
Human Rotavirus Replicates in Salivary Glands and Primes Immune Responses in Facial and Intestinal Lymphoid Tissues of Gnotobiotic Pigs

 ,
,  ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Virus Preparation
2.3. Rotavirus RT-qPCR
2.4. Detection of Infectious Rotavirus Viral Particles via Virus ELISPOT
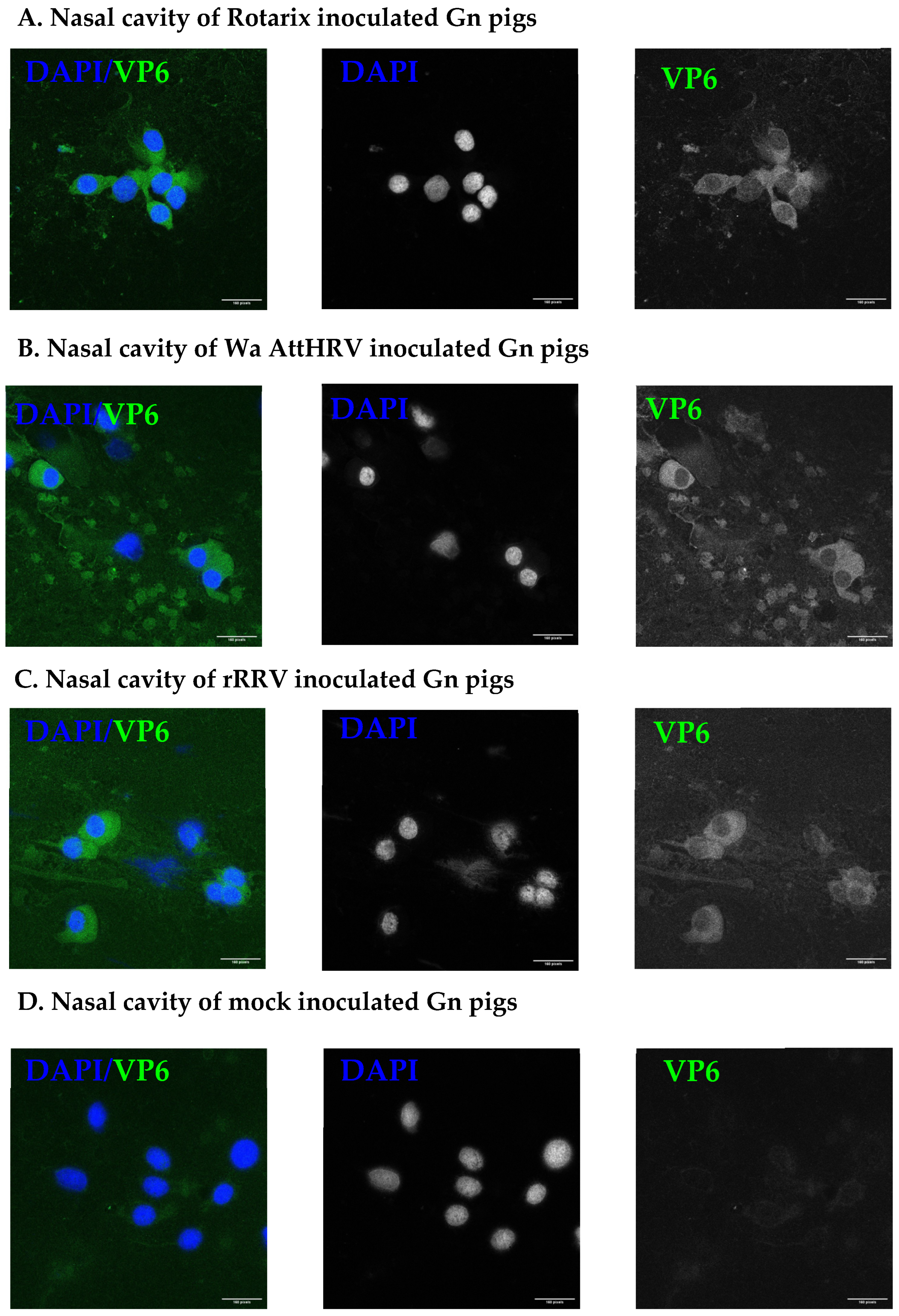
2.5. Immunostaining for Rotavirus in the Nasal Cavity
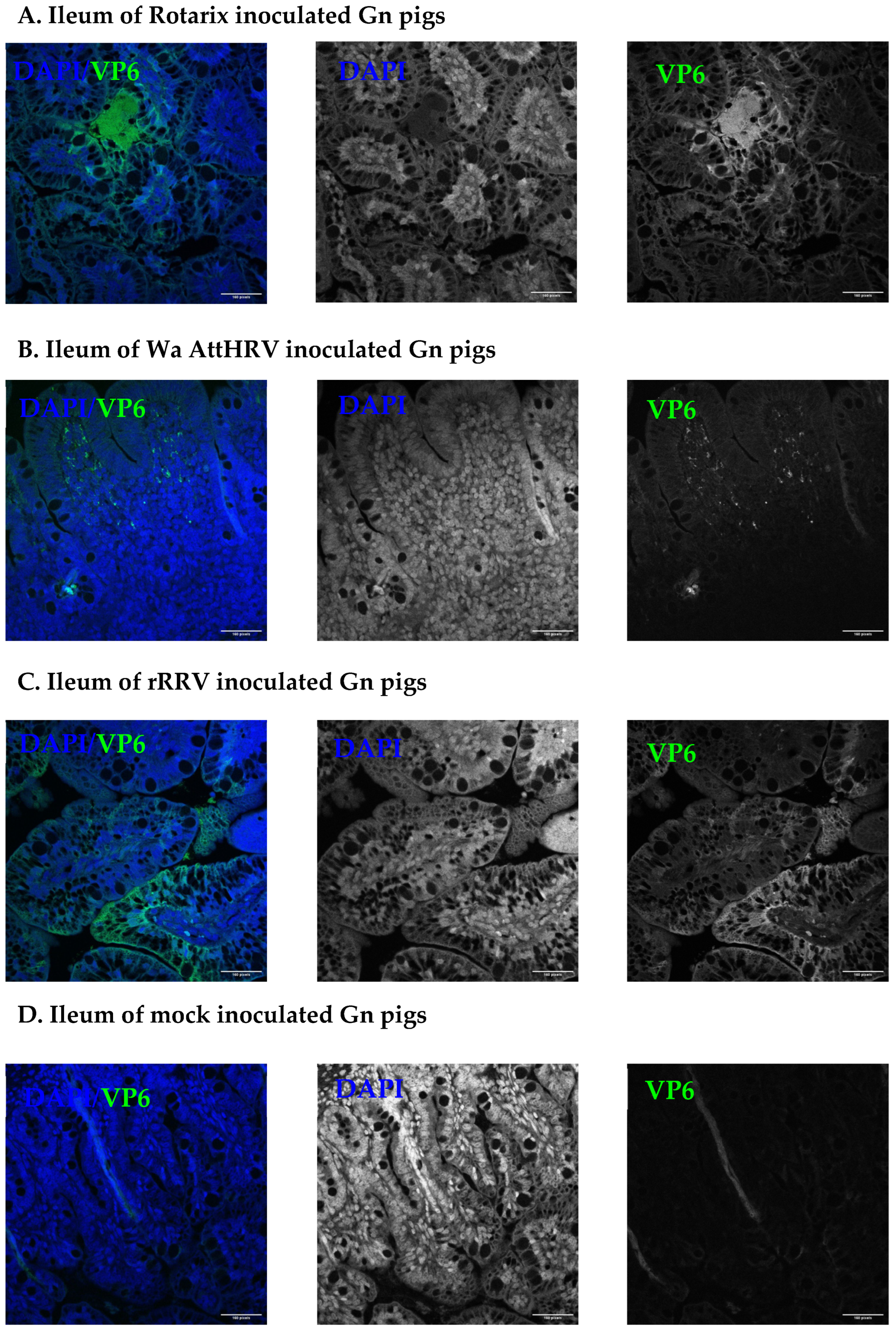
2.6. Immunostaining for Rotavirus in Salivary Glands and Ileum Sections
2.7. Detection of Virus-Specific IgM ASC by ELISPOT
2.8. Detection of Virus-Specific IgM by Antibody ELISA
3. Results
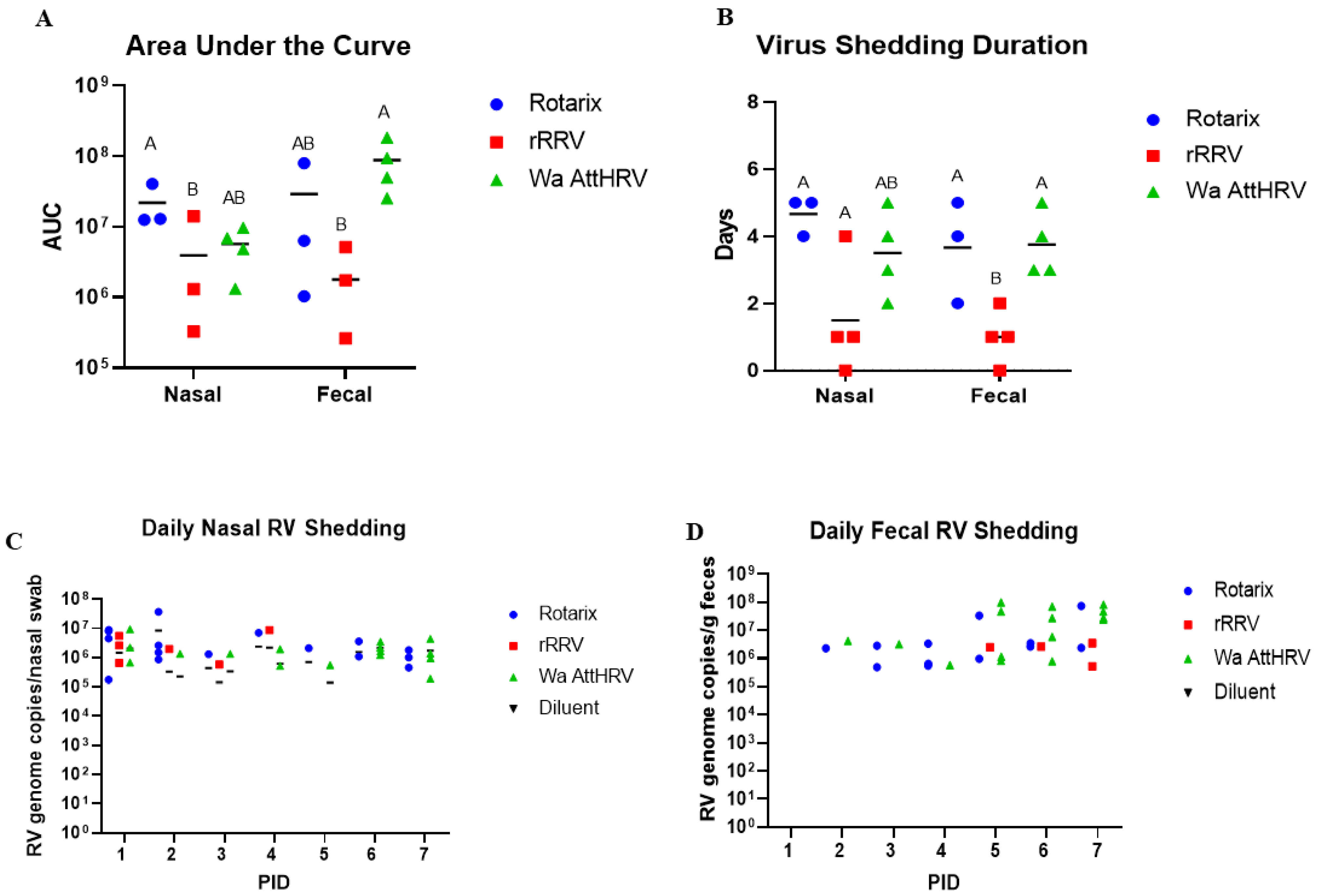
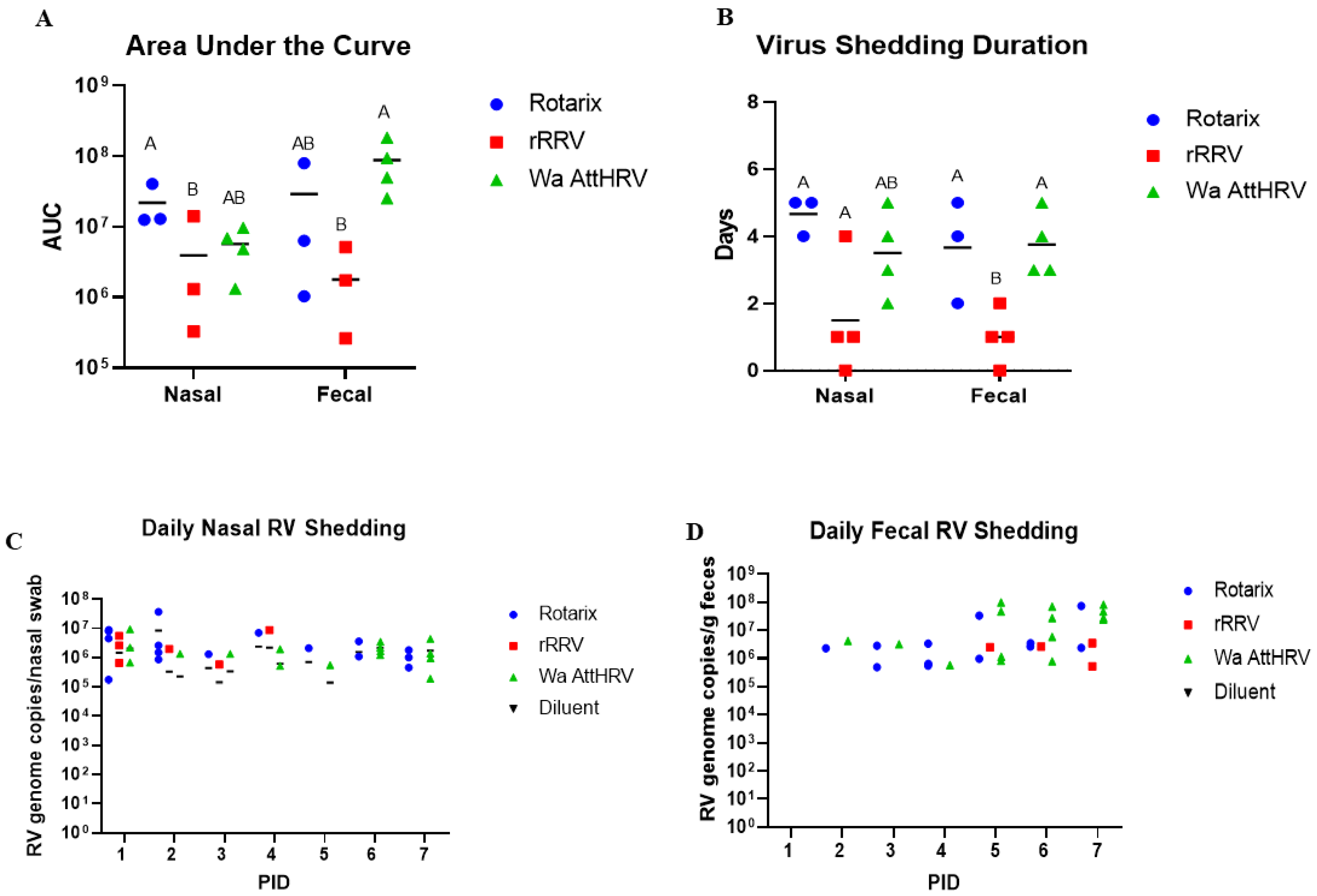
3.1. Nasal and Fecal Human Rotavirus Shedding
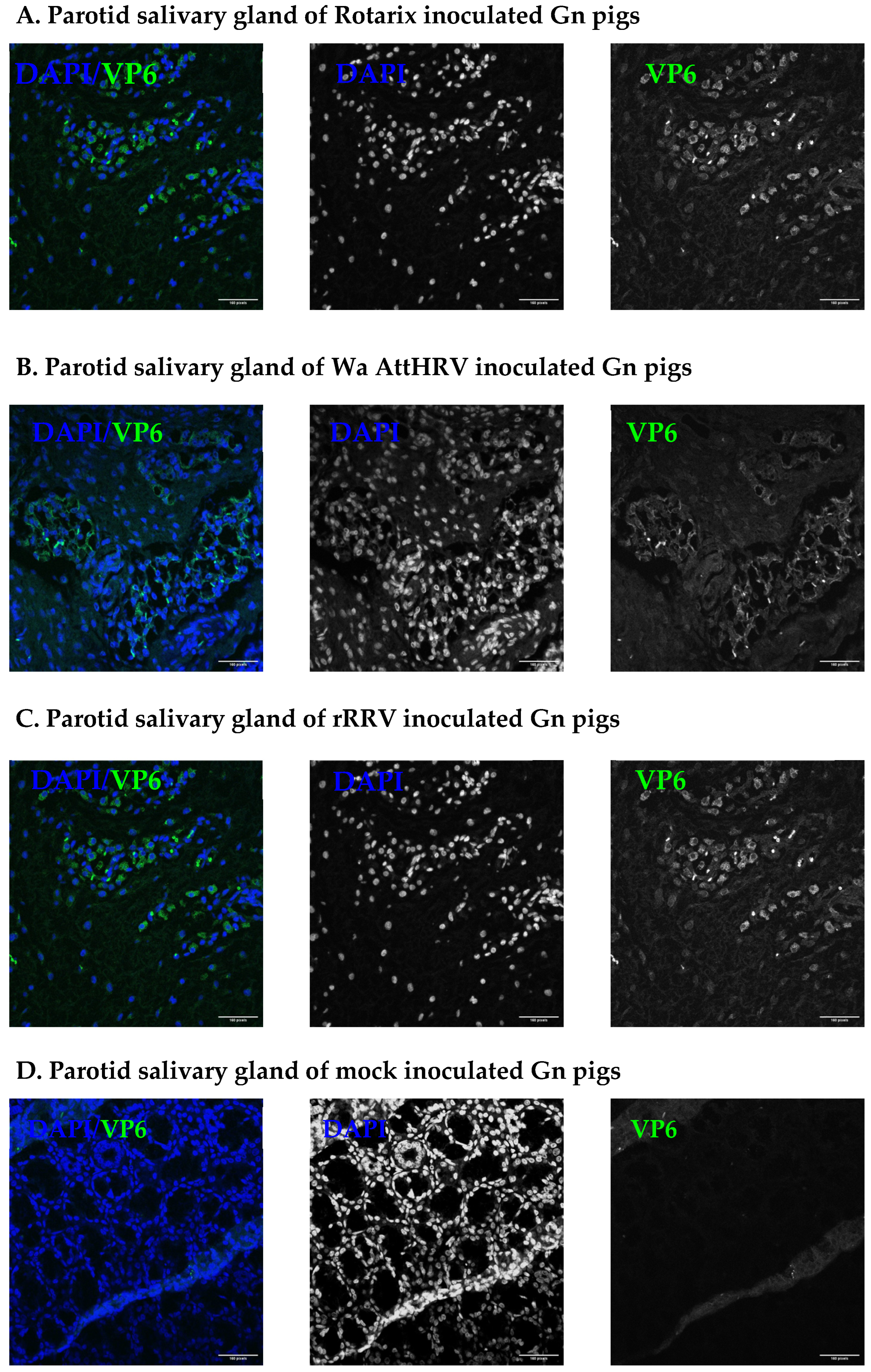
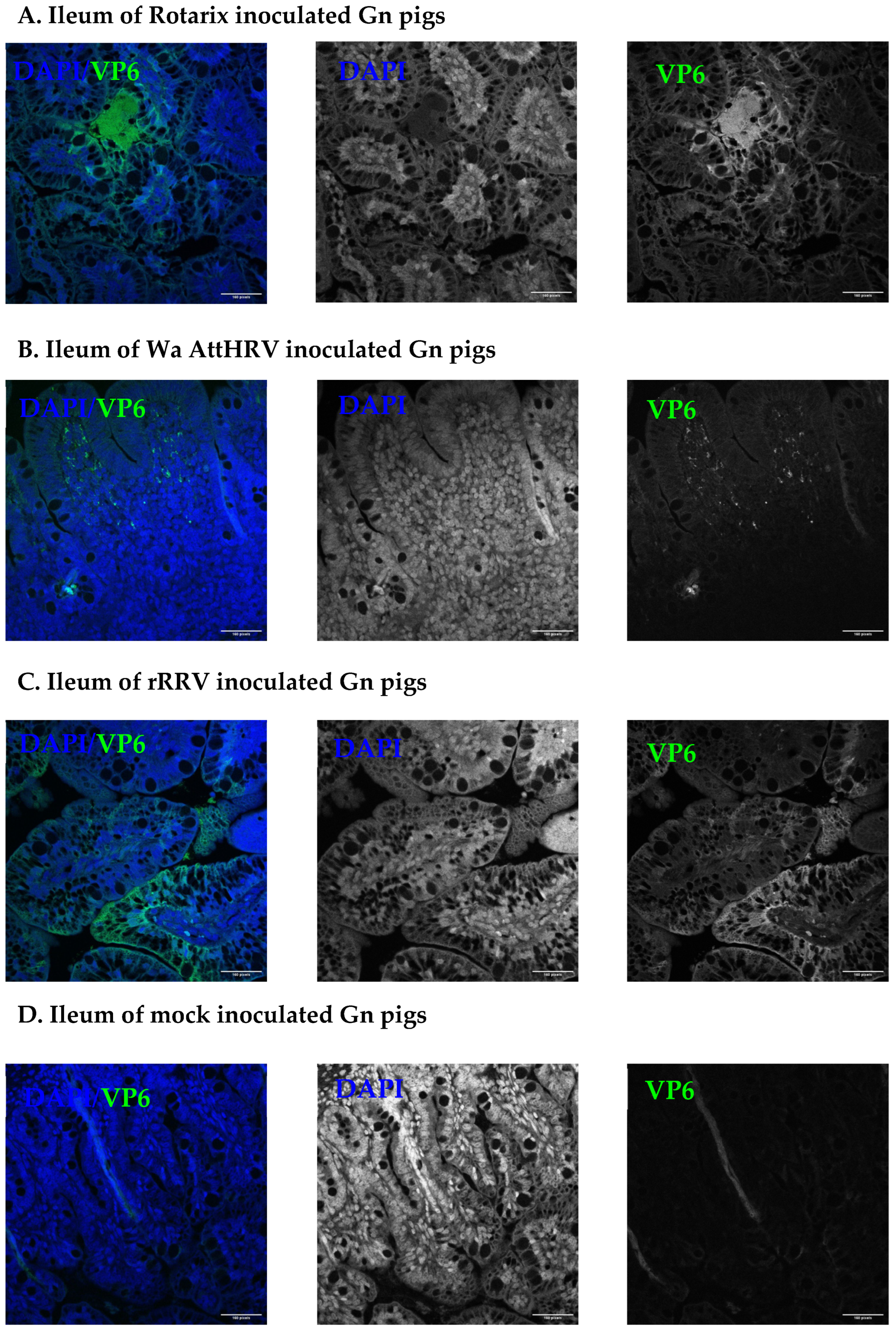
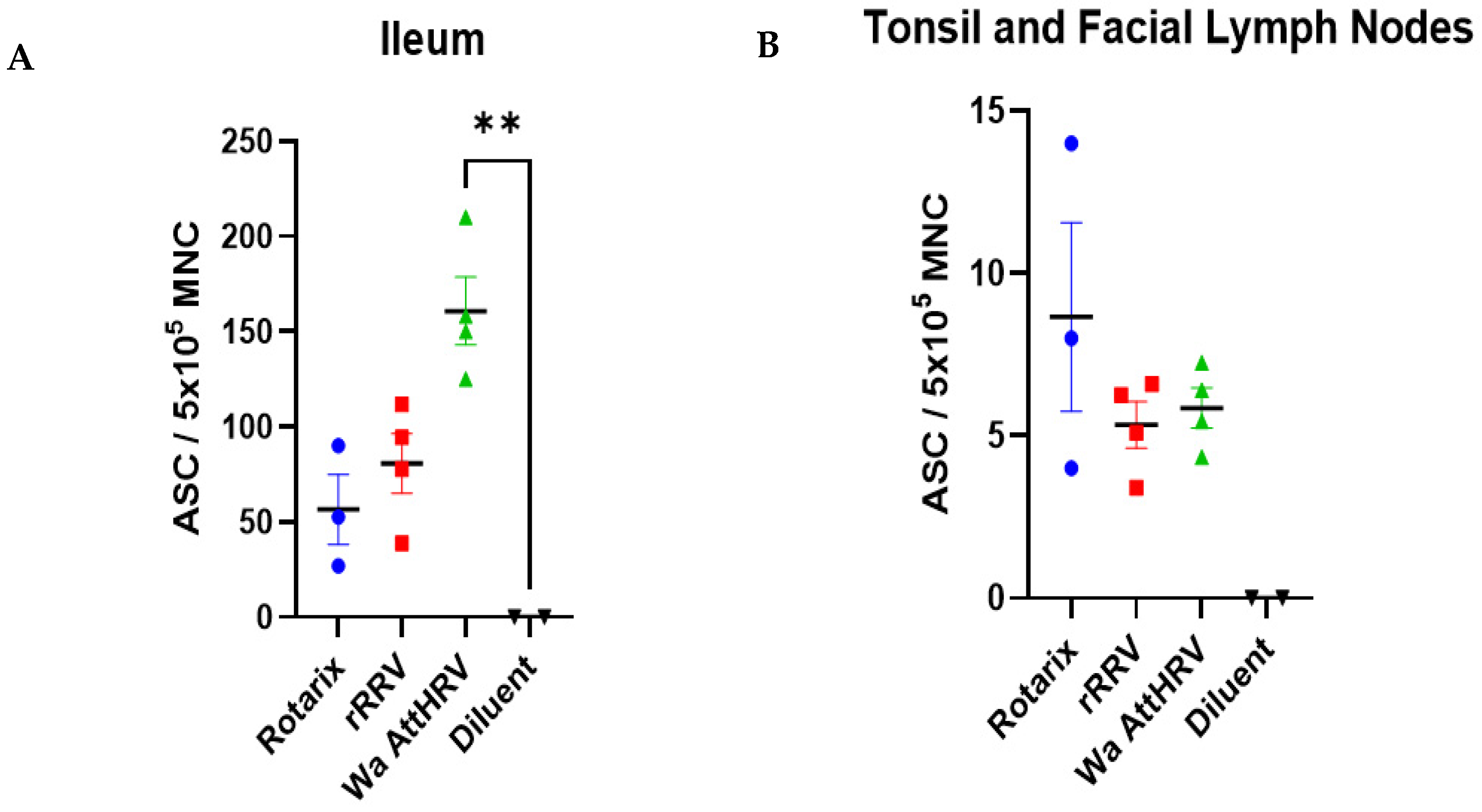
3.2. Presence of Human Rotavirus in Salivary Glands and Intestines
3.3. Infectious Human Rotavirus Particles in Saliva
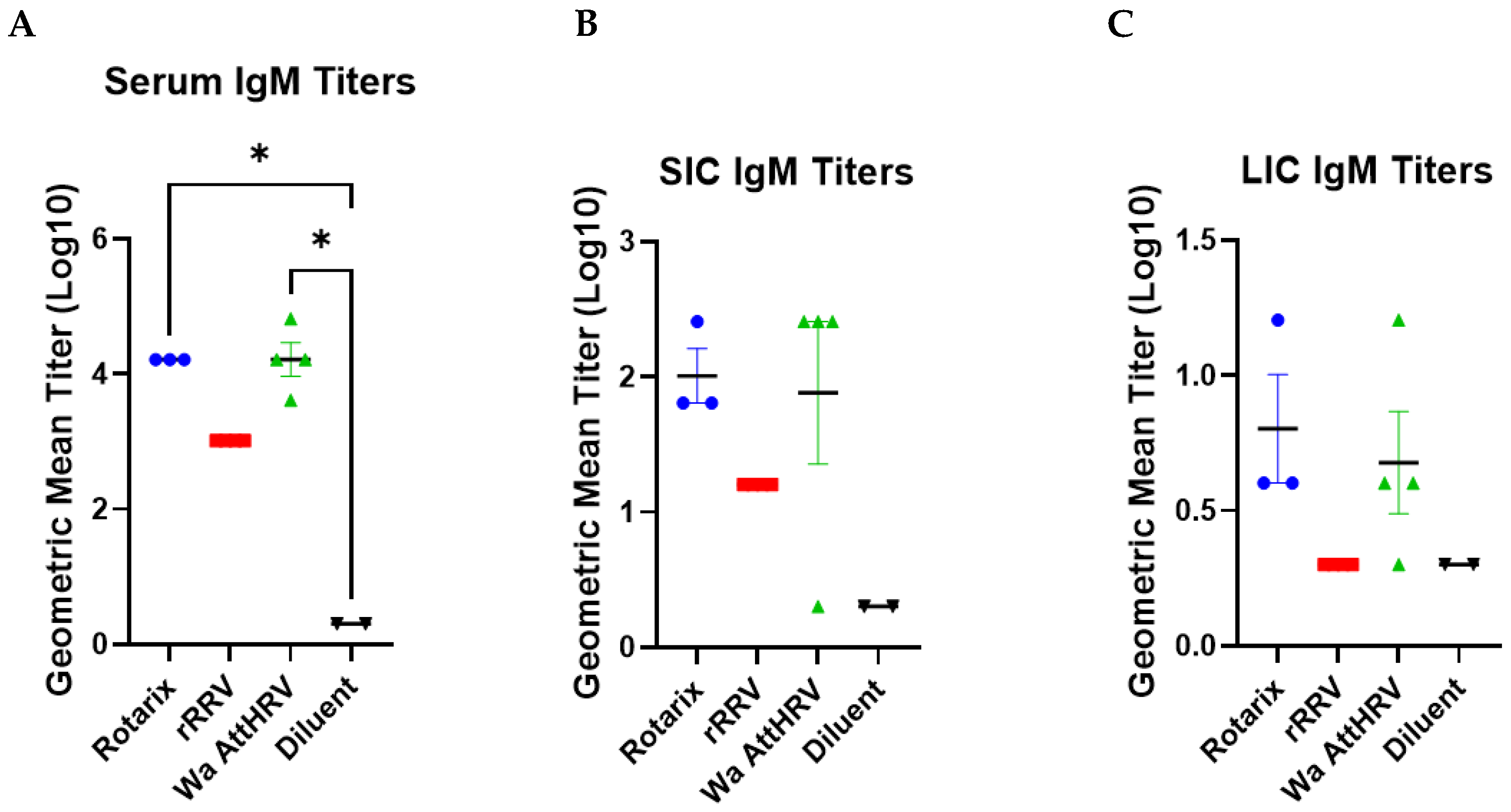
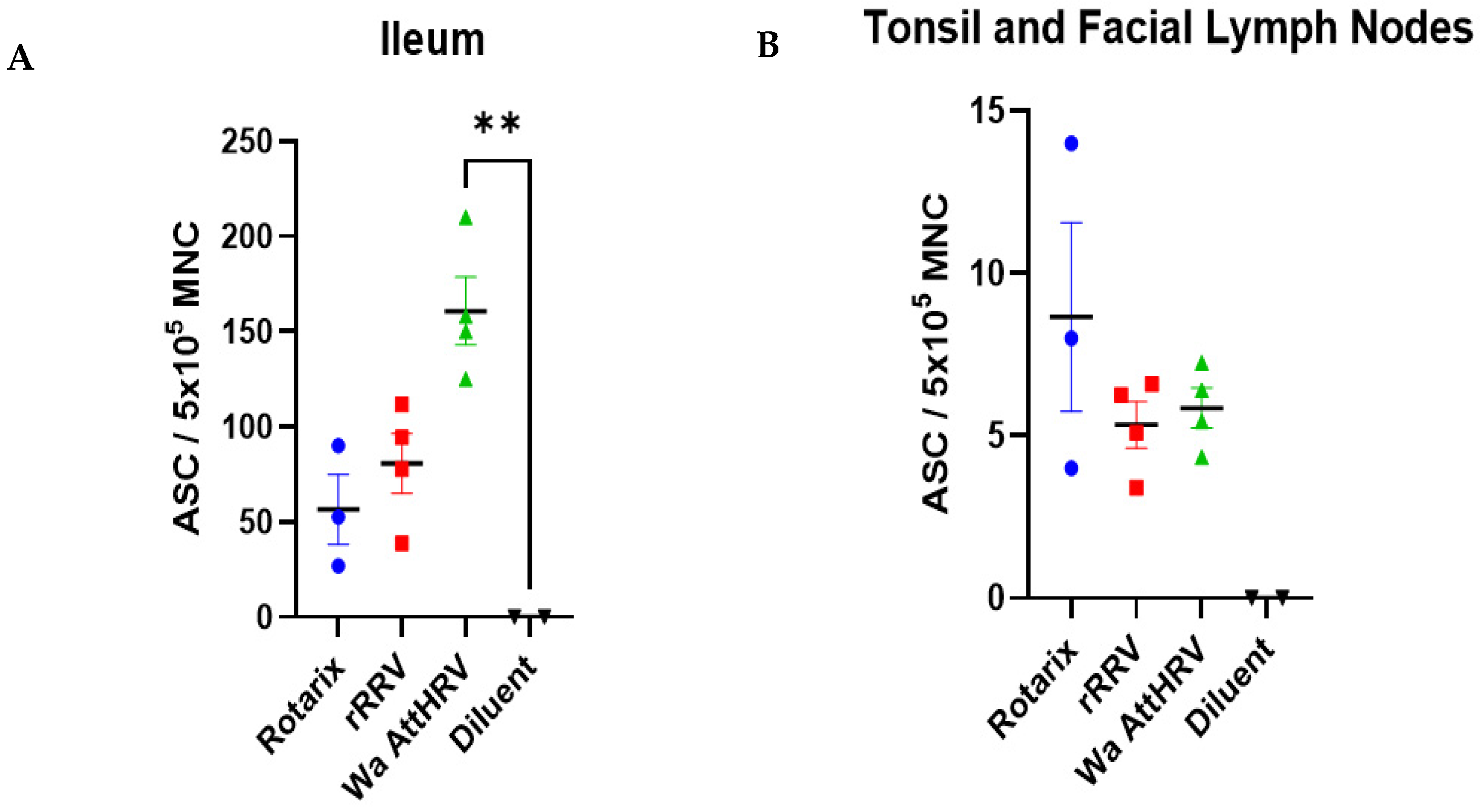
3.4. Detection of Rotavirus-Specific IgM Antibodies and IgM Antibody Secreting Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mboko, W.P.; Chhabra, P.; Valcarce, M.D.; Costantini, V.; Vinjé, J. Advances in understanding of the innate immune response to human norovirus infection using organoid models. J. Gen. Virol. 2022, 103, 1099. [Google Scholar] [CrossRef]
- Kirkwood, C.D.; Ma, L.F.; Carey, M.E.; Steele, A.D. The rotavirus vaccine development pipeline. Vaccine 2019, 37, 7328–7335. [Google Scholar] [CrossRef]
- Bányai, K.; Estes, M.K.; Martella, V.; Parashar, U.D. Viral gastroenteritis. Lancet 2018, 392, 175–186. [Google Scholar] [CrossRef]
- Ludwig-Begall, L.F.; Mauroy, A.; Thiry, E. Noroviruses-The State of the Art, Nearly Fifty Years after Their Initial Discovery. Viruses 2021, 13, 1541. [Google Scholar] [CrossRef]
- Fields, B.N. Fields Virology; Wolters Kluwer Health/Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2013; Volume 6. [Google Scholar]
- Amimo, J.O.; Raev, S.A.; Chepngeno, J.; Mainga, A.O.; Guo, Y.; Saif, L.; Vlasova, A.N. Rotavirus Interactions With Host Intestinal Epithelial Cells. Front. Immunol. 2021, 12, 793841. [Google Scholar] [CrossRef]
- Estes, M.K.; Kang, G.; Zeng, C.Q.; Crawford, S.E.; Ciarlet, M. Pathogenesis of rotavirus gastroenteritis. Novartis. Found Symp. 2001, 238, 82–96; discussion 96–100. [Google Scholar] [CrossRef]
- Ghosh, S.; Kumar, M.; Santiana, M.; Mishra, A.; Zhang, M.; Labayo, H.; Chibly, A.M.; Nakamura, H.; Tanaka, T.; Henderson, W.; et al. Enteric viruses replicate in salivary glands and infect through saliva. Nature 2022, 607, 345–350. [Google Scholar] [CrossRef]
- Gómez-Rial, J.; Curras-Tuala, M.J.; Rivero-Calle, I.; Rodríguez-Tenreiro, C.; Redondo-Collazo, L.; Gómez-Carballa, A.; Pardo-Seco, J.; Salas, A.; Martinón-Torres, F. Rotavirus intestinal infection induces an oral mucosa cytokine response. PLoS ONE 2018, 13, e0195314. [Google Scholar] [CrossRef]
- Saif, L.J.; Ward, L.A.; Yuan, L.; Rosen, B.I.; To, T.L. The gnotobiotic piglet as a model for studies of disease pathogenesis and immunity to human rotaviruses. Arch. Virol. Suppl. 1996, 12, 153–161. [Google Scholar] [CrossRef]
- Yuan, L. Vaccine Efficacy Evaluation the Gnotobiotic Pig Model; CRC Press/Taylor & Francis Group, LLC: Boca Raton, FL, USA, 2022. [Google Scholar]
- Hensley, C.; Zhou, P.; Schnur, S.; Mahsoub, H.M.; Liang, Y.; Wang, M.X.; Page, C.; Yuan, L.; Bronshtein, V. Thermostable, Dissolvable Buccal Film Rotavirus Vaccine Is Highly Effective in Neonatal Gnotobiotic Pig Challenge Model. Vaccines 2021, 9, 437. [Google Scholar] [CrossRef]
- GSK. Rotarix Suspension. 2022. Available online: https://gskpro.com/content/dam/global/hcpportal/en_US/Prescribing_Information/Rotarix/pdf/ROTARIX-PI-PIL.PDF (accessed on 5 July 2022).
- Dennehy, P.H. Rotavirus vaccines: An overview. Clin. Microbiol. Rev. 2008, 21, 198–208. [Google Scholar] [CrossRef]
- Ramesh, A.; Mao, J.; Lei, S.; Twitchell, E.; Shiraz, A.; Jiang, X.; Tan, M.; Yuan, A.L. Parenterally Administered P24-VP8* Nanoparticle Vaccine Conferred Strong Protection against Rotavirus Diarrhea and Virus Shedding in Gnotobiotic Pigs. Vaccines 2019, 7, 177. [Google Scholar] [CrossRef]
- Li, T.; Lin, H.; Yu, L.; Xue, M.; Ge, S.; Zhao, Q.; Zhang, J.; Xia, N. Development of an enzyme-linked immunospot assay for determination of rotavirus infectivity. J. Virol. Methods 2014, 209, 7–14. [Google Scholar] [CrossRef]
- Philip, A.A.; Perry, J.L.; Eaton, H.E.; Shmulevitz, M.; Hyser, J.M.; Patton, J.T. Generation of Recombinant Rotavirus Expressing NSP3-UnaG Fusion Protein by a Simplified Reverse Genetics System. J. Virol. 2019, 93, e01616–e01619. [Google Scholar] [CrossRef]
- Philip, A.A.; Dai, J.; Katen, S.P.; Patton, J.T. Simplified Reverse Genetics Method to Recover Recombinant Rotaviruses Expressing Reporter Proteins. J. Vis. Exp. 2020, 158, e61039. [Google Scholar] [CrossRef]
- Mijatovic-Rustempasic, S.; Tam, K.I.; Kerin, T.K.; Lewis, J.M.; Gautam, R.; Quaye, O.; Gentsch, J.R.; Bowen, M.D. Sensitive and specific quantitative detection of rotavirus A by one-step real-time reverse transcription-PCR assay without antecedent double-stranded-RNA denaturation. J. Clin. Microbiol. 2013, 51, 3047–3054. [Google Scholar] [CrossRef]
- Vega, C.G.; Garaicoechea, L.L.; Degiuseppe, J.I.; Bok, M.; Rivolta, A.A.; Piantanida, A.P.; Asenzo, G.; Adúriz Guerrero, M.; Wigdorovitz, A.; Stupka, J.A.; et al. ROTADIAL: The first nanobody-based immunoassay to detect Group A Rotavirus. J. Virol. Methods 2021, 298, 114279. [Google Scholar] [CrossRef]
- Garaicoechea, L.; Olichon, A.; Marcoppido, G.; Wigdorovitz, A.; Mozgovoj, M.; Saif, L.; Surrey, T.; Parreño, V. Llama-derived single-chain antibody fragments directed to rotavirus VP6 protein possess broad neutralizing activity in vitro and confer protection against diarrhea in mice. J. Virol. 2008, 82, 9753–9764. [Google Scholar] [CrossRef]
- Lei, S.; Ryu, J.; Wen, K.; Twitchell, E.; Bui, T.; Ramesh, A.; Weiss, M.; Li, G.; Samuel, H.; Clark-Deener, S.; et al. Increased and prolonged human norovirus infection in RAG2/IL2RG deficient gnotobiotic pigs with severe combined immunodeficiency. Sci. Rep. 2016, 6, 25222. [Google Scholar] [CrossRef]
- Johnston, A.; Sigurdardottir, S.L.; Ryon, J.J. Isolation of mononuclear cells from tonsillar tissue. Curr. Protoc. Immunol. 2009, 86, 7.8.1–7.8.4. [Google Scholar] [CrossRef]
- Twitchell, E.L.; Tin, C.; Wen, K.; Zhang, H.; Becker-Dreps, S.; Azcarate-Peril, M.A.; Vilchez, S.; Li, G.; Ramesh, A.; Weiss, M.; et al. Modeling human enteric dysbiosis and rotavirus immunity in gnotobiotic pigs. Gut. Pathog. 2016, 8, 51. [Google Scholar] [CrossRef]
- Azevedo, M.S.; Yuan, L.; Jeong, K.I.; Gonzalez, A.; Nguyen, T.V.; Pouly, S.; Gochnauer, M.; Zhang, W.; Azevedo, A.; Saif, L.J. Viremia and nasal and rectal shedding of rotavirus in gnotobiotic pigs inoculated with Wa human rotavirus. J. Virol. 2005, 79, 5428–5436. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Hagbom, M.; Svensson, L.; Nordgren, J. The Impact of Human Genetic Polymorphisms on Rotavirus Susceptibility, Epidemiology, and Vaccine Take. Viruses 2020, 12, 324. [Google Scholar] [CrossRef]
- Philip, A.A.; Patton, J.T. Generation of Recombinant Rotaviruses Expressing Human Norovirus Capsid Proteins. J. Virol. 2022, 96, e0126222. [Google Scholar] [CrossRef]
- Kawagishi, T.; Sánchez-Tacuba, L.; Feng, N.; Costantini, V.P.; Tan, M.; Jiang, X.; Green, K.Y.; Vinjé, J.; Ding, S.; Greenberg, H.B. Mucosal and systemic neutralizing antibodies to norovirus induced in infant mice orally inoculated with recombinant rotaviruses. Proc. Natl. Acad. Sci. USA 2023, 120, e2214421120. [Google Scholar] [CrossRef]
- Gómez-Rial, J.; Curras-Tuala, M.J.; Talavero-González, C.; Rodríguez-Tenreiro, C.; Vilanova-Trillo, L.; Gómez-Carballa, A.; Rivero-Calle, I.; Justicia-Grande, A.; Pardo-Seco, J.; Redondo-Collazo, L.; et al. Salivary epidermal growth factor correlates with hospitalization length in rotavirus infection. BMC Infect. Dis. 2017, 17, 370. [Google Scholar] [CrossRef]
- Laban, N.M.; Goodier, M.R.; Bosomprah, S.; Simuyandi, M.; Chisenga, C.; Chilyabanyama, O.N.; Chilengi, R. T-Cell Responses after Rotavirus Infection or Vaccination in Children: A Systematic Review. Viruses 2022, 14, 459. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nasal | |||||
|---|---|---|---|---|---|
| Group | %Shedding Virus | Mean Days to Onset | Mean Duration | Mean Peak Titer | AUC Virus Shedding |
| Rotarix | 4/5 (80%) A | 1.0 (0) A | 4.67 (0.33) A | 1.81 × 107 (9.5 × 106) | 1.32 × 107 (7.41 × 106) A |
| rRRV | 3/6 (50%) AB | 1.5 (0.5) A | 1.5 (0.87) B | 3.0 × 106 (2.0 × 106) | 2.4 × 106 (2.34 × 106) B |
| Wa AttHRV | 4/6 (66.7%) AB | 3.0 (1.23) A | 3.5 (0.65) AB | 4.61 × 106 (1.69 × 106) | 3.78 × 106 (1.63 × 106) AB |
| Diluent | 0/1 B | NA | 0 | 0 | 0 |
| Fecal | |||||
| Rotarix | 4/5 (80%) A | 1.0 (0) B | 3.67 (0.88) A | 2.63 × 107 (2.41 × 107) | 1.77 × 107 (1.56 × 107) AB |
| rRRV | 3/6 (50%) AB | 1.5 (0.5) A | 1.0 (0.41) B | 1.67 × 106 (8.37 × 105) | 1.19 × 106 (8.39 × 105) B |
| Wa AttHRV | 4/6 (66.7%) AB | 3.0 (1.23) B | 3.75 (0.48) A | 6.44 × 107 (1.64 × 107) | 5.89 × 107 (2.88 × 107) A |
| Diluent | 0/1 B | NA | 0 | 0 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nyblade, C.; Zhou, P.; Frazier, M.; Frazier, A.; Hensley, C.; Fantasia-Davis, A.; Shahrudin, S.; Hoffer, M.; Agbemabiese, C.A.; LaRue, L.; et al. Human Rotavirus Replicates in Salivary Glands and Primes Immune Responses in Facial and Intestinal Lymphoid Tissues of Gnotobiotic Pigs. Viruses 2023, 15, 1864. https://doi.org/10.3390/v15091864
Nyblade C, Zhou P, Frazier M, Frazier A, Hensley C, Fantasia-Davis A, Shahrudin S, Hoffer M, Agbemabiese CA, LaRue L, et al. Human Rotavirus Replicates in Salivary Glands and Primes Immune Responses in Facial and Intestinal Lymphoid Tissues of Gnotobiotic Pigs. Viruses. 2023; 15(9):1864. https://doi.org/10.3390/v15091864
Chicago/Turabian StyleNyblade, Charlotte, Peng Zhou, Maggie Frazier, Annie Frazier, Casey Hensley, Ariana Fantasia-Davis, Shabihah Shahrudin, Miranda Hoffer, Chantal Ama Agbemabiese, Lauren LaRue, and et al. 2023. "Human Rotavirus Replicates in Salivary Glands and Primes Immune Responses in Facial and Intestinal Lymphoid Tissues of Gnotobiotic Pigs" Viruses 15, no. 9: 1864. https://doi.org/10.3390/v15091864
APA StyleNyblade, C., Zhou, P., Frazier, M., Frazier, A., Hensley, C., Fantasia-Davis, A., Shahrudin, S., Hoffer, M., Agbemabiese, C. A., LaRue, L., Barro, M., Patton, J. T., Parreño, V., & Yuan, L. (2023). Human Rotavirus Replicates in Salivary Glands and Primes Immune Responses in Facial and Intestinal Lymphoid Tissues of Gnotobiotic Pigs. Viruses, 15(9), 1864. https://doi.org/10.3390/v15091864