
Detection and Genetic Characterization of Astroviruses in Brain Tissues of Wild Raccoon Dogs


Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Viral RNA Extraction and cDNA Synthesis
2.3. Astrovirus Screening and ORF2 PCR
2.4. Genetic Analysis of Astrovirus Sequences
3. Results
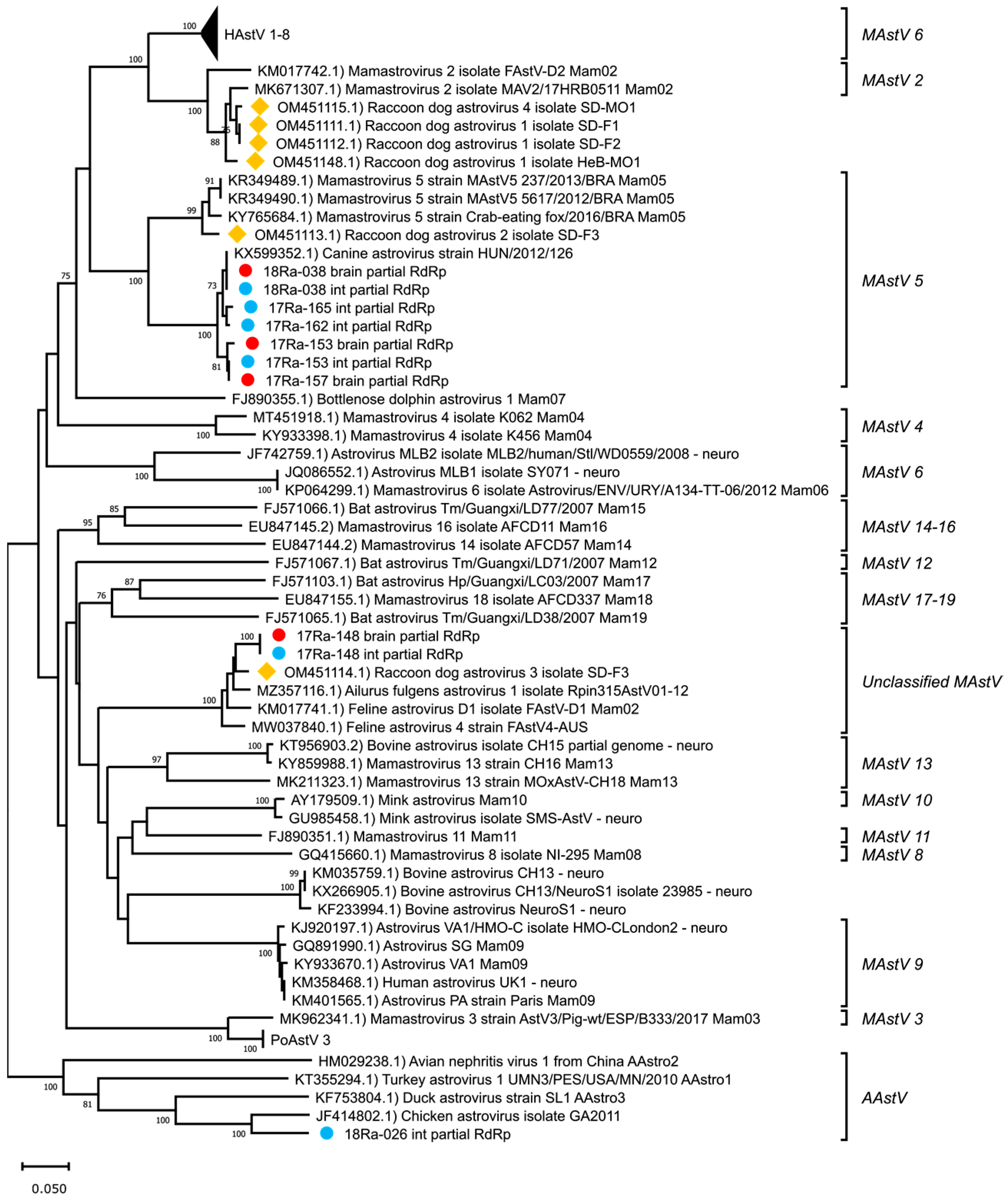
3.1. Detection of Raccoon Dog Astrovirus
3.2. Genetic Analysis of Partial RdRp Sequences
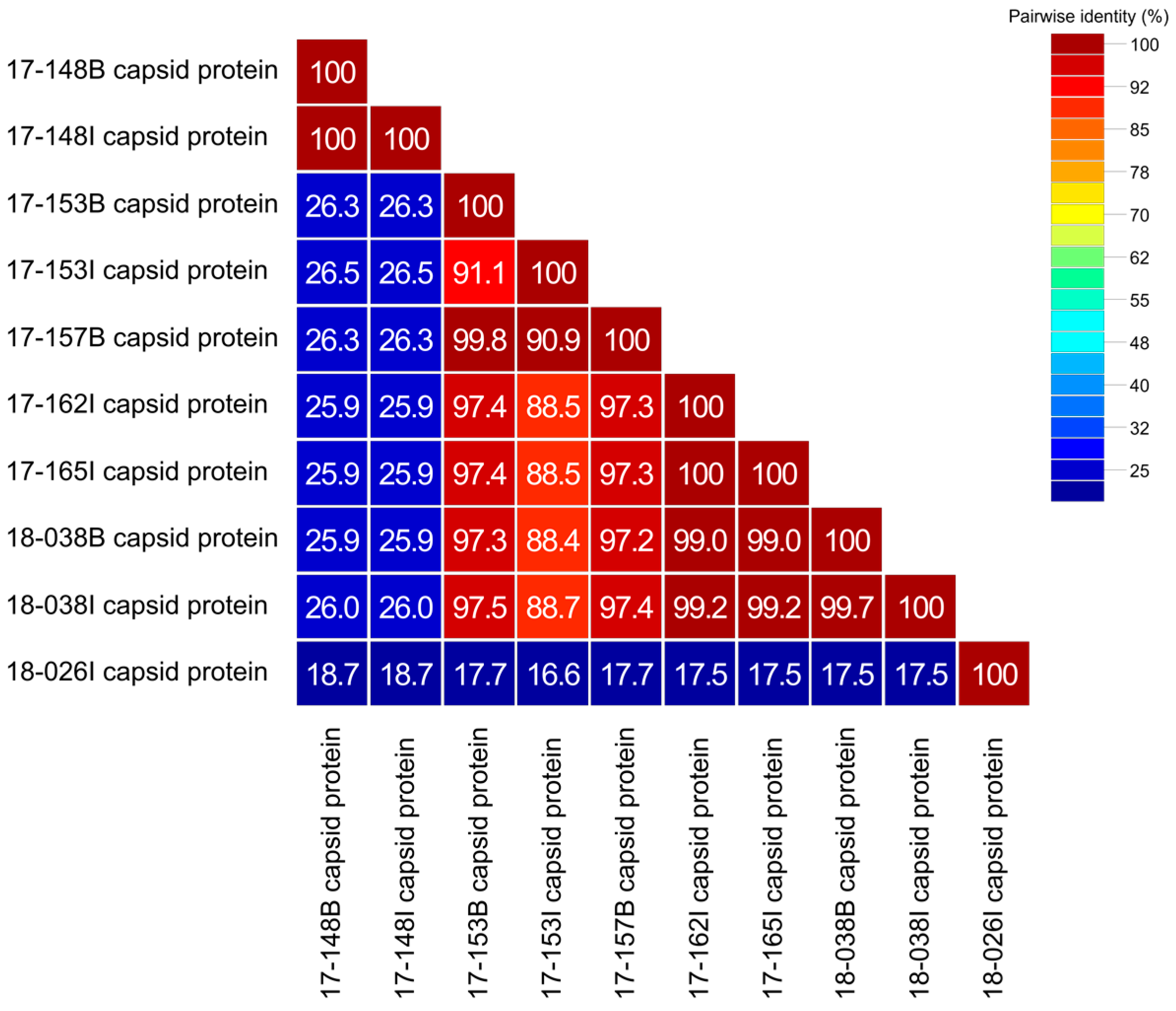
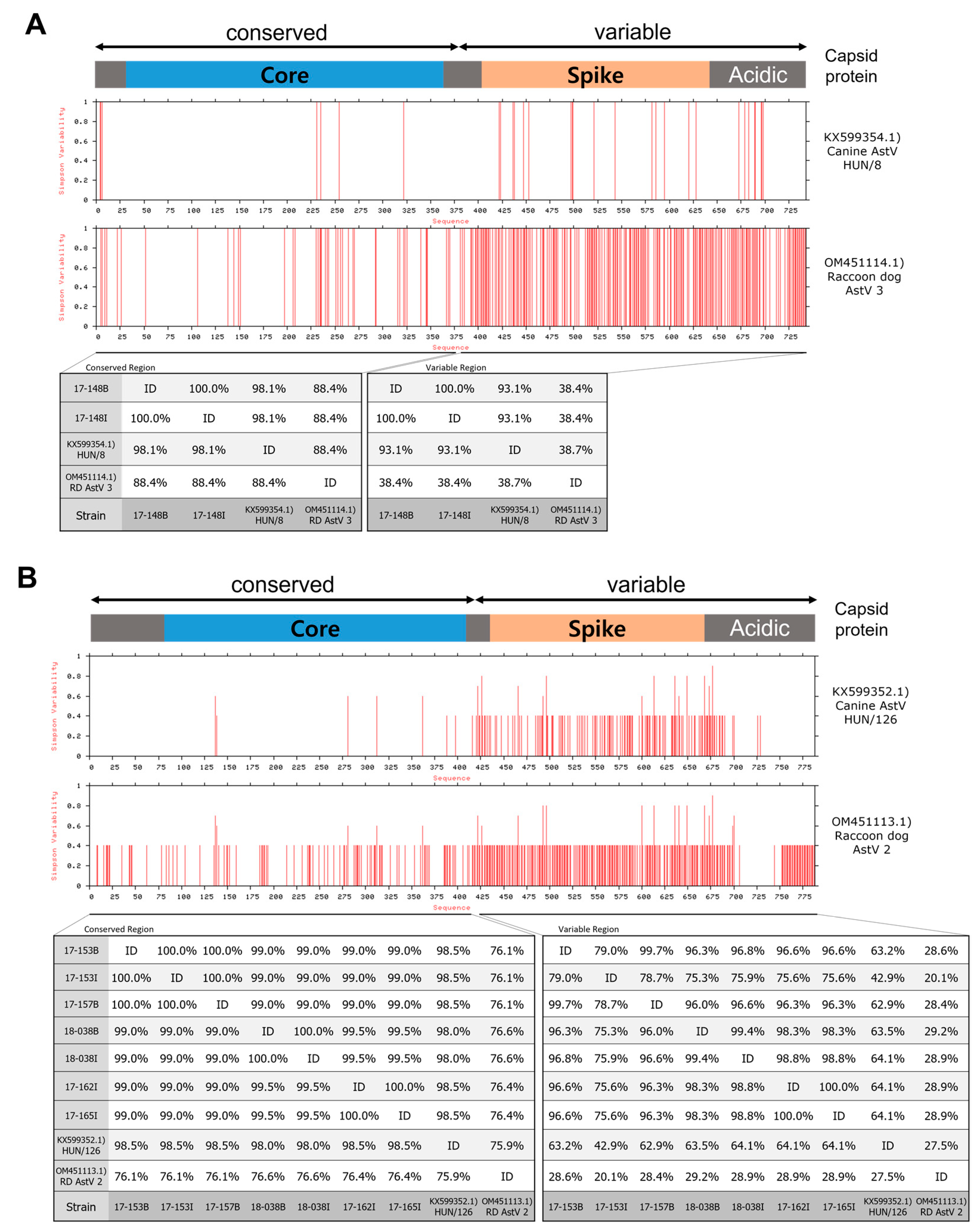
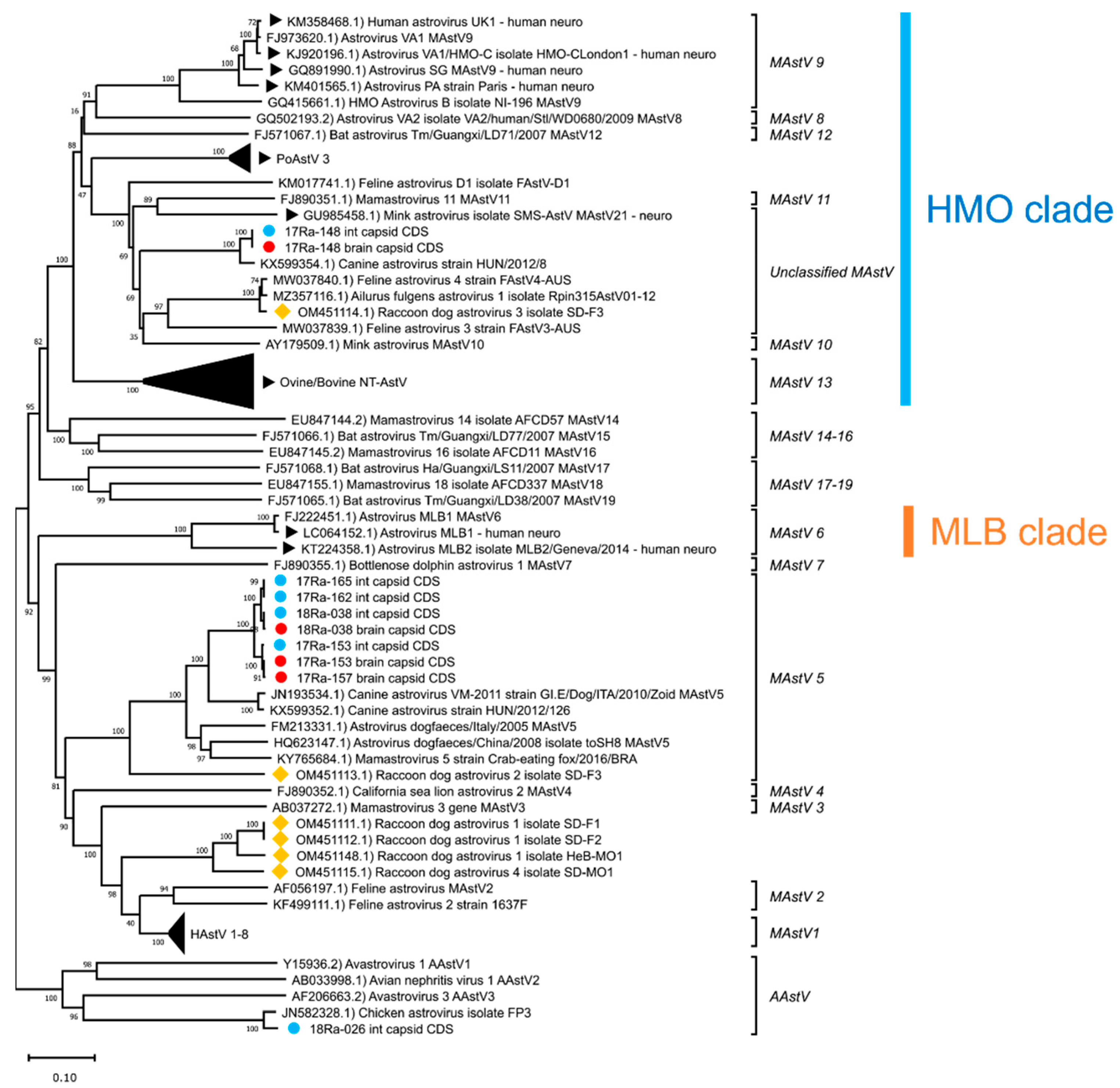
3.3. Genetic Analysis of Complete Capsid Protein Amino Acid Sequences
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bouzalas, I.G.; Wüthrich, D.; Selimovic-Hamza, S.; Drögemüller, C.; Bruggmann, R.; Seuberlich, T. Full-genome based molecular characterization of encephalitis-associated bovine astroviruses. Infect. Genet. Evol. 2016, 44, 162–168. [Google Scholar] [CrossRef]
- Cortez, V.; Meliopoulos, V.A.; Karlsson, E.A.; Hargest, V.; Johnson, C.; Schultz-Cherry, S. Astrovirus Biology and Pathogenesis. Annu. Rev. Virol. 2017, 4, 327–348. [Google Scholar] [CrossRef]
- Guix, S.; Bosch, A.; Pintó, R. Astrovirus Taxonomy. In Astrovirus Research: Essential Ideas, Everyday Impacts, Future Directions; Springer: Berlin/Heidelberg, Germany, 2012; pp. 97–118. [Google Scholar]
- Janowski, A.B. Beyond the Gastrointestinal Tract: The Emerging and Diverse Tissue Tropisms of Astroviruses. Viruses 2021, 13, 732. [Google Scholar] [CrossRef] [PubMed]
- Sun, N.; Yang, Y.; Wang, G.-S.; Shao, X.-Q.; Zhang, S.-Q.; Wang, F.-X.; Tan, B.; Tian, F.-L.; Cheng, S.-P.; Wen, Y.-J. Detection and characterization of avastrovirus associated with diarrhea isolated from minks in China. Food Environ. Virol. 2014, 6, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, E.; Small, C.T.; Freiden, P.; Feeroz, M.M.; Iv, F.A.M.; San, S.; Hasan, M.K.; Wang, D.; Jones-Engel, L.; Schultz-Cherry, S. Non-Human Primates Harbor Diverse Mammalian and Avian Astroviruses Including Those Associated with Human Infections. PLOS Pathog. 2015, 11, e1005225. [Google Scholar] [CrossRef]
- Wohlgemuth, N.; Honce, R.; Schultz-Cherry, S. Astrovirus evolution and emergence. Infect. Genet. Evol. 2019, 69, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Pan, M.; Wang, X.; Xu, Y.; Xie, X.; Knowles, N.J.; Yang, H.; Zhang, D. Complete sequence of a duck astrovirus associated with fatal hepatitis in ducklings. J. Gen. Virol. 2009, 90 Pt 5, 1104–1108. [Google Scholar] [CrossRef] [PubMed]
- Bosch, A.; Pintó, R.M.; Guix, S. Human astroviruses. Clin. Microbiol. Rev. 2014, 27, 1048–1074. [Google Scholar] [CrossRef]
- Quan, P.-L.; Wagner, T.A.; Briese, T.; Torgerson, T.R.; Hornig, M.; Tashmukhamedova, A.; Firth, C.; Palacios, G.; Baisre-De-Leon, A.; Paddock, C.D.; et al. Astrovirus encephalitis in boy with X-linked agammaglobulinemia. Emerg. Infect. Dis. 2010, 16, 918–925. [Google Scholar] [CrossRef]
- Wildi, N.; Seuberlich, T. Neurotropic Astroviruses in Animals. Viruses 2021, 13, 1201. [Google Scholar] [CrossRef]
- Reuter, G.; Pankovics, P.; Boros, Á. Nonsuppurative (Aseptic) Meningoencephalomyelitis Associated with Neurovirulent Astrovirus Infections in Humans and Animals. Clin. Microbiol. Rev. 2018, 31, e00040-18. [Google Scholar] [CrossRef] [PubMed]
- Kauer, R.V.; Koch, M.C.; Schönecker, L.; Becker, J.; Holwerda, M.; Glaus, A.N.; Hierweger, M.M.; Werder, S.; Dijkman, R.; Meylan, M.; et al. Fecal Shedding of Bovine Astrovirus CH13/NeuroS1 in Veal Calves. J. Clin. Microbiol. 2020, 58, e01964-19. [Google Scholar] [CrossRef] [PubMed]
- Jakubczak, A.; Kowalczyk, M.; Mazurkiewicz, I.; Kondracki, M. Detection of mink astrovirus in Poland and further phylogenetic comparison with other European and Canadian astroviruses. Virus Genes 2021, 57, 258–265. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.-K.; Lee, S.H.; Kim, H.-H.; Kim, J.-T.; Ahn, S.; Cho, I.-S. Detection of viral infections in wild Korean raccoon dogs (Nyctereutes procyonoides koreensis). Korean J. Veter- Res. 2017, 57, 209–214. [Google Scholar] [CrossRef]
- Matysiak, A.; Wasielewski, O.; Wlodarek, J.; Ondrejkova, A.; Tryjanowski, P. First report of ticks in the subcutaneous tissue of the raccoon dog Nyctereutes procyonoides. Veterinární Med. 2018, 63, 571–574. [Google Scholar] [CrossRef]
- Yang, S.; He, Y.; Chen, X.; Kalim, U.; Wang, Y.; Yang, S.; Qi, H.; Cheng, H.; Lu, X.; Wang, X.; et al. Viral Metagenomics Reveals Diverse Viruses in the Feces Samples of Raccoon Dogs. Front. Vet. Sci. 2021, 8, 693564. [Google Scholar] [CrossRef]
- He, W.-T.; Hou, X.; Zhao, J.; Sun, J.; He, H.; Si, W.; Wang, J.; Jiang, Z.; Yan, Z.; Xing, G.; et al. Virome characterization of game animals in China reveals a spectrum of emerging pathogens. Cell 2022, 185, 1117–1129.e8. [Google Scholar] [CrossRef]
- Hong, Y.J.; Kim, K.S.; Min, M.-S.; Lee, H. Population structure of the raccoon dog (Nyctereutes procyonoides) using microsatellite loci analysis in South Korea: Implications for disease management. J. Veter.-Med. Sci. 2018, 80, 1631–1638. [Google Scholar] [CrossRef]
- Sutor, A.; Schwarz, S.; Conraths, F.J. The biological potential of the raccoon dog (Nyctereutes procyonoides, Gray 1834) as an invasive species in Europe-new risks for disease spread? Acta Theriol. 2014, 59, 49–59. [Google Scholar] [CrossRef]
- Zhang, X.; McFarland, T.J.; Vartanian, K.; Zhu, Y.; Harrington, C.A.; Chu, C.Q. RNA isolation from micro-quantity of articular cartilage for quantitative gene expression by microarray analysis. Int. J. Med. Sci. 2022, 19, 98–104. [Google Scholar] [CrossRef]
- Chu, D.K.W.; Poon, L.L.M.; Guan, Y.; Peiris, J.S.M. Novel astroviruses in insectivorous bats. J. Virol. 2008, 82, 9107–9114. [Google Scholar] [CrossRef]
- Toffan, A.; Jonassen, C.M.; De Battisti, C.; Schiavon, E.; Kofstad, T.; Capua, I.; Cattoli, G. Genetic characterization of a new astrovirus detected in dogs suffering from diarrhoea. Veter. Microbiol. 2009, 139, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Zhou, J.; Hou, Y.; Wang, R.; Li, Q.; Wang, Y.; Yuan, R.; Chen, P.; Wang, B.; Xia, X. Diversity and Genetic Characteristics of Astroviruses from Animals in Yunnan Province. Viruses 2022, 14, 2234. [Google Scholar] [CrossRef]
- Fernández-Correa, I.; Truchado, D.A.; Gomez-Lucia, E.; Doménech, A.; Pérez-Tris, J.; Schmidt-Chanasit, J.; Cadar, D.; Benítez, L. A novel group of avian astroviruses from Neotropical passerine birds broaden the diversity and host range of Astroviridae. Sci. Rep. 2019, 9, 9513. [Google Scholar] [CrossRef] [PubMed]
- De Benedictis, P.; Schultz-Cherry, S.; Burnham, A.; Cattoli, G. Astrovirus infections in humans and animals—Molecular biology, genetic diversity, and interspecies transmissions. Infect. Genet. Evol. 2011, 11, 1529–1544. [Google Scholar] [CrossRef] [PubMed]
- Arias, C.F.; DuBois, R.M. The Astrovirus Capsid: A Review. Viruses 2017, 9, 15. [Google Scholar] [CrossRef]
- Caddy, S.L.; Goodfellow, I. Complete genome sequence of canine astrovirus with molecular and epidemiological characterisation of UK strains. Vet. Microbiol. 2015, 177, 206–213. [Google Scholar] [CrossRef]
- Yi, S.; Niu, J.; Wang, H.; Dong, G.; Guo, Y.; Dong, H.; Wang, K.; Hu, G. Molecular characterization of feline astrovirus in domestic cats from Northeast China. PLoS ONE 2018, 13, e0205441. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, R.; Liang, J.; Zhao, N.; Li, G.; Gao, Q.; Su, S. Epidemiology, genetic diversity and evolution of canine astrovirus. Transbound. Emerg. Dis. 2020, 67, 2901–2910. [Google Scholar] [CrossRef]
- Blomström, A.L.; Ley, C.; Jacobson, M. Astrovirus as a possible cause of congenital tremor type AII in piglets? Acta Vet. Scand. 2014, 56, 82. [Google Scholar] [CrossRef]
- Alves, C.D.B.T.; Budaszewski, R.D.F.; Cibulski, S.P.; Weber, M.N.; Mayer, F.Q.; Bianchi, M.V.; Zafalon-Silva, B.; Konradt, G.; Slaviero, M.; Sonne, L.; et al. Mamastrovirus 5 detected in a crab-eating fox (Cerdocyon thous): Expanding wildlife host range of astroviruses. Comp. Immunol. Microbiol. Infect. Dis. 2018, 58, 36–43. [Google Scholar]
- Sato, M.; Kuroda, M.; Kasai, M.; Matsui, H.; Fukuyama, T.; Katano, H.; Tanaka-Taya, K. Acute encephalopathy in an immunocompromised boy with astrovirus-MLB1 infection detected by next generation sequencing. J. Clin. Virol. 2016, 78, 66–70. [Google Scholar] [CrossRef] [PubMed]
- Lawler, P.E.; Cook, K.A.; Williams, H.G.; Archer, L.L.; Schaedel, K.E.; Isaza, N.M.; Wellehan, J.J.F.X. Determination of the diversity of astroviruses in feces from cats in Florida. J. Veter.-Diagn. Investig. 2018, 30, 275–279. [Google Scholar] [CrossRef] [PubMed]
- Meyer, C.T.; Bauer, I.K.; Antonio, M.; Adeyemi, M.; Saha, D.; Oundo, J.O.; Ochieng, J.B.; Omore, R.; Stine, O.C.; Wang, D.; et al. Prevalence of classic, MLB-clade and VA-clade Astroviruses in Kenya and The Gambia. Virol. J. 2015, 12, 78. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Position | Sequence | Reference | |
|---|---|---|---|---|
| RAD AstV 148 | RdRp | F1 | TGG ATG AGC AAT ATC AGA CAC C | customized primer |
| RAD AstV 153 | CAT CAA GAG GCT ACG CTG G | |||
| RAD AstV 157 | ||||
| RAD AstV 038 | TAT CAA GAA ACT ACG CTG G | |||
| RAD AstV 026 | TAG CCT CAA AGT ATA AGA CGC A | |||
| s2m_rev | ORF2 | R | CCC TCG ATC CTA CTC GG | [23] |
| RAD AstV 148 | RdRp | F2 | CGA TTG GTA TTG TAA GAA CAT C | customized primer |
| RAD AstV 153 | TGG TTA ATG CCG AGC AGC GGA A | |||
| RAD AstV 157 | ||||
| RAD AstV 038 | ||||
| RAD AstV 026 | ACA AGG GGT TGT TCG ATT G |
| Raccoon Dog No. | Case no. | Region | AstV Positive | Prevalence | ||
|---|---|---|---|---|---|---|
| Brain Tissue | Intestine Tissue | Brain | Intestine | |||
| 17-148 | KNU-082 | Gyeonggi-do | + | + | 3% (4/133) | 7.8% (6/77) |
| 17-153 | KNU-087 | Gyeonggi-do | + | + | ||
| 17-157 | KNU-091 | Gangwon-do | + | - | ||
| 17-162 | KNU-096 | Gangwon-do | - | + | ||
| 17-165 | KNU-099 | Gyeonggi-do | - | + | ||
| 18-026 | CB-010 | Chungcheong-do | - | + | ||
| 18-038 | KNU-027 | Gyeonggi-do | + | + | ||
| Strain | Canine AstV Strain HUN 126 (KX599352.1) | Ailurus fulgens AstV 1 (MZ357116.1) | Chicken AstV Isolate GA2011 (JF414802.1) | Raccoon Dog AstV (China Strain) |
|---|---|---|---|---|
| 17-148B | 55.6% | 90.1% | 45.2% | 54.6–89.3% |
| 17-148I | 55.6% | 90.1% | 45.2% | 54.6–89.3% |
| 17-153B | 93.2% | 55.1% | 45.0% | 55.6–74.4% |
| 17-153I | 94.2% | 55.6% | 46.0% | 56.2–75.5% |
| 17-157B | 94.2% | 55.6% | 46.0% | 56.2–75.5% |
| 17-162I | 95.1% | 54.9% | 45.2% | 54.9–75.2% |
| 17-165I | 94.8% | 54.6% | 45.0% | 54.6–75.0% |
| 18-038B | 97.5% | 56.7% | 45.7% | 56.2–75.0% |
| 18-038I | 97.5% | 56.7% | 45.7% | 56.2–75.0% |
| 18-026I | 45.2% | 46.2% | 76.7% | 43.3–47.2% |
| Strain | Canine AstV Strain HUN/126 (KX599352.1) | Canine AstV Strain HUN/8 (KX599354.1) | Chicken AstV (JN582328.1) | Raccoon Dog AstV (China Strain) |
|---|---|---|---|---|
| 17-148B | 18.4% | 91.4% | 13.4% | 17.2–59.1% |
| 17-148I | 18.4% | 91.4% | 13.4% | 17.2–59.1% |
| 17-153B | 83.5% | 19.3% | 14.0% | 18.3–49.8% |
| 17-153I | 74.9% | 19.5% | 13.2% | 18.0–47.2% |
| 17-157B | 83.4% | 19.3% | 14.0% | 18.3–49.8% |
| 17-162I | 83.5% | 19.4% | 14.1% | 18.4–50.0% |
| 17-165I | 83.5% | 19.4% | 14.1% | 18.4–50.0% |
| 18-038B | 83.0% | 19.4% | 14.0% | 18.4–50.2% |
| 18-038I | 83.3% | 19.5% | 14.0% | 18.4–50.1% |
| 18-026I | 13.9% | 12.6% | 96.3% | 13.1–16.0% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chae, S.-B.; Jeong, C.-G.; Park, J.-S.; Na, E.-J.; Oem, J.-K. Detection and Genetic Characterization of Astroviruses in Brain Tissues of Wild Raccoon Dogs. Viruses 2023, 15, 1488. https://doi.org/10.3390/v15071488
Chae S-B, Jeong C-G, Park J-S, Na E-J, Oem J-K. Detection and Genetic Characterization of Astroviruses in Brain Tissues of Wild Raccoon Dogs. Viruses. 2023; 15(7):1488. https://doi.org/10.3390/v15071488
Chicago/Turabian StyleChae, Su-Beom, Chang-Gi Jeong, Jun-Soo Park, Eun-Jee Na, and Jae-Ku Oem. 2023. "Detection and Genetic Characterization of Astroviruses in Brain Tissues of Wild Raccoon Dogs" Viruses 15, no. 7: 1488. https://doi.org/10.3390/v15071488
APA StyleChae, S.-B., Jeong, C.-G., Park, J.-S., Na, E.-J., & Oem, J.-K. (2023). Detection and Genetic Characterization of Astroviruses in Brain Tissues of Wild Raccoon Dogs. Viruses, 15(7), 1488. https://doi.org/10.3390/v15071488