


Dynamics of Infection of Atypical Porcine Pestivirus in Commercial Pigs from Birth to Market: A Longitudinal Study

 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Care and Use
2.2. Longitudinal Field Study
2.3. APPV Nucleic Acid Quantification
2.4. Indirect Enzyme-Linked Immunosorbent Assays (ELISAs)
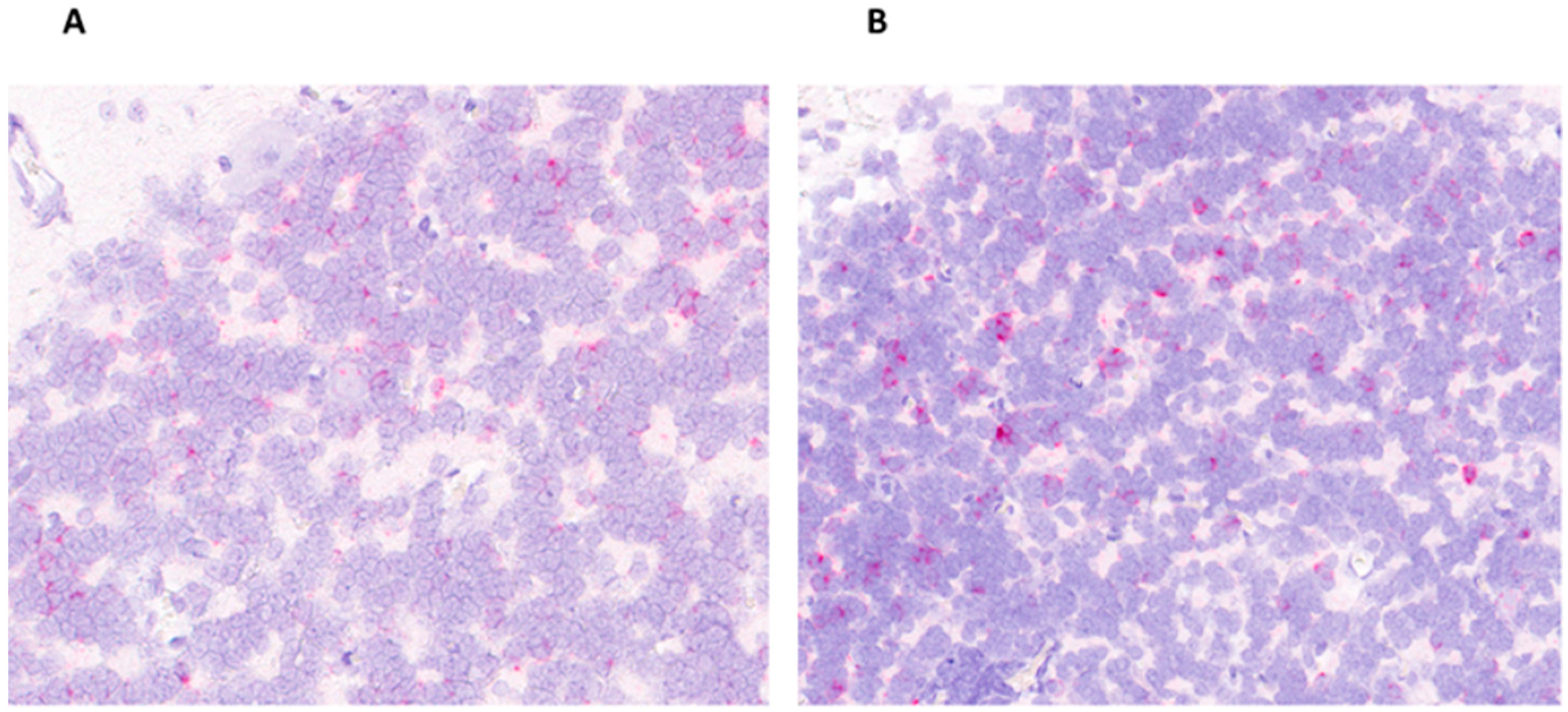
2.5. In Situ Hybridization
2.6. Data Analysis
3. Results
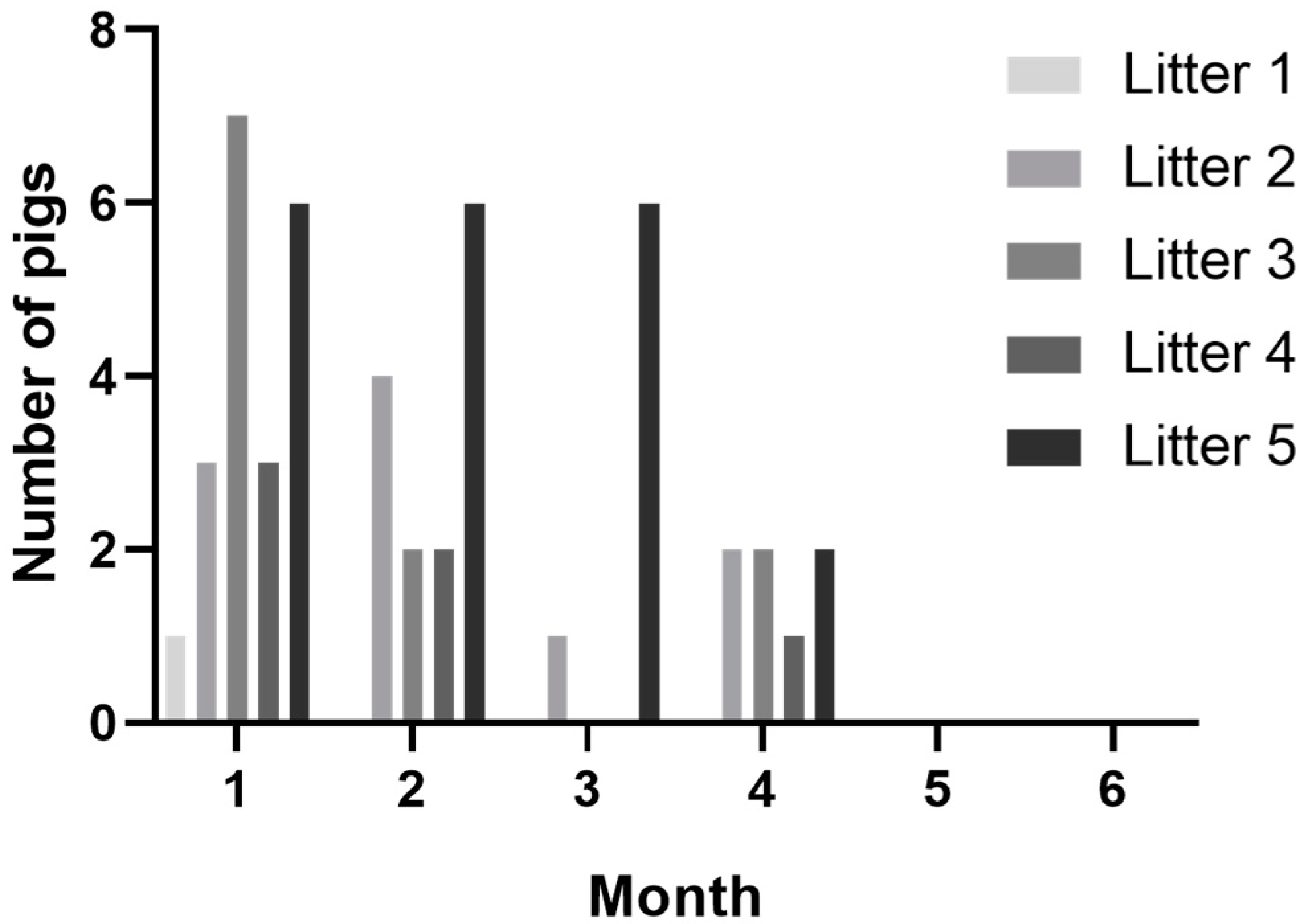
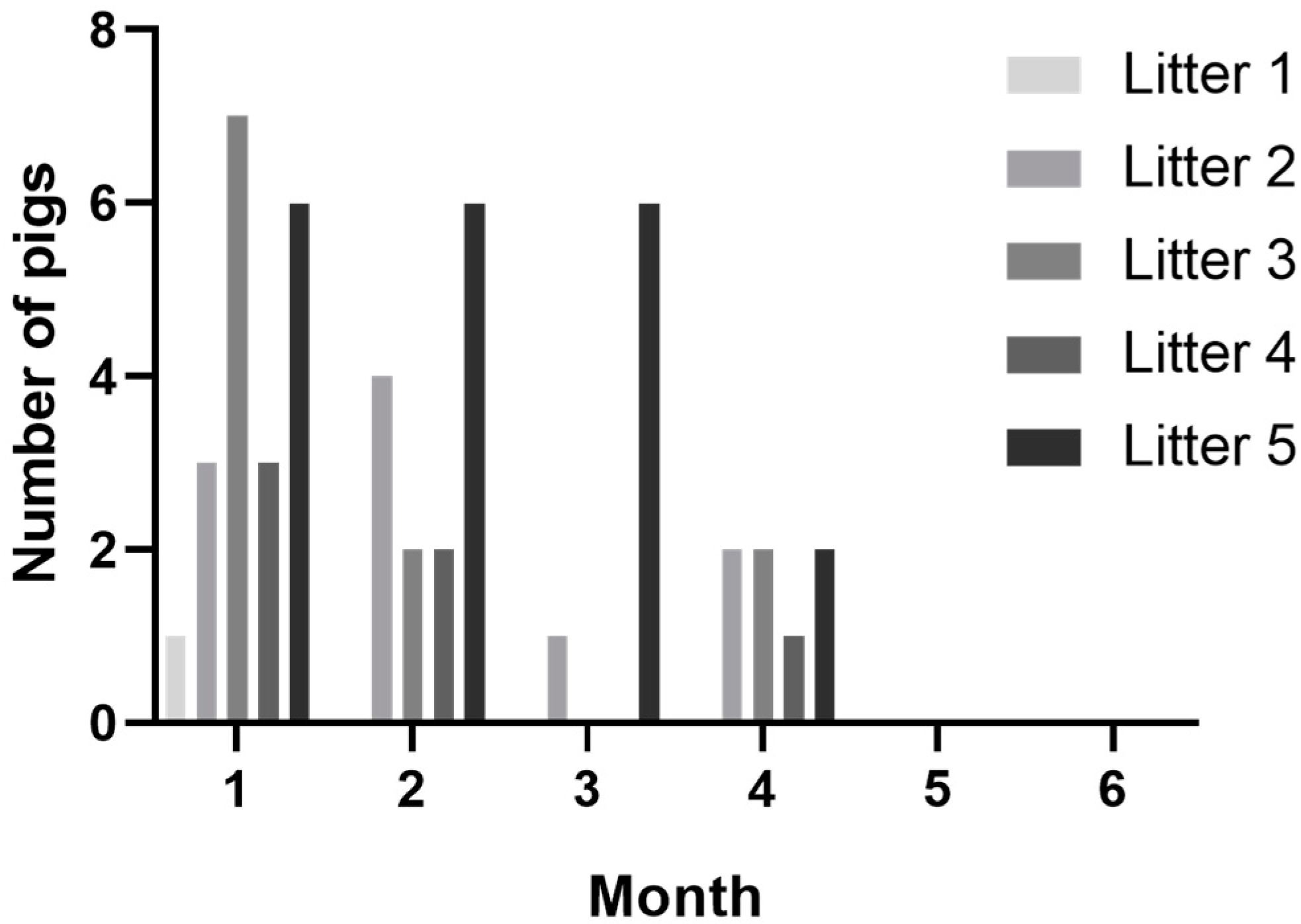
3.1. Congenital Tremors
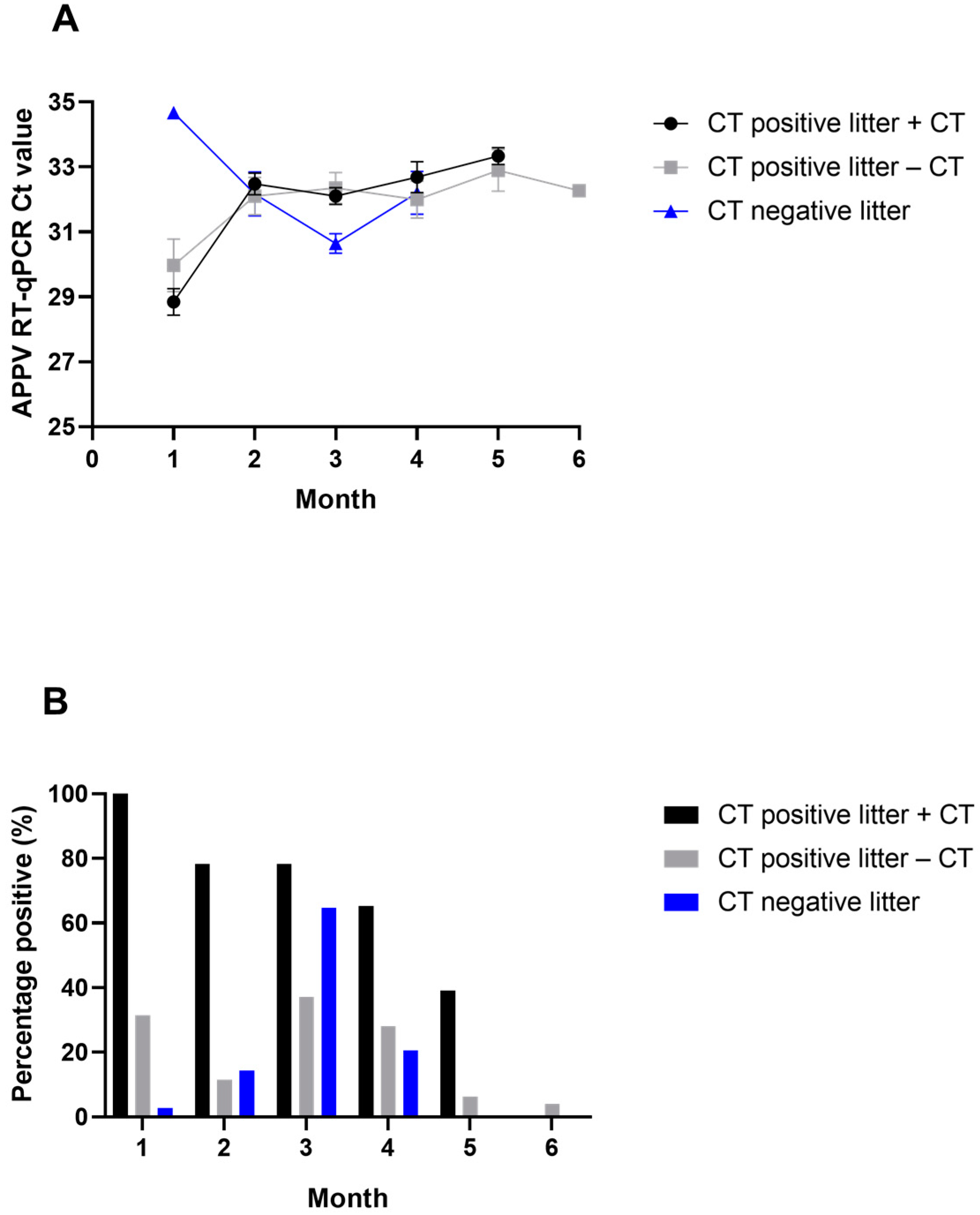
3.2. APPV RNA Detection by RT-qPCR
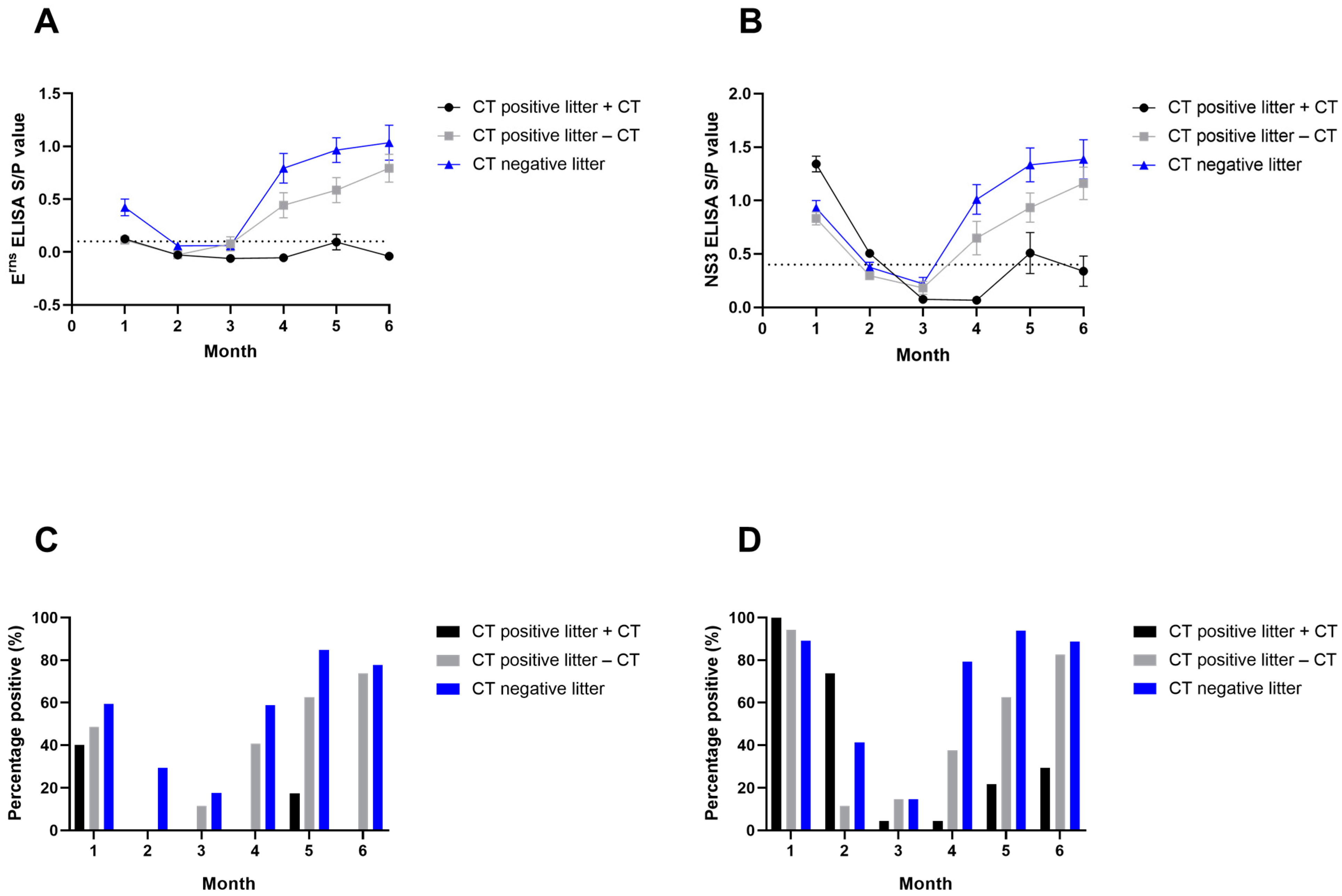
3.3. APPV Antibody Detection by ELISA
3.4. APPV RNA across Tissues
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Groof, A.; Deijs, M.; Guelen, L.; Van Grinsven, L.; Van Os-Galdos, L.; Vogels, W.; Derks, C.; Cruijsen, T.; Geurts, V.; Vrijenhoek, M.; et al. Atypical Porcine Pestivirus: A Possible Cause of Congenital Tremor Type A-II in Newborn Piglets. Viruses 2016, 8, 271. [Google Scholar] [CrossRef] [PubMed]
- Stenberg, H.; Jacobson, M.; Malmberg, M. A review of congenital tremor type A-II in piglets. Anim. Health Res. Rev. 2020, 21, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Pan, S.; Mou, C.; Chen, Z. An emerging novel virus: Atypical porcine pestivirus (APPV). Rev. Med. Virol. 2018, 29, e2018. [Google Scholar] [CrossRef] [PubMed]
- Arruda, B.L.; Arruda, P.H.; Magstadt, D.R.; Schwartz, K.J.; Dohlman, T.; Schleining, J.A.; Patterson, A.R.; Visek, C.A.; Victoria, J.G. Identification of a Divergent Lineage Porcine Pestivirus in Nursing Piglets with Congenital Tremors and Reproduction of Disease following Experimental Inoculation. PLoS ONE 2016, 11, e0150104. [Google Scholar] [CrossRef]
- Hause, B.M.; Collin, E.A.; Peddireddi, L.; Yuan, F.; Chen, Z.; Hesse, R.A.; Gauger, P.C.; Clement, T.; Fang, Y.; Anderson, G. Discovery of a novel putative atypical porcine pestivirus in pigs in the USA. J. Gen. Virol. 2015, 96, 2994–2998. [Google Scholar] [CrossRef]
- Zhang, K.; Wu, K.; Liu, J.; Ge, S.; Xiao, Y.; Shang, Y.; Ning, Z. Identification of atypical porcine pestivirus infection in swine herds in China. Transbound. Emerg. Dis. 2017, 64, 1020–1023. [Google Scholar] [CrossRef]
- Gatto, I.R.H.; Harmon, K.; Bradner, L.; Silva, P.; Linhares, D.C.L.; Arruda, P.H.; de Oliveira, L.G.; Arruda, B.L. Detection of atypical porcine pestivirus in Brazil in the central nervous system of suckling piglets with congenital tremor. Transbound. Emerg. Dis. 2018, 65, 375–380. [Google Scholar] [CrossRef]
- Beer, M.; Wernike, K.; Dräger, C.; Höper, D.; Pohlmann, A.; Bergermann, C.; Schröder, C.; Klinkhammer, S.; Blome, S.; Hoffmann, B. High Prevalence of Highly Variable Atypical Porcine Pestiviruses Found in Germany. Transbound. Emerg. Dis. 2017, 64, e22–e26. [Google Scholar] [CrossRef]
- Postel, A.; Meyer, D.; Cagatay, G.N.; Feliziani, F.; De Mia, G.M.; Fischer, N.; Grundhoff, A.; Milićević, V.; Deng, M.-C.; Chang, C.-Y.; et al. High Abundance and Genetic Variability of Atypical Porcine Pestivirus in Pigs from Europe and Asia. Emerg. Infect. Dis. 2017, 23, 2104–2107. [Google Scholar] [CrossRef]
- Postel, A.; Meyer, D.; Petrov, A.; Becher, P. Recent emergence of a novel porcine pestivirus: Interference with classical swine fever diagnosis? Emerg. Microbes Infect. 2017, 6, e19. [Google Scholar] [CrossRef]
- Cagatay, G.N.; Meyer, D.; Wendt, M.; Becher, P.; Postel, A. Characterization of the Humoral Immune Response Induced after In-fection with Atypical Porcine Pestivirus (APPV). Viruses 2019, 11, 880. [Google Scholar] [CrossRef] [PubMed]
- Arruda, B.L.; Falkenberg, S.; Mora-Díaz, J.-C.; Ferreyra, F.S.M.; Magtoto, R.; Giménez-Lirola, L. Development and Evaluation of Antigen-Specific Dual Matrix Pestivirus K ELISAs Using Longitudinal Known Infectious Status Samples. J. Clin. Microbiol. 2022, 60, e0069722. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Yu, Y.; Song, C.; Cao, Z. Genetic characterization of atypical porcine pestivirus (APPV) in China and the successful isolation of a novel APPV strain within genotype 2. Microb. Pathog. 2021, 161, 105282. [Google Scholar] [CrossRef]
- Schwarz, L.; Riedel, C.; Högler, S.; Sinn, L.J.; Voglmayr, T.; Wöchtl, B.; Dinhopl, N.; Rebel-Bauder, B.; Weissenböck, H.; Ladinig, A.; et al. Congenital infection with atypical porcine pestivirus (APPV) is associated with disease and viral persistence. Vet. Res. 2017, 48, 1. [Google Scholar] [CrossRef]
- Folgueiras-González, A.; Braak, R.V.D.; Simmelink, B.; Deijs, M.; Van Der Hoek, L.; De Groof, A. Atypical Porcine Pestivirus Circulation and Molecular Evolution within an Affected Swine Herd. Viruses 2020, 12, 1080. [Google Scholar] [CrossRef]
- Buckley, A.C.; Falkenberg, S.M.; Palmer, M.V.; Arruda, P.H.; Magstadt, D.R.; Schwartz, K.J.; Gatto, I.R.; Neill, J.D.; Arruda, B.L. Distribution and persistence of atypical porcine pestivirus in experimentally inoculated pigs. J. Vet. Diagn. Investig. 2021, 33, 952–955. [Google Scholar] [CrossRef] [PubMed]
- Finlaison, D.S.; Kirkland, P.D. The Outcome of Porcine Foetal Infection with Bungowannah Virus is Dependent on the Stage of Gestation at Which Infection Occurs. Part 1: Serology and Virology. Viruses 2020, 12, 691. [Google Scholar] [CrossRef]
- Muñoz-González, S.; Ruggli, N.; Rosell, R.; Pérez, L.J.; Frías-Leuporeau, M.T.; Fraile, L.; Montoya, M.; Cordoba, L.; Domingo, M.; Ehrensperger, F.; et al. Postnatal Persistent Infection with Classical Swine Fever Virus and Its Immunological Implications. PLoS ONE 2015, 10, e0125692. [Google Scholar] [CrossRef]
- Schweizer, M.; Peterhans, E. Pestiviruses. Annu. Rev. Anim. Biosci. 2014, 2, 141–163. [Google Scholar] [CrossRef]
- Liu, J.; Li, Z.; Ren, X.; Li, H.; Lu, R.; Zhang, Y.; Ning, Z. Viral load and histological distribution of atypical porcine pestivirus in different tissues of naturally infected piglets. Arch. Virol. 2019, 164, 2519–2523. [Google Scholar] [CrossRef]
- Yuan, J.; Han, Z.; Li, J.; Huang, Y.; Yang, J.; Ding, H.; Zhang, J.; Zhu, M.; Zhang, Y.; Liao, J.; et al. Atypical Porcine Pestivirus as a Novel Type of Pestivirus in Pigs in China. Front. Microbiol. 2017, 8, 862. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Qian, P.; Hu, Z.; Chen, H.; Li, X. Genetic characterization of atypical porcine pestivirus from neonatal piglets with congenital tremor in Hubei province, China. Virol. J. 2022, 19, 51. [Google Scholar] [CrossRef] [PubMed]
- Postel, A.; Hansmann, F.; Baechlein, C.; Fischer, N.; Alawi, M.; Grundhoff, A.; Derking, S.; Tenhündfeld, J.; Pfankuche, V.M.; Herder, V.; et al. Presence of atypical porcine pestivirus (APPV) genomes in newborn piglets correlates with congenital tremor. Sci. Rep. 2016, 6, 27735. [Google Scholar] [CrossRef] [PubMed]
- Sutton, K.M.; Lahmers, K.K.; Harris, S.P.; Wijesena, H.R.; Mote, B.E.; Kachman, S.D.; Borza, T.; Ciobanu, D.C. Detection of atypical porcine pestivirus genome in newborn piglets affected by congenital tremor and high preweaning mortality1. J. Anim. Sci. 2019, 97, 4093–4100. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-González, S.; Canturri, A.; Pérez-Simó, M.; Bohórquez, J.A.; Rosell, R.; Cabezón, O.; Segalés, J.; Domingo, M.; Ganges, L. First report of the novel atypical porcine pestivirus in Spain and a retrospective study. Transbound. Emerg. Dis. 2017, 64, 1645–1649. [Google Scholar] [CrossRef]
- Dessureault, F.G.; Choinière, M.; Provost, C.; Gagnon, C.A. First report of atypical porcine pestivirus in piglets with congenital tremor in Canada. Can. Vet. J. 2018, 59, 429–432. [Google Scholar]
- Ganges, L.; Crooke, H.R.; Bohórquez, J.A.; Postel, A.; Sakoda, Y.; Becher, P.; Ruggli, N. Classical swine fever virus: The past, present and future. Virus Res. 2020, 289, 198151. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dam # | Parity | Pig Age When Enrolled (Days) | Litter Size | CT-Positive Pigs (%) | # Males (# CT-Positive Males) | # Females (# CT-Positive Females) |
|---|---|---|---|---|---|---|
| 1 | 1 | 13 | 12 | 1 (8%) | 7 (0) | 5 (1) |
| 2 | 1 | 13 | 11 * | 4 (36%) | 4 (1) | 6 (2) |
| 3 | 1 | 13 | 12 | 8 (67%) | 5 (3) | 7 (5) |
| 4 | 1 | 13 | 13 | 3 (23%) | 10 (3) | 3 (0) |
| 5 | 3 | 3 | 12 * | 9 (75%) | 6 (4) | 5 (4) |
| 6 | 1 | 17 | 11 | 0 (0%) | 7 (0) | 4 (0) |
| 7 | 1 | 16 | 13 * | 0 (0%) | 3 (0) | 9 (0) |
| 8 | 3 | 4 | 13 # | 0 (0%) | 6 (0) | 5 (0) |
| Pig # (Sex) | CT Status of Litter | Right Cerebellum | Left Cerebellum | Nasal Turbinate | Mandibular Lymph Node | Mandibular Salivary Gland | Tonsil | Palatoglossal Arch |
|---|---|---|---|---|---|---|---|---|
| 50 (F) | Positive | 21.8 | 20.9 | 27.0 | ND | 26.6 | 32.3 | 29.5 |
| 52 (M) | Positive | 21.9 | 22.3 | 29.1 | ND | 32.5 | ND | 29.8 |
| 53 (M) | Positive | 24.9 | 24.0 | 26.0 | ND | ND | ND | ND |
| 55 (M) | Positive | 19.4 | 20.2 | 22.5 | 32.9 | 26.1 | 29.8 | 32.3 |
| 61 (M) | Negative | 21.5 | 21.6 | 24.7 | ND | 25.5 | 28.3 | 28.1 |
| 62 (F) | Negative | 24.0 | 34.0 | 23.7 | ND | 33.5 | 33.9 | 33.4 |
| 70 (F) | Negative | 30.9 | 24.3 | 21.2 | ND | 29.8 | 31.8 | 32.0 |
| 91 (M) | Negative | 34.9 | 20.0 | 22.5 | ND | ND | ND | ND |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buckley, A.C.; Mora-Díaz, J.-C.; Magtoto, R.L.; Hulzen, A.V.; Ferreyra, F.M.; Falkenberg, S.M.; Giménez-Lirola, L.G.; Arruda, B.L. Dynamics of Infection of Atypical Porcine Pestivirus in Commercial Pigs from Birth to Market: A Longitudinal Study. Viruses 2023, 15, 1767. https://doi.org/10.3390/v15081767
Buckley AC, Mora-Díaz J-C, Magtoto RL, Hulzen AV, Ferreyra FM, Falkenberg SM, Giménez-Lirola LG, Arruda BL. Dynamics of Infection of Atypical Porcine Pestivirus in Commercial Pigs from Birth to Market: A Longitudinal Study. Viruses. 2023; 15(8):1767. https://doi.org/10.3390/v15081767
Chicago/Turabian StyleBuckley, Alexandra C., Juan-Carlos Mora-Díaz, Ronaldo L. Magtoto, Amberly Van Hulzen, Franco Matias Ferreyra, Shollie M. Falkenberg, Luis G. Giménez-Lirola, and Bailey L. Arruda. 2023. "Dynamics of Infection of Atypical Porcine Pestivirus in Commercial Pigs from Birth to Market: A Longitudinal Study" Viruses 15, no. 8: 1767. https://doi.org/10.3390/v15081767
APA StyleBuckley, A. C., Mora-Díaz, J.-C., Magtoto, R. L., Hulzen, A. V., Ferreyra, F. M., Falkenberg, S. M., Giménez-Lirola, L. G., & Arruda, B. L. (2023). Dynamics of Infection of Atypical Porcine Pestivirus in Commercial Pigs from Birth to Market: A Longitudinal Study. Viruses, 15(8), 1767. https://doi.org/10.3390/v15081767