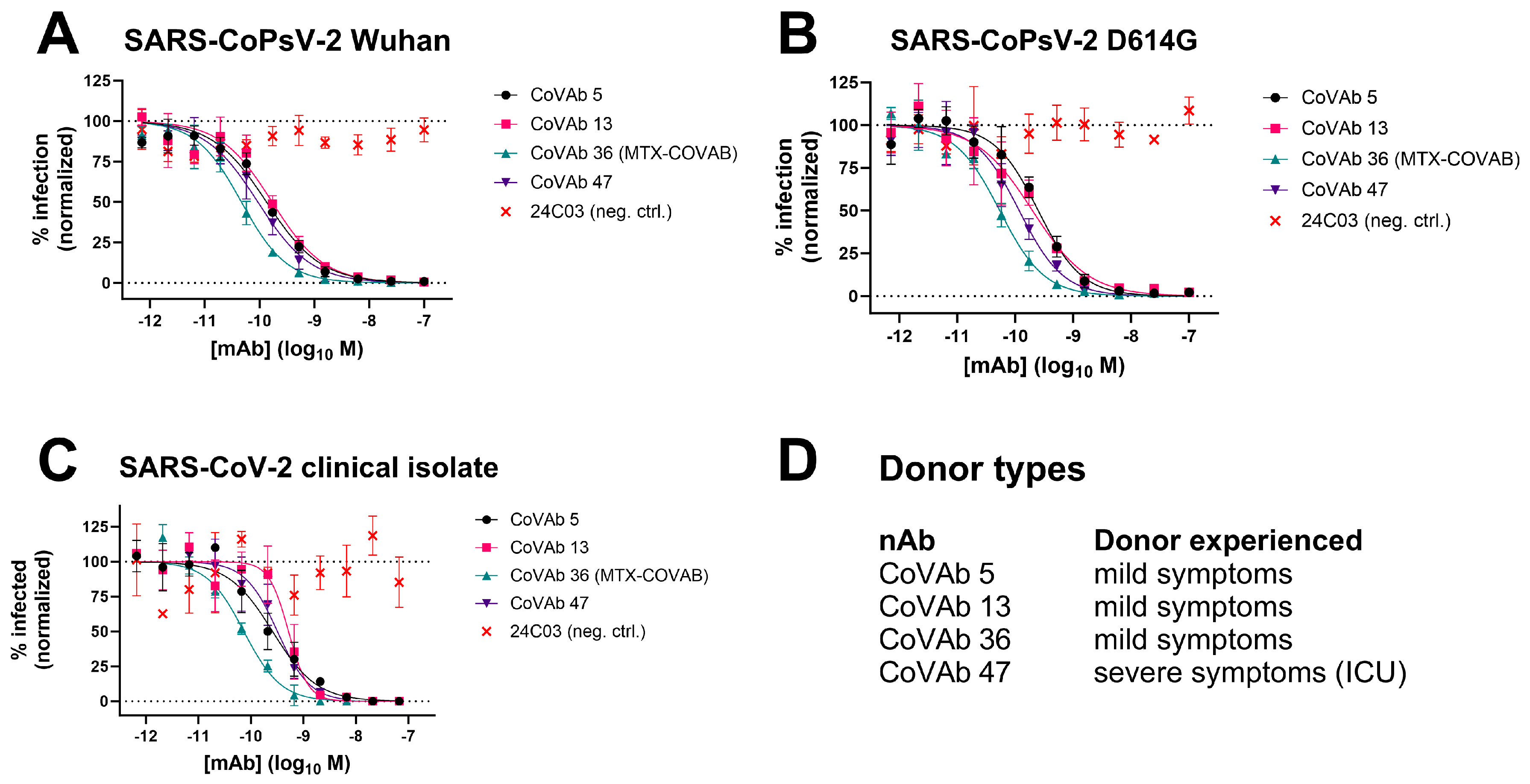
3.2. Characterization of Antibodies in Neutralization of SARS-CoPsV-2 and Clinical Isolate
3.3. MTX-COVAB Neutralizes Wild-Type SARS-CoV-2 Efficiently in an In Vivo Hamster Model
3.4. Fc-Effector Functions
3.5. Comparison to Other Neutralizing Antibodies
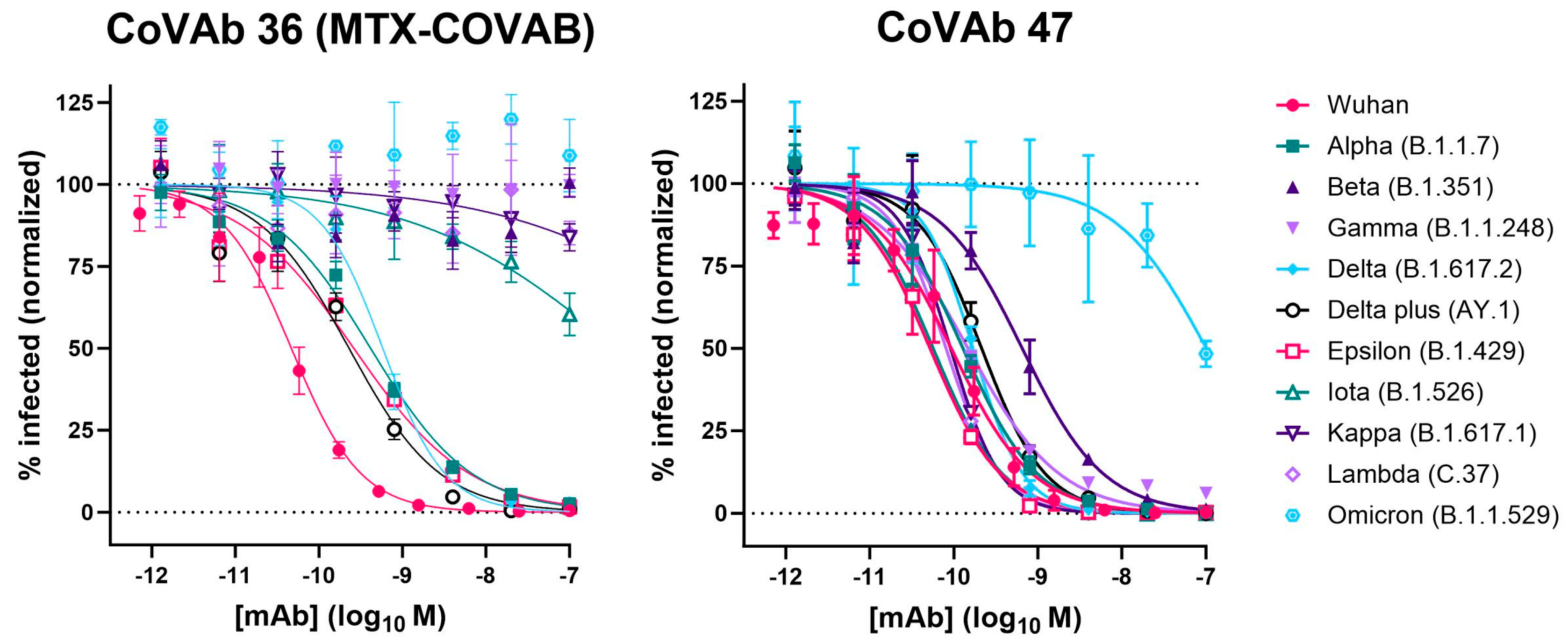
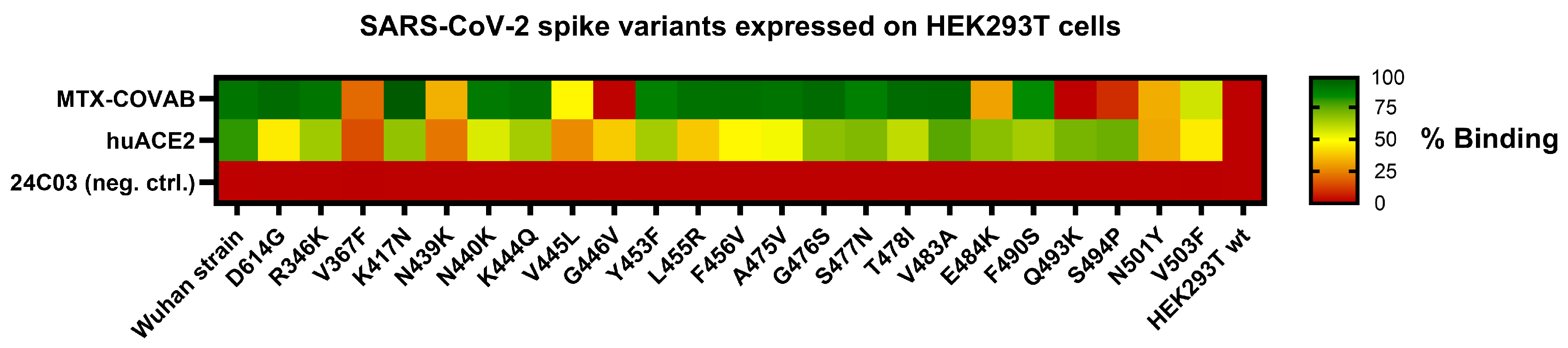
3.6. Spike Mutant Binding and Neutralization
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhou, P.; Yang, X.-L.; Wang, X.-G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.-R.; Zhu, Y.; Li, B.; Huang, C.-L.; et al. A Pneumonia Outbreak Associated with a New Coronavirus of Probable Bat Origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [PubMed]
- Baum, A.; Fulton, B.O.; Wloga, E.; Copin, R.; Pascal, K.E.; Russo, V.; Giordano, S.; Lanza, K.; Negron, N.; Ni, M.; et al. Antibody Cocktail to SARS-CoV-2 Spike Protein Prevents Rapid Mutational Escape Seen with Individual Antibodies. Science 2020, 369, 1014–1018. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Su, B.; Guo, X.; Sun, W.; Deng, Y.; Bao, L.; Zhu, Q.; Zhang, X.; Zheng, Y.; Geng, C.; et al. Potent Neutralizing Antibodies against SARS-CoV-2 Identified by High-Throughput Single-Cell Sequencing of Convalescent Patients’ B Cells. Cell 2020, 182, 73–84.e16. [Google Scholar] [CrossRef]
- Hansen, J.; Baum, A.; Pascal, K.E.; Russo, V.; Giordano, S.; Wloga, E.; Fulton, B.O.; Yan, Y.; Koon, K.; Patel, K.; et al. Studies in Humanized Mice and Convalescent Humans Yield a SARS-CoV-2 Antibody Cocktail. Science 2020, 369, 1010–1014. [Google Scholar] [CrossRef]
- Ju, B.; Zhang, Q.; Ge, J.; Wang, R.; Sun, J.; Ge, X.; Yu, J.; Shan, S.; Zhou, B.; Song, S.; et al. Human Neutralizing Antibodies Elicited by SARS-CoV-2 Infection. Nature 2020, 584, 115–119. [Google Scholar] [CrossRef]
- Wang, C.; Li, W.; Drabek, D.; Okba, N.M.A.; van Haperen, R.; Osterhaus, A.D.M.E.; van Kuppeveld, F.J.M.; Haagmans, B.L.; Grosveld, F.; Bosch, B.-J. A Human Monoclonal Antibody Blocking SARS-CoV-2 Infection. Nat. Commun. 2020, 11, 2251. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Wang, P.; Nair, M.S.; Yu, J.; Rapp, M.; Wang, Q.; Luo, Y.; Chan, J.F.-W.; Sahi, V.; Figueroa, A.; et al. Potent Neutralizing Antibodies Directed to Multiple Epitopes on SARS-CoV-2 Spike. Nature 2020, 584, 450–456. [Google Scholar] [CrossRef] [PubMed]
- Shi, R.; Shan, C.; Duan, X.; Chen, Z.; Liu, P.; Song, J.; Song, T.; Bi, X.; Han, C.; Wu, L.; et al. A Human Neutralizing Antibody Targets the Receptor Binding Site of SARS-CoV-2. Nature 2020, 584, 120–124. [Google Scholar] [CrossRef]
- Jones, B.E.; Brown-Augsburger, P.L.; Corbett, K.S.; Westendorf, K.; Davies, J.; Cujec, T.P.; Wiethoff, C.M.; Blackbourne, J.L.; Heinz, B.A.; Foster, D.; et al. LY-CoV555, a Rapidly Isolated Potent Neutralizing Antibody, Provides Protection in a Non-Human Primate Model of SARS-CoV-2 Infection. BioRxiv 2020. [Google Scholar]
- Zost, S.J.; Gilchuk, P.; Chen, R.E.; Case, J.B.; Reidy, J.X.; Trivette, A.; Nargi, R.S.; Sutton, R.E.; Suryadevara, N.; Chen, E.C.; et al. Rapid Isolation and Profiling of a Diverse Panel of Human Monoclonal Antibodies Targeting the SARS-CoV-2 Spike Protein. Nat. Med. 2020, 26, 1422–1427. [Google Scholar] [CrossRef]
- Singh, M.; Bansal, V.; Feschotte, C. A Single-Cell RNA Expression Map of Human Coronavirus Entry Factors. Cell Rep. 2020, 32, 108175. [Google Scholar] [CrossRef]
- Tragni, V.; Preziusi, F.; Laera, L.; Onofrio, A.; Mercurio, I.; Todisco, S.; Volpicella, M.; De Grassi, A.; Pierri, C.L. Modeling SARS-CoV-2 Spike/ACE2 Protein–Protein Interactions for Predicting the Binding Affinity of New Spike Variants for ACE2, and Novel ACE2 Structurally Related Human Protein Targets, for COVID-19 Handling in the 3PM Context. EPMA J. 2022, 13, 149–175. [Google Scholar] [CrossRef]
- Wrapp, D.; Wang, N.; Corbett, K.S.; Goldsmith, J.A.; Hsieh, C.-L.; Abiona, O.; Graham, B.S.; McLellan, J.S. Cryo-EM Structure of the 2019-nCoV Spike in the Prefusion Conformation. Science 2020, 367, 1260–1263. [Google Scholar] [CrossRef] [PubMed]
- Yan, R.; Zhang, Y.; Li, Y.; Xia, L.; Guo, Y.; Zhou, Q. Structural Basis for the Recognition of the SARS-CoV-2 by Full-Length Human ACE2. Science 2020, 367, 1444–1448. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zhang, Y.; Wu, L.; Niu, S.; Song, C.; Zhang, Z.; Lu, G.; Qiao, C.; Hu, Y.; Yuen, K.-Y.; et al. Structural and Functional Basis of SARS-CoV-2 Entry by Using Human ACE2. Cell 2020, 181, 894–904.e9. [Google Scholar] [CrossRef]
- Deshpande, A.; Harris, B.D.; Martinez-Sobrido, L.; Kobie, J.J.; Walter, M.R. Epitope Classification and RBD Binding Properties of Neutralizing Antibodies Against SARS-CoV-2 Variants of Concern. Front. Immunol. 2021, 12, 691715. [Google Scholar] [CrossRef]
- Pierri, C.L. SARS-CoV-2 Spike Protein: Flexibility as a New Target for Fighting Infection. Signal Transduct. Target. Ther. 2020, 5, 254. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Zhou, J.; Tian, M.; Huang, M.; Liu, S.; Xie, Y.; Han, P.; Bai, C.; Han, P.; Zheng, A.; et al. Omicron SARS-CoV-2 Mutations Stabilize Spike up-RBD Conformation and Lead to a Non-RBM-Binding Monoclonal Antibody Escape. Nat. Commun. 2022, 13, 4958. [Google Scholar] [CrossRef]
- Regeneron Pharmaceuticals: Delivering Life-Transforming Medicines. Available online: https://www.regeneron.com/covid19 (accessed on 24 January 2024).
- Update on US Food and Drug Administration Emergency Use Authorisation of Evusheld. Available online: https://www.astrazeneca.com/media-centre/press-releases/2023/update-on-evusheld-us-eua.html (accessed on 26 January 2024).
- January 24, 2022|Important Update|HHS/ASPR. Available online: https://aspr.hhs.gov:443/COVID-19/Therapeutics/updates/Pages/important-update-24January2022.aspx (accessed on 26 January 2024).
- Wang, X.; Stollar, B.D. Human Immunoglobulin Variable Region Gene Analysis by Single Cell RT-PCR. J. Immunol. Methods 2000, 244, 217–225. [Google Scholar] [CrossRef]
- Wardemann, H. Predominant Autoantibody Production by Early Human B Cell Precursors. Science 2003, 301, 1374–1377. [Google Scholar] [CrossRef]
- Ou, X.; Liu, Y.; Lei, X.; Li, P.; Mi, D.; Ren, L.; Guo, L.; Guo, R.; Chen, T.; Hu, J.; et al. Characterization of Spike Glycoprotein of SARS-CoV-2 on Virus Entry and Its Immune Cross-Reactivity with SARS-CoV. Nat. Commun. 2020, 11, 1620. [Google Scholar] [CrossRef]
- Walls, A.C.; Park, Y.-J.; Tortorici, M.A.; Wall, A.; McGuire, A.T.; Veesler, D. Structure, Function, and Antigenicity of the SARS-CoV-2 Spike Glycoprotein. Cell 2020, 181, 281–292.e6. [Google Scholar] [CrossRef]
- Petit, C.M.; Melancon, J.M.; Chouljenko, V.N.; Colgrove, R.; Farzan, M.; Knipe, D.M.; Kousoulas, K.G. Genetic Analysis of the SARS-Coronavirus Spike Glycoprotein Functional Domains Involved in Cell-Surface Expression and Cell-to-Cell Fusion. Virology 2005, 341, 215–230. [Google Scholar] [CrossRef] [PubMed]
- Laha, S.; Chakraborty, J.; Das, S.; Manna, S.K.; Biswas, S.; Chatterjee, R. Characterizations of SARS-CoV-2 Mutational Profile, Spike Protein Stability and Viral Transmission. Infect. Genet. Evol. 2020, 85, 104445. [Google Scholar] [CrossRef] [PubMed]
- Imai, M.; Iwatsuki-Horimoto, K.; Hatta, M.; Loeber, S.; Halfmann, P.J.; Nakajima, N.; Watanabe, T.; Ujie, M.; Takahashi, K.; Ito, M.; et al. Syrian Hamsters as a Small Animal Model for SARS-CoV-2 Infection and Countermeasure Development. Proc. Natl. Acad. Sci. USA 2020, 117, 16587–16595. [Google Scholar] [CrossRef] [PubMed]
- Kreye, J.; Reincke, S.M.; Kornau, H.-C.; Sánchez-Sendin, E.; Corman, V.M.; Liu, H.; Yuan, M.; Wu, N.C.; Zhu, X.; Lee, C.-C.D.; et al. A Therapeutic Non-Self-Reactive SARS-CoV-2 Antibody Protects from Lung Pathology in a COVID-19 Hamster Model. Cell 2020, 183, 1058–1069.e19. [Google Scholar] [CrossRef] [PubMed]
- Mahita, J.; Ha, B.; Gambiez, A.; Schendel, S.L.; Li, H.; Hastie, K.M.; Dennison, S.M.; Li, K.; Kuzmina, N.; Periasamy, S.; et al. Coronavirus Immunotherapeutic Consortium Database. Database 2023, 2023, baac112. [Google Scholar] [CrossRef] [PubMed]
- Weisblum, Y.; Schmidt, F.; Zhang, F.; DaSilva, J.; Poston, D.; Lorenzi, J.C.C.; Muecksch, F.; Rutkowska, M.; Hoffmann, H.-H.; Michailidis, E.; et al. Escape from Neutralizing Antibodies by SARS-CoV-2 Spike Protein Variants. bioRxiv 2020. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wang, R.; Wang, M.; Wei, G.-W. Mutations Strengthened SARS-CoV-2 Infectivity. J. Mol. Biol. 2020, 432, 5212–5226. [Google Scholar] [CrossRef]
- Wise, J. COVID-19: New Coronavirus Variant Is Identified in UK. BMJ 2020, 371, m4857. [Google Scholar] [CrossRef]
- GISAID Initiative—Tracking of Variants. Available online: https://www.gisaid.org/hcov19-variants/ (accessed on 24 January 2024).
- Tan, K.W.; Ji, P.; Qian, Z.; Gao, Q.; Wang, S.; Li, Q.; Gu, M.; Zhang, Q.; Hou, C.; Huang, Y.; et al. Rapidly Accelerated Development of Neutralizing COVID-19 Antibodies by Reducing Cell Line and CMC Development Timelines. Biotech. Bioeng. 2022, bit.28302. [Google Scholar] [CrossRef]
- Deming, M.E.; Michael, N.L.; Robb, M.; Cohen, M.S.; Neuzil, K.M. Accelerating Development of SARS-CoV-2 Vaccines—The Role for Controlled Human Infection Models. N. Engl. J. Med. 2020, 383, e63. [Google Scholar] [CrossRef]
- Otsubo, R.; Yasui, T. Monoclonal Antibody Therapeutics for Infectious Diseases: Beyond Normal Human Immunoglobulin. Pharmacol. Ther. 2022, 240, 108233. [Google Scholar] [CrossRef]
- Winkler, E.S.; Gilchuk, P.; Yu, J.; Bailey, A.L.; Chen, R.E.; Chong, Z.; Zost, S.J.; Jang, H.; Huang, Y.; Allen, J.D.; et al. Human Neutralizing Antibodies against SARS-CoV-2 Require Intact Fc Effector Functions for Optimal Therapeutic Protection. Cell 2021, 184, 1804–1820.e16. [Google Scholar] [CrossRef]
- Taft, J.M.; Weber, C.R.; Gao, B.; Ehling, R.A.; Han, J.; Frei, L.; Metcalfe, S.W.; Overath, M.D.; Yermanos, A.; Kelton, W.; et al. Deep Mutational Learning Predicts ACE2 Binding and Antibody Escape to Combinatorial Mutations in the SARS-CoV-2 Receptor-Binding Domain. Cell 2022, 185, 4008–4022.e14. [Google Scholar] [CrossRef] [PubMed]
- Frei, L.; Gao, B.; Han, J.; Taft, J.M.; Irvine, E.B.; Weber, C.R.; Kumar, R.K.; Eisinger, B.N.; Reddy, S.T. Deep Learning-Guided Selection of Antibody Therapies with Enhanced Resistance to Current and Prospective SARS-CoV-2 Omicron Variants. bioRxiv 2023. [Google Scholar] [CrossRef]
- Tragni, V.; Mercurio, I.; Paoletti, D.P.; Onofrio, A.; Laera, L.; Cafferati Beltrame, L.; Sgobba, M.N.; Guerra, L.; Volpicella, M.; De Grassi, A.; et al. Deconstructing SARS-CoV-2 Neutralization: A Modular Molecular Framework for Computational Design and Comparison of Antibodies and Nanobodies Targeting the Spike RBD. J. Med. Virol. 2023, 95, e28875. [Google Scholar] [CrossRef]
- Han, W.; Chen, N.; Xu, X.; Sahil, A.; Zhou, J.; Li, Z.; Zhong, H.; Gao, E.; Zhang, R.; Wang, Y.; et al. Predicting the Antigenic Evolution of SARS-CoV-2 with Deep Learning. Nat. Commun. 2023, 14, 3478. [Google Scholar] [CrossRef] [PubMed]
- Johnson, N.V.; Wall, S.C.; Kramer, K.J.; Holt, C.M.; Periasamy, S.; Richardson, S.; Suryadevara, N.; Andreano, E.; Paciello, I.; Pierleoni, G.; et al. Discovery and Characterization of a Pan-Betacoronavirus S2-Binding Antibody. bioRxiv 2024. [Google Scholar] [CrossRef]
- Walls, A.C.; Sprouse, K.R.; Bowen, J.E.; Joshi, A.; Franko, N.; Navarro, M.J.; Stewart, C.; Cameroni, E.; McCallum, M.; Goecker, E.A.; et al. SARS-CoV-2 Breakthrough Infections Elicit Potent, Broad, and Durable Neutralizing Antibody Responses. Cell 2022, 185, 872–880.e3. [Google Scholar] [CrossRef]
- Guo, Y.; Zhang, G.; Yang, Q.; Xie, X.; Lu, Y.; Cheng, X.; Wang, H.; Liang, J.; Tang, J.; Gao, Y.; et al. Discovery and Characterization of Potent Pan-Variant SARS-CoV-2 Neutralizing Antibodies from Individuals with Omicron Breakthrough Infection. Nat. Commun. 2023, 14, 3537. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SARS-CoPsV-2 Wuhan | SARS-CoPsV-2 D614G | SARS-CoV-2 Clinical Isolate | ||||
|---|---|---|---|---|---|---|
| IC50 (nM) | 95% CI of IC50 (nM) | IC50 (nM) | 95% CI of IC50 (nM) | IC50 (nM) | 95% CI of IC50 (nM) | |
| CoVAb 5 | 0.134 | 0.108 to 0.165 | 0.250 | 0.199 to 0.313 | 0.260 | 0.193 to 0.350 |
| CoVAb 13 | 0.170 | 0.126 to 0.227 | 0.199 | 0.140 to 0.282 | 0.523 | 0.399 to 0.667 |
| CoVAb 36 (MTX-COVAB) | 0.047 | 0.039 to 0.056 | 0.054 | 0.045 to 0.064 | 0.074 | 0.060 to 0.092 |
| CoVAb 47 | 0.093 | 0.072 to 0.119 | 0.117 | 0.093 to 0.147 | 0.321 | 0.237 to 0.433 |
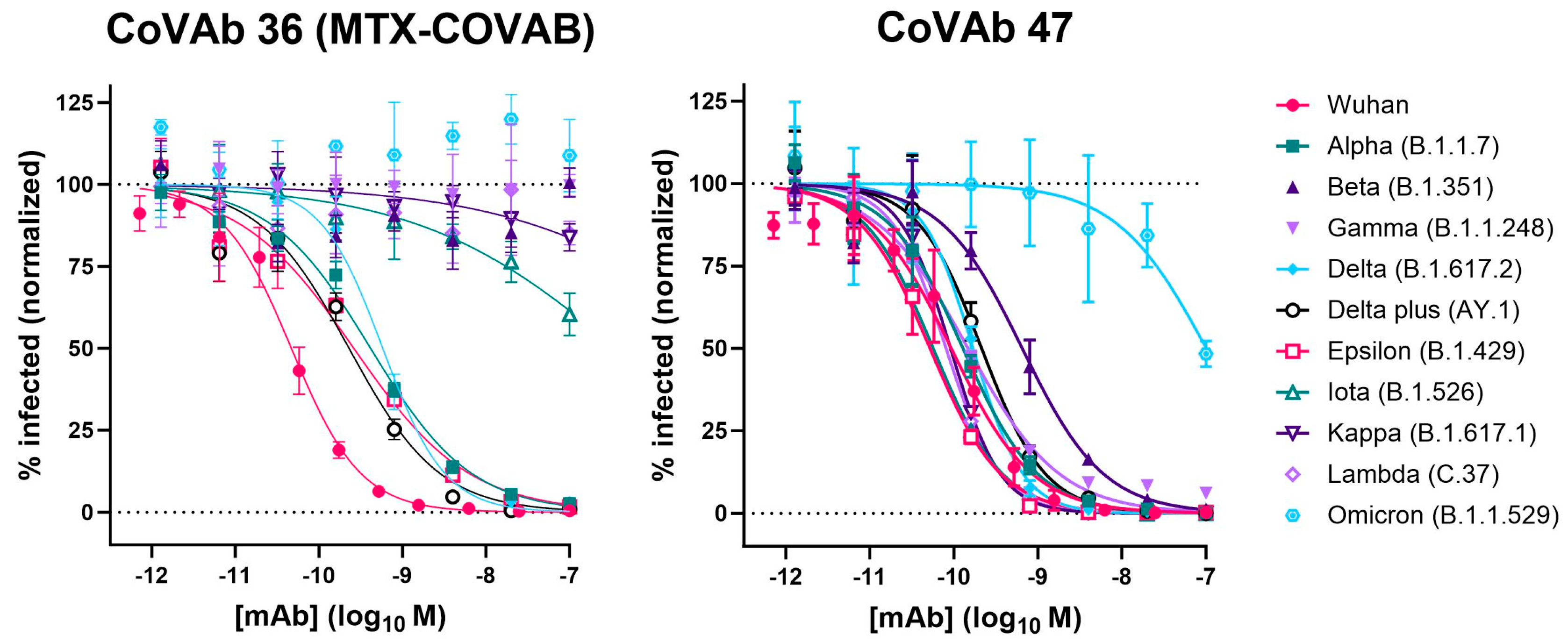
| MTX-COVAB | CoVAb 47 | |||
|---|---|---|---|---|
| IC50 (nM) | 95% CI of IC50 (nM) | IC50 (nM) | 95% CI of IC50 (nM) | |
| Wuhan | 0.047 | 0.039 to 0.056 | 0.093 | 0.072 to 0.119 |
| Alpha (B.1.1.7) | 0.415 | 0.340 to 0.506 | 0.129 | 0.104 to 0.159 |
| Beta (B.1.351) | ND | ND | 0.652 | 0.478 to 0.888 |
| Gamma (B.1.1.248) | ND | ND | 0.148 | 0.124 to 0.177 |
| Delta (B.1.617.2) | 0.581 | 0.461 to 0.736 | 0.169 | 0.126 to 0.226 |
| Delta plus (AY.1) | 0.229 | 0.161 to 0.322 | 0.214 | 0.165 to 0.278 |
| Epsilon (B.1.429) | 0.261 | 0.188 to 0.361 | 0.052 | 0.043 to 0.063 |
| Iota (B.1.526) | ~400 | 133.8 to 2319 | 0.057 | 0.051 to 0.063 |
| Kappa (B.1.617.1) | ND | ND | 0.091 | 0.079 to 0.105 |
| Lambda (C.37) | ND | ND | 0.081 | 0.068 to 0.096 |
| Omicron (B.1.1.529) | ND | ND | ~100 | 56.15 to 387.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hillenbrand, M.; Esslinger, C.; Seidenberg, J.; Weber, M.; Zingg, A.; Townsend, C.; Eicher, B.; Rutkauskaite, J.; Riese, P.; Guzman, C.A.; et al. Fast-Track Discovery of SARS-CoV-2-Neutralizing Antibodies from Human B Cells by Direct Functional Screening. Viruses 2024, 16, 339. https://doi.org/10.3390/v16030339
Hillenbrand M, Esslinger C, Seidenberg J, Weber M, Zingg A, Townsend C, Eicher B, Rutkauskaite J, Riese P, Guzman CA, et al. Fast-Track Discovery of SARS-CoV-2-Neutralizing Antibodies from Human B Cells by Direct Functional Screening. Viruses. 2024; 16(3):339. https://doi.org/10.3390/v16030339
Chicago/Turabian StyleHillenbrand, Matthias, Christoph Esslinger, Jemima Seidenberg, Marcel Weber, Andreas Zingg, Catherine Townsend, Barbara Eicher, Justina Rutkauskaite, Peggy Riese, Carlos A. Guzman, and et al. 2024. "Fast-Track Discovery of SARS-CoV-2-Neutralizing Antibodies from Human B Cells by Direct Functional Screening" Viruses 16, no. 3: 339. https://doi.org/10.3390/v16030339
APA StyleHillenbrand, M., Esslinger, C., Seidenberg, J., Weber, M., Zingg, A., Townsend, C., Eicher, B., Rutkauskaite, J., Riese, P., Guzman, C. A., Fischer, K., & Schmitt, S. (2024). Fast-Track Discovery of SARS-CoV-2-Neutralizing Antibodies from Human B Cells by Direct Functional Screening. Viruses, 16(3), 339. https://doi.org/10.3390/v16030339