
BNT162b2 Elicited an Efficient Cell-Mediated Response against SARS-CoV-2 in Kidney Transplant Recipients and Common Variable Immunodeficiency Patients

 , , , ,
, , , ,  ,
,  ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Samples Collection
2.3. Determination of Released Interferon-γ in Plasma
2.4. Humoral Antibody Response
2.5. SARS-CoV-2 Neutralization Assay (MNA)
2.6. ELISA Assay
2.7. Statistical Analysis
3. Results
3.1. Serum Levels of IgG Directed against the (Trimeric) SARS-CoV-2 Spike-Proteinin CVID and KTR Patients after the Third Dose of mRNA Vaccine
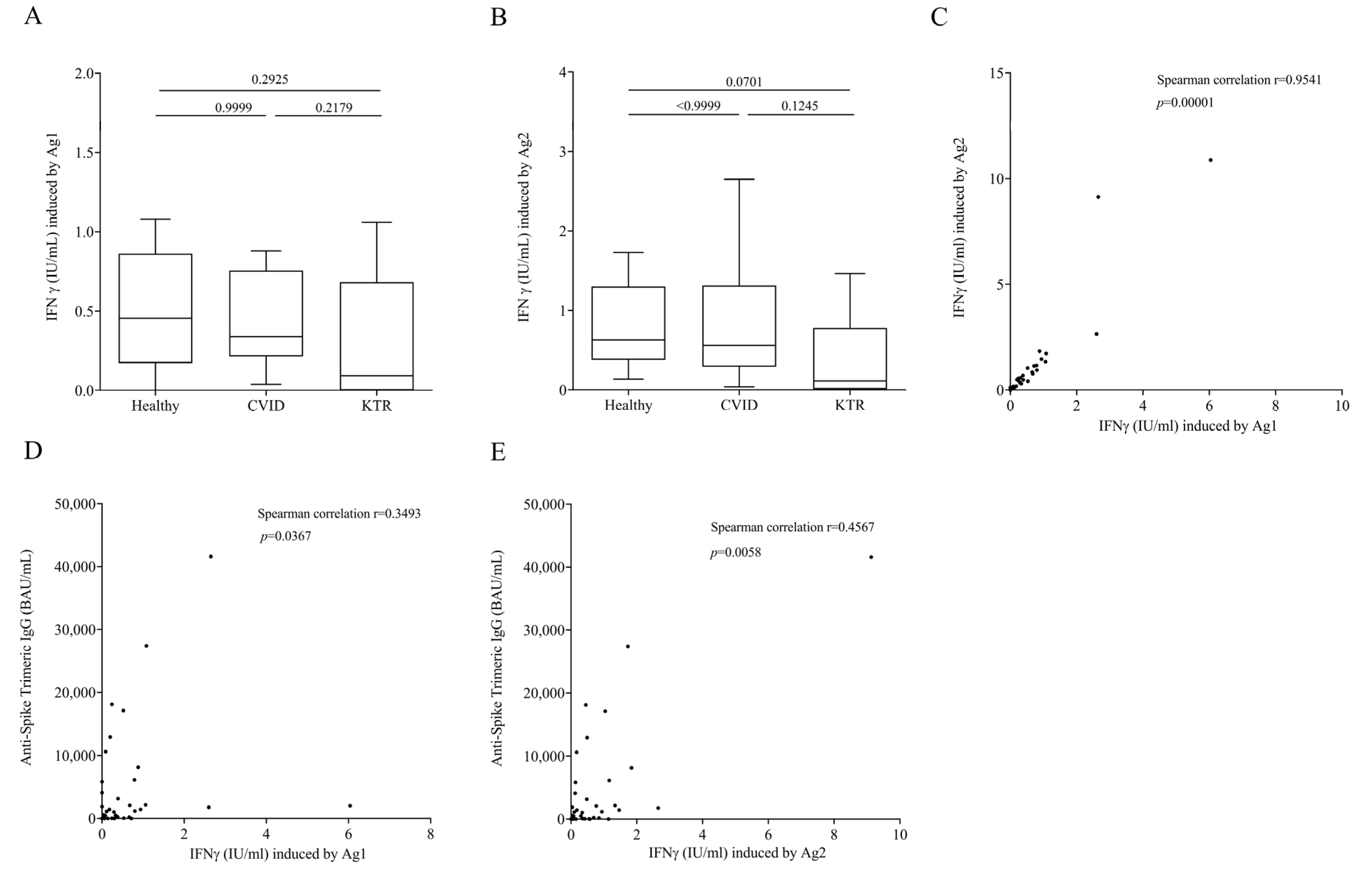
3.2. QFN SARS-CoV-2 Assay in Vaccinated, but Not Infected Subjects
3.3. Cytokine and Chemokine Concentration upon Ag1 and Ag2 Stimulation in Plasma of Vaccinated, but Not Infected Subjects
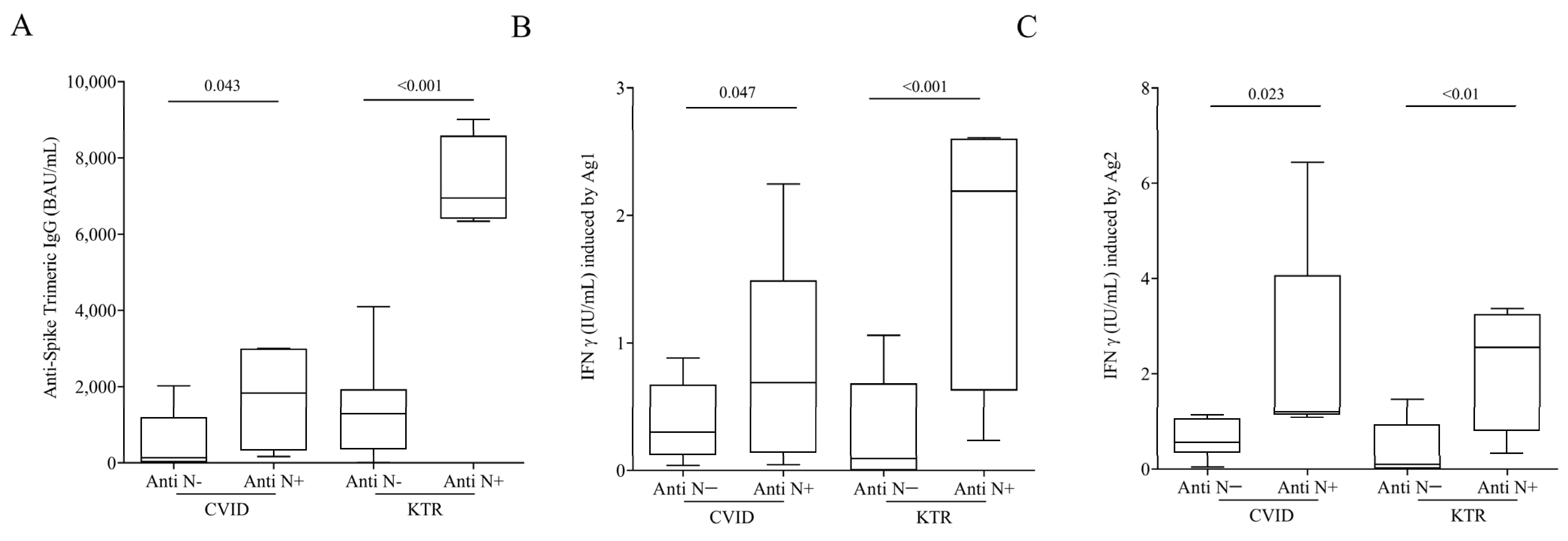
3.4. Humoral and Cell-Mediated Response to SARS-CoV-2 in Hybrid Immunized CVID and KTR Patients
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pereira, M.R.; Mohan, S.; Cohen, D.J.; Husain, S.A.; Dube, G.K.; Ratner, L.E.; Arcasoy, S.; Aversa, M.M.; Benvenuto, L.J.; Dadhania, D.M.; et al. COVID-19 in solid organ transplant recipients: Initial report from the US epicenter. Am. J. Transpl. 2020, 20, 1800–1808. [Google Scholar] [CrossRef]
- Babaha, F.; Rezaei, N. Primary Immunodeficiency Diseases in COVID-19 Pandemic: A Predisposing or Protective Factor? Am. J. Med. Sci. 2020, 360, 740–741. [Google Scholar] [CrossRef]
- Caillard, S.; Thaunat, O. COVID-19 vaccination in kidney transplant recipients. Nat. Rev. Nephrol. 2021, 17, 785–787. [Google Scholar] [CrossRef]
- Chapel, H.; Lucas, M.; Lee, M.; Bjorkander, J.; Webster, D.; Grimbacher, B.; Fieschi, C.; Thon, V.; Abedi, M.R.; Hammarstrom, L. Common variable immunodeficiency disorders: Division into distinct clinical phenotypes. Blood 2008, 112, 277–286. [Google Scholar] [CrossRef]
- Quinti, I.; Soresina, A.; Spadaro, G.; Martino, S.; Donnanno, S.; Agostini, C.; Claudio, P.; Franco, D.; Maria Pesce, A.; Borghese, F.; et al. Long-term follow-up and outcome of a large cohort of patients with common variable immunodeficiency. J. Clin. Immunol. 2007, 27, 308–316. [Google Scholar] [CrossRef] [PubMed]
- Martire, B.; Azzari, C.; Badolato, R.; Canessa, C.; Cirillo, E.; Gallo, V.; Graziani, S.; Lorenzini, T.; Milito, C.; Panza, R.; et al. Vaccination in immunocompromised host: Recommendations of Italian Primary Immunodeficiency Network Centers (IPINET). Vaccine 2018, 36, 3541–3554. [Google Scholar] [CrossRef] [PubMed]
- Voora, S.; Adey, D.B. Management of Kidney Transplant Recipients by General Nephrologists: Core Curriculum 2019. Am. J. Kidney Dis. 2019, 73, 866–879. [Google Scholar] [CrossRef] [PubMed]
- Rubin, E.J.; Longo, D.L. SARS-CoV-2 Vaccination—An Ounce (Actually, Much Less) of Prevention. N. Engl. J. Med. 2020, 383, 2677–2678. [Google Scholar] [CrossRef] [PubMed]
- Espi, M.; Charmetant, X.; Mathieu, C.; Lalande, A.; Decimo, D.; Koppe, L.; Pelletier, C.; Ovize, A.; Barbry, A.; Morelon, E.; et al. Rapid waning of immune memory against SARS-CoV-2 in maintenance hemodialysis patients after mRNA vaccination and impact of a booster dose. Kidney Int. Rep. 2023, 8, 907–911. [Google Scholar] [CrossRef]
- Amanna, I.J.; Slifka, M.K. Contributions of humoral and cellular immunity to vaccine-induced protection in humans. Virology 2011, 411, 206–215. [Google Scholar] [CrossRef]
- Eckerle, I.; Rosenberger, K.D.; Zwahlen, M.; Junghanss, T. Serologic vaccination response after solid organ transplantation: A systematic review. PLoS ONE 2013, 8, e56974. [Google Scholar] [CrossRef] [PubMed]
- Aiello, A.; Coppola, A.; Vanini, V.; Petrone, L.; Cuzzi, G.; Salmi, A.; Altera, A.M.G.; Tortorella, C.; Gualano, G.; Gasperini, C.; et al. Accuracy of QuantiFERON SARS-CoV-2 research use only assay and characterization of the CD4(+) and CD8(+) T cell-SARS-CoV-2 response: Comparison with a homemade interferon-gamma release assay. Int. J. Infect. Dis. 2022, 122, 841–849. [Google Scholar] [CrossRef] [PubMed]
- Re, D.; Seitz-Polski, B.; Brglez, V.; Carles, M.; Graca, D.; Benzaken, S.; Liguori, S.; Zahreddine, K.; Delforge, M.; Bailly-Maitre, B.; et al. Humoral and cellular responses after a third dose of SARS-CoV-2 BNT162b2 vaccine in patients with lymphoid malignancies. Nat. Commun. 2022, 13, 864. [Google Scholar] [CrossRef] [PubMed]
- Malato, J.; Ribeiro, R.M.; Fernandes, E.; Leite, P.P.; Casaca, P.; Antunes, C.; Fonseca, V.R.; Gomes, M.C.; Graca, L. Stability of hybrid versus vaccine immunity against BA.5 infection over 8 months. Lancet Infect. Dis. 2023, 23, 148–150. [Google Scholar] [CrossRef]
- Blixt, L.; Gao, Y.; Wullimann, D.; Muren Ingelman-Sundberg, H.; Muschiol, S.; Healy, K.; Bogdanovic, G.; Pin, E.; Nilsson, P.; Kjellander, C.; et al. Hybrid immunity in immunocompromised patients with CLL after SARS-CoV-2 infection followed by booster mRNA vaccination. Blood 2022, 140, 2403–2407. [Google Scholar] [CrossRef]
- Malipiero, G.; Moratto, A.; Infantino, M.; D’Agaro, P.; Piscianz, E.; Manfredi, M.; Grossi, V.; Benvenuti, E.; Bulgaresi, M.; Benucci, M.; et al. Assessment of humoral and cellular immunity induced by the BNT162b2 SARS-CoV-2 vaccine in healthcare workers, elderly people, and immunosuppressed patients with autoimmune disease. Immunol. Res. 2021, 69, 576–583. [Google Scholar] [CrossRef]
- Boyarsky, B.J.; Werbel, W.A.; Avery, R.K.; Tobian, A.A.R.; Massie, A.B.; Segev, D.L.; Garonzik-Wang, J.M. Immunogenicity of a Single Dose of SARS-CoV-2 Messenger RNA Vaccine in Solid Organ Transplant Recipients. JAMA 2021, 325, 1784–1786. [Google Scholar] [CrossRef]
- Boyarsky, B.J.; Werbel, W.A.; Avery, R.K.; Tobian, A.A.R.; Massie, A.B.; Segev, D.L.; Garonzik-Wang, J.M. Antibody Response to 2-Dose SARS-CoV-2 mRNA Vaccine Series in Solid Organ Transplant Recipients. JAMA 2021, 325, 2204–2206. [Google Scholar] [CrossRef]
- Grupper, A.; Rabinowich, L.; Schwartz, D.; Schwartz, I.F.; Ben-Yehoyada, M.; Shashar, M.; Katchman, E.; Halperin, T.; Turner, D.; Goykhman, Y.; et al. Reduced humoral response to mRNA SARS-CoV-2 BNT162b2 vaccine in kidney transplant recipients without prior exposure to the virus. Am. J. Transpl. 2021, 21, 2719–2726. [Google Scholar] [CrossRef]
- Cristelli, M.P.; Viana, L.A.; Fortaleza, C.M.; Granato, C.; Nakamura, M.R.; Santos, D.; Foresto, R.D.; Tedesco-Silva, H.; Medina-Pestana, J. Lower seroprevalence for SARS-CoV-2-specific antibodies among kidney transplant recipients compared to the general population in the city of Sao Paulo, Brazil. Transpl. Infect. Dis. 2021, 23, e13706. [Google Scholar] [CrossRef]
- Nielsen, B.U.; Drabe, C.H.; Barnkob, M.B.; Johansen, I.S.; Hansen, A.K.K.; Nilsson, A.C.; Rasmussen, L.D. Antibody response following the third and fourth SARS-CoV-2 vaccine dose in individuals with common variable immunodeficiency. Front. Immunol. 2022, 13, 934476. [Google Scholar] [CrossRef] [PubMed]
- Scalvini, D.; Schiepatti, A.; Maimaris, S.; Cosentini, E.; Muscia, R.; Gregorio, V.; Roda, E.; Fassio, F.; Baiardi, P.; Locatelli, C.A.; et al. Humoral immunogenicity of COVID-19 vaccines in patients with coeliac disease and other noncoeliac enteropathies compared to healthy controls. Eur. J. Gastroenterol. Hepatol. 2023, 35, 167–173. [Google Scholar] [CrossRef]
- Keshavarz, B.; Richards, N.E.; Workman, L.J.; Patel, J.; Muehling, L.M.; Canderan, G.; Murphy, D.D.; Brovero, S.G.; Ailsworth, S.M.; Eschenbacher, W.H.; et al. Trajectory of IgG to SARS-CoV-2 After Vaccination With BNT162b2 or mRNA-1273 in an Employee Cohort and Comparison With Natural Infection. Front. Immunol. 2022, 13, 850987. [Google Scholar] [CrossRef] [PubMed]
- Kodali, L.; Budhiraja, P.; Gea-Banacloche, J. COVID-19 in kidney transplantation-implications for immunosuppression and vaccination. Front. Med. 2022, 9, 1060265. [Google Scholar] [CrossRef]
- Alfonso-Dunn, R.; Lin, J.; Lei, J.; Liu, J.; Roche, M.; De Oliveira, A.; Raisingani, A.; Kumar, A.; Kirschner, V.; Feuer, G.; et al. Humoral and cellular responses to repeated COVID-19 exposure in multiple sclerosis patients receiving B-cell depleting therapies: A single-center, one-year, prospective study. Front. Immunol. 2023, 14, 1194671. [Google Scholar] [CrossRef]
- Hou, M.M.; Barrett, J.R.; Themistocleous, Y.; Rawlinson, T.A.; Diouf, A.; Martinez, F.J.; Nielsen, C.M.; Lias, A.M.; King, L.D.W.; Edwards, N.J.; et al. Vaccination with Plasmodium vivax Duffy-binding protein inhibits parasite growth during controlled human malaria infection. Sci. Transl. Med. 2023, 15, eadf1782. [Google Scholar] [CrossRef] [PubMed]
- Taus, E.; Shino, M.Y.; Ibarrondo, F.J.; Hausner, M.A.; Hofmann, C.; Yang, O.O. Predominantly defective CD8(+) T cell immunity to SARS-CoV-2 mRNA vaccination in lung transplant recipients. J. Transl. Med. 2023, 21, 374. [Google Scholar] [CrossRef]
- Wang, X.; Yuen, T.T.; Dou, Y.; Hu, J.; Li, R.; Zeng, Z.; Lin, X.; Gong, H.; Chan, C.H.; Yoon, C.; et al. Vaccine-induced protection against SARS-CoV-2 requires IFN-gamma-driven cellular immune response. Nat. Commun. 2023, 14, 3440. [Google Scholar] [CrossRef]
- Skelly, D.T.; Harding, A.C.; Gilbert-Jaramillo, J.; Knight, M.L.; Longet, S.; Brown, A.; Adele, S.; Adland, E.; Brown, H.; Tipton, T.; et al. Two doses of SARS-CoV-2 vaccination induce robust immune responses to emerging SARS-CoV-2 variants of concern. Nat. Commun. 2021, 12, 5061. [Google Scholar] [CrossRef]
- Milito, C.; Cinetto, F.; Palladino, A.; Garzi, G.; Punziano, A.; Lagnese, G.; Scarpa, R.; Rattazzi, M.; Pesce, A.M.; Pulvirenti, F.; et al. Mortality in Severe Antibody Deficiencies Patients during the First Two Years of the COVID-19 Pandemic: Vaccination and Monoclonal Antibodies Efficacy. Biomedicines 2022, 10, 1026. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Healthy (n = 10) | CVID (n = 19) | KTR (n = 17) |
|---|---|---|---|
| Age (y), median (range) | 39 (27–69) | 41 (23–74) | 50.7 (24–77) |
| Gender Male (%) | 4 (40%) | 10 (52.6%) | 13 (76.4%) |
| Gender Female (%) | 6 (60%) | 9 (47.3%) | 4 (23.5%) |
| Caucasian race (%) | 10 (100%) | 19 (100%) | 16 (94.1%) |
| Anti-N antibodies positive | 5 (26.3%) | 3 (17.6%) | |
| Comorbidity (%) | 0 | 6 (31.6%) | 17 (100%) |
| Autoimmune manifestation | 6 | ||
| Immune thrombocytopenia | 5 | ||
| Vitiligo | 1 | ||
| Hypertension | 15 | ||
| Ischemic heart disease | 1 | ||
| Diabetes | 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
La Civita, E.; Zannella, C.; Brusa, S.; Romano, P.; Schettino, E.; Salemi, F.; Carrano, R.; Gentile, L.; Punziano, A.; Lagnese, G.; et al. BNT162b2 Elicited an Efficient Cell-Mediated Response against SARS-CoV-2 in Kidney Transplant Recipients and Common Variable Immunodeficiency Patients. Viruses 2023, 15, 1659. https://doi.org/10.3390/v15081659
La Civita E, Zannella C, Brusa S, Romano P, Schettino E, Salemi F, Carrano R, Gentile L, Punziano A, Lagnese G, et al. BNT162b2 Elicited an Efficient Cell-Mediated Response against SARS-CoV-2 in Kidney Transplant Recipients and Common Variable Immunodeficiency Patients. Viruses. 2023; 15(8):1659. https://doi.org/10.3390/v15081659
Chicago/Turabian StyleLa Civita, Evelina, Carla Zannella, Stefano Brusa, Paolo Romano, Elisa Schettino, Fabrizio Salemi, Rosa Carrano, Luca Gentile, Alessandra Punziano, Gianluca Lagnese, and et al. 2023. "BNT162b2 Elicited an Efficient Cell-Mediated Response against SARS-CoV-2 in Kidney Transplant Recipients and Common Variable Immunodeficiency Patients" Viruses 15, no. 8: 1659. https://doi.org/10.3390/v15081659
APA StyleLa Civita, E., Zannella, C., Brusa, S., Romano, P., Schettino, E., Salemi, F., Carrano, R., Gentile, L., Punziano, A., Lagnese, G., Spadaro, G., Franci, G., Galdiero, M., Terracciano, D., Portella, G., & Loffredo, S. (2023). BNT162b2 Elicited an Efficient Cell-Mediated Response against SARS-CoV-2 in Kidney Transplant Recipients and Common Variable Immunodeficiency Patients. Viruses, 15(8), 1659. https://doi.org/10.3390/v15081659