
Influence of N-Glycosylation on Virus–Host Interactions in Halorubrum lacusprofundi

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
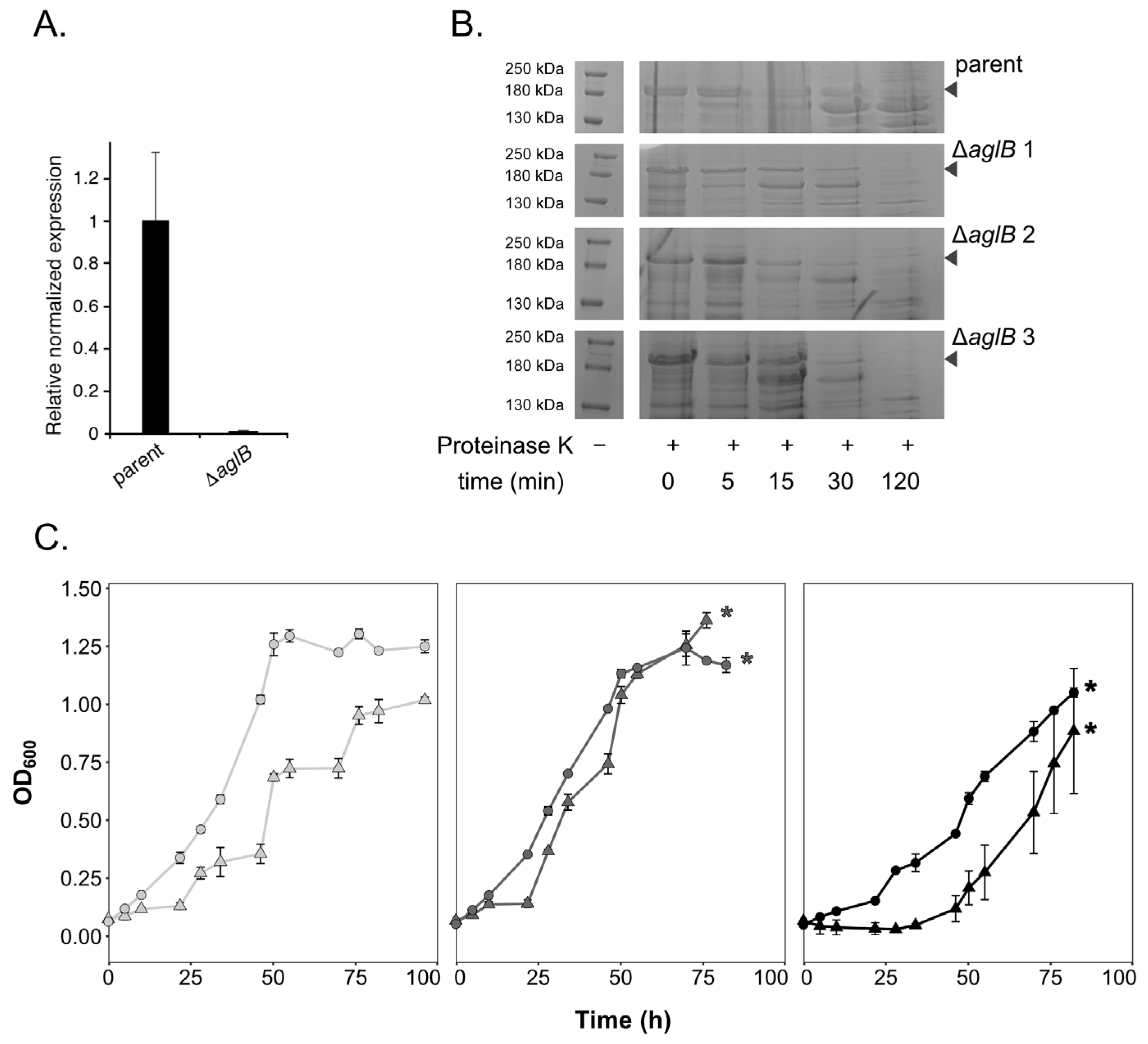
2.1. Strains and Cultivation Conditions
2.2. Deletion of Hlac_1062 (aglB)
2.3. Proteinase K Digestion of the S-Layer Glycoprotein
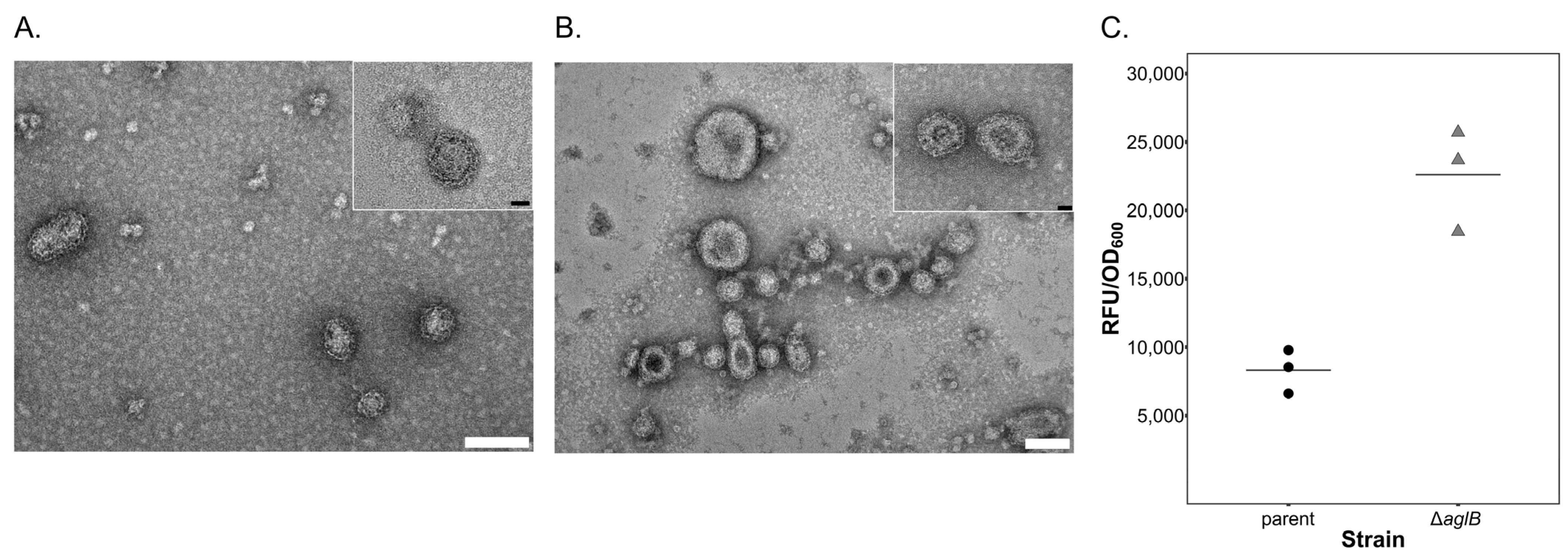
2.4. Quantification of Extracellular Vesicles
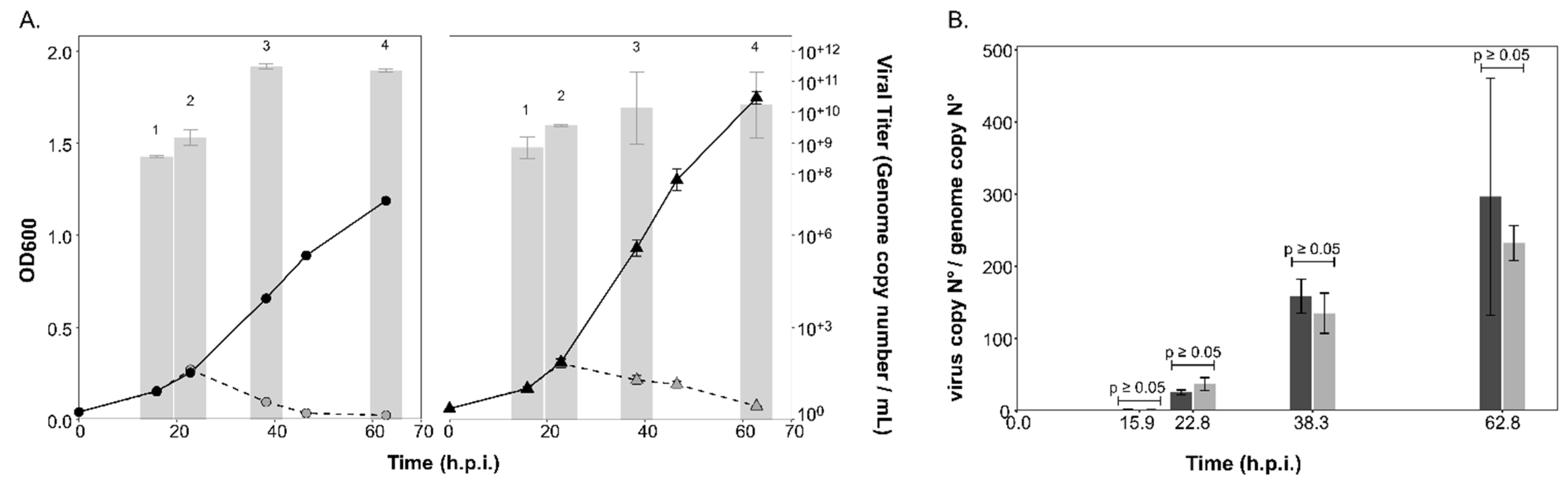
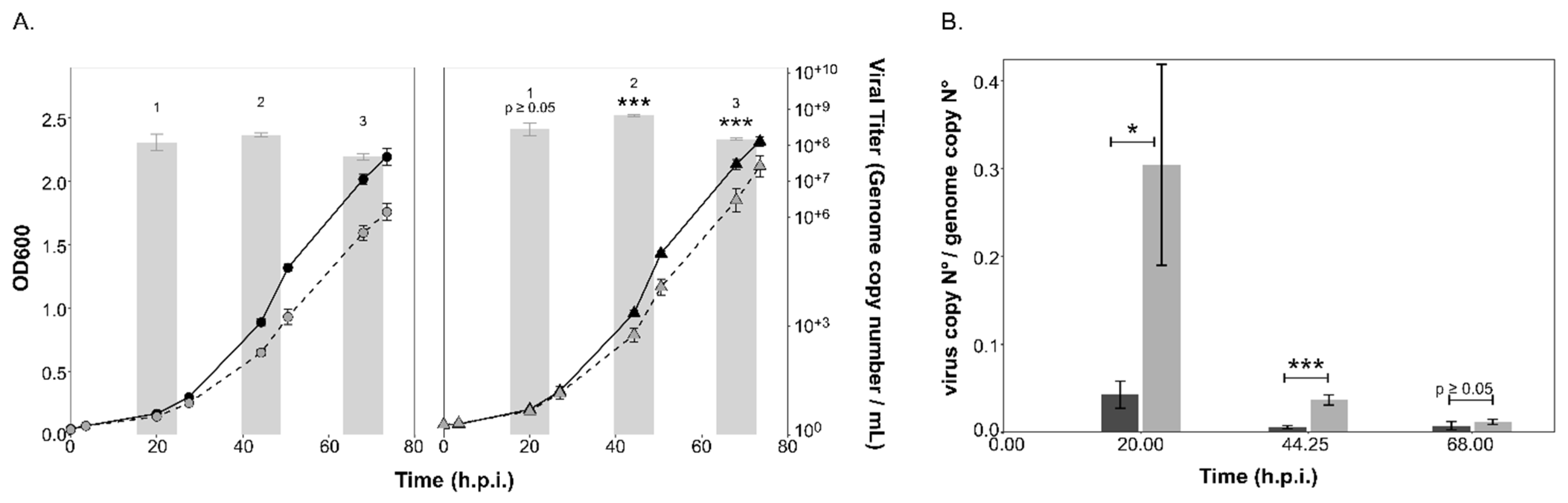
2.5. Isolation of PVs and Viruses, Infection and Growth Experiments
2.6. Virus and PV Quantification
2.7. Glycostaining of Cells and Particles
2.8. Determination of Virus Stability
3. Results
3.1. Deletion of Hrr. lacusprofundi aglB Leads to Growth Deficiencies at Varying Salt Concentrations and S-Layer Instability
3.2. The Life cycle of the Lytic Virus HRTV-DL1 Is not Influenced by Changes in Host N-Glycosylation
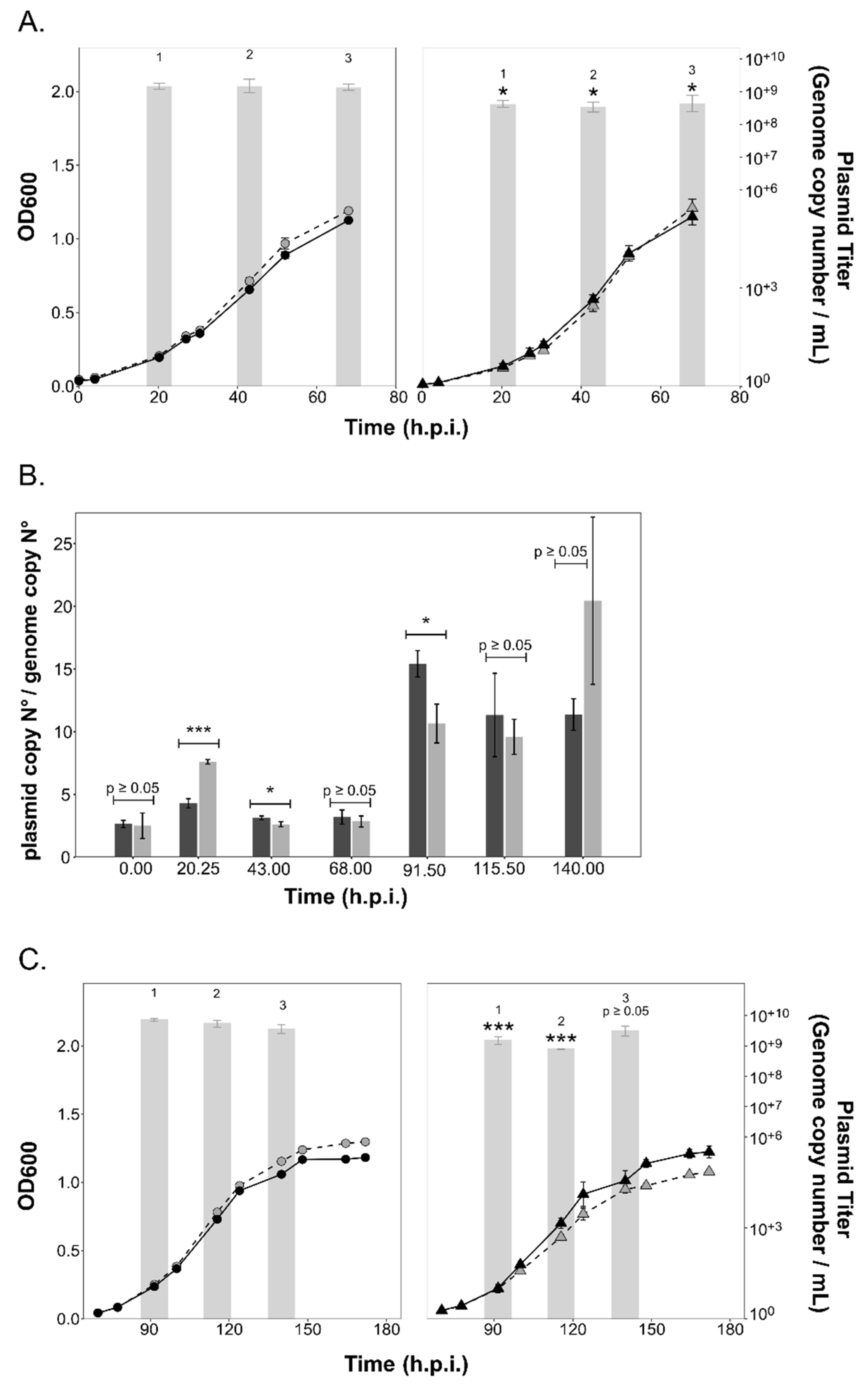
3.3. Intracellular and Extracellular Virus Numbers of the Chronic Virus HFPV-1 Are Significantly Increased in the ∆aglB Mutant
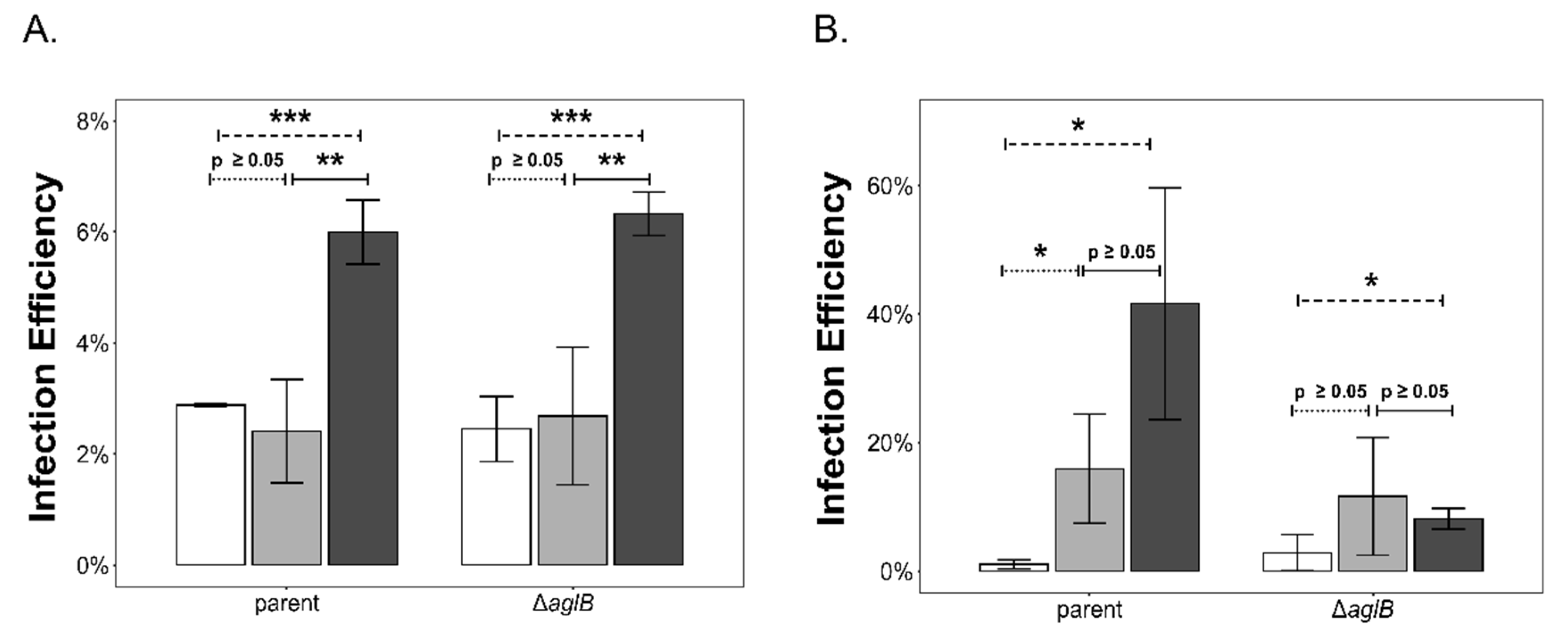
3.4. PV Production Is Reduced in the aglB Deletion Strain
3.5. Impaired Glycosylation has an Impact on the Stability of HFPV-1
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jarrell, K.F.; Ding, Y.; Meyer, B.H.; Albers, S.-V.; Kaminski, L.; Eichler, J. N-Linked Glycosylation in Archaea: A Structural, Functional, and Genetic Analysis. Microbiol. Mol. Biol. Rev. 2014, 78, 304–341. [Google Scholar] [CrossRef] [PubMed]
- Eichler, J. N-glycosylation in Archaea—New roles for an ancient posttranslational modification. Mol. Microbiol. 2020, 114, 735–741. [Google Scholar] [CrossRef] [PubMed]
- Magidovich, H.; Eichler, J. Glycosyltransferases and oligosaccharyltransferases in Archaea: Putative components of the N-glycosylation pathway in the third domain of life. FEMS Microbiol. Lett. 2009, 300, 122–130. [Google Scholar] [CrossRef] [PubMed]
- Kaminski, L.; Lurie-Weinberger, M.N.; Allers, T.; Gophna, U.; Eichler, J. Phylogenetic- and genome-derived insight into the evolution of N-glycosylation in Archaea. Mol. Phylogenet. Evol. 2013, 68, 327–339. [Google Scholar] [CrossRef]
- Abdul Halim, M.F.; Karch, K.R.; Zhou, Y.; Haft, D.H.; Garcia, B.A.; Pohlschroder, M. Permuting the PGF Signature Motif Blocks both Archaeosortase-Dependent C-Terminal Cleavage and Prenyl Lipid Attachment for the Haloferax volcanii S-Layer Glycoprotein. J. Bacteriol. 2015, 198, 808–815. [Google Scholar] [CrossRef] [PubMed]
- Nikolayev, S.; Cohen-Rosenzweig, C.; Eichler, J. Evolutionary considerations of the oligosaccharyltransferase AglB and other aspects of N-glycosylation across Archaea. Mol. Phylogenetics Evol. 2020, 153, 106951. [Google Scholar] [CrossRef]
- Yurist-Doutsch, S.; Magidovich, H.; Ventura, V.V.; Hitchen, P.G.; Dell, A.; Eichler, J. N-glycosylation in Archaea: On the coordinated actions of Haloferax volcanii AglF and AglM. Mol. Microbiol. 2010, 75, 1047–1058. [Google Scholar] [CrossRef]
- Tamir, A.; Eichler, J. N-glycosylation Is Important for Proper Haloferax volcanii S-Layer Stability and Function. Appl. Environ. Microbiol. 2017, 83, e03152-16. [Google Scholar] [CrossRef]
- Vershinin, Z.; Zaretsky, M.; Guan, Z.; Eichler, J. Identifying Components of a Halobacterium salinarum N-glycosylation Pathway. Front. Microbiol. 2021, 12, 779599. [Google Scholar] [CrossRef]
- Shalev, Y.; Turgeman-Grott, I.; Tamir, A.; Eichler, J.; Gophna, U. Cell Surface Glycosylation Is Required for Efficient Mating of Haloferax volcanii. Front. Microbiol. 2017, 8, 1253. [Google Scholar] [CrossRef]
- Shalev, Y.; Soucy, S.M.; Papke, R.T.; Gogarten, J.P.; Eichler, J.; Gophna, U. Comparative Analysis of Surface Layer Glycoproteins and Genes Involved in Protein Glycosylation in the Genus Haloferax. Genes 2018, 9, 172. [Google Scholar] [CrossRef] [PubMed]
- Kandiba, L.; Aitio, O.; Helin, J.; Guan, Z.; Permi, P.; Bamford, D.H.; Eichler, J.; Roine, E. Diversity in prokaryotic glycosylation: An archaeal-derived N-linked glycan contains legionaminic acid. Mol. Microbiol. 2012, 84, 578–593. [Google Scholar] [CrossRef] [PubMed]
- Zaretsky, M.; Roine, E.; Eichler, J. Sialic Acid-Like Sugars in Archaea: Legionaminic Acid Biosynthesis in the Halophile Halorubrum sp. PV6. Front. Microbiol. 2018, 9, 2133. [Google Scholar] [CrossRef]
- Abu-Qarn, M.; Giordano, A.; Battaglia, F.; Trauner, A.; Hitchen, P.G.; Morris, H.R.; Dell, A.; Eichler, J. Identification of AglE, a Second Glycosyltransferase Involved in N Glycosylation of the Haloferax volcanii S-Layer Glycoprotein. J. Bacteriol. 2008, 190, 3140–3146. [Google Scholar] [CrossRef]
- Yurist-Doutsch, S.; Abu-Qarn, M.; Battaglia, F.; Morris, H.R.; Hitchen, P.G.; Dell, A.; Eichler, J. aglF, aglG and aglI, novel members of a gene island involved in the N-glycosylation of the Haloferax volcanii S-layer glycoprotein. Mol. Microbiol. 2008, 69, 1234–1245. [Google Scholar] [CrossRef] [PubMed]
- Guan, Z.; Naparstek, S.; Kaminski, L.; Konrad, Z.; Eichler, J. Distinct glycan-charged phosphodolichol carriers are required for the assembly of the pentasaccharide N-linked to the Haloferax volcanii S-layer glycoprotein. Mol. Microbiol. 2010, 78, 1294–1303. [Google Scholar] [CrossRef]
- Kaminski, L.; Eichler, J. Identification of Residues Important for the Activity of Haloferax volcanii AglD, a Component of the Archaeal N-glycosylation Pathway. Archaea 2010, 2010, 315108. [Google Scholar] [CrossRef]
- Abu-Qarn, M.; Yurist-Doutsch, S.; Giordano, A.; Trauner, A.; Morris, H.R.; Hitchen, P.; Medalia, O.; Dell, A.; Eichler, J. Haloferax volcanii AglB and AglD are Involved in N-glycosylation of the S-layer Glycoprotein and Proper Assembly of the Surface Layer. J. Mol. Biol. 2007, 374, 1224–1236. [Google Scholar] [CrossRef]
- Kandiba, L.; Lin, C.W.; Aebi, M.; Eichler, J.; Guerardel, Y. Structural characterization of the N-linked pentasaccharide decorating glycoproteins of the halophilic archaeon Haloferax volcanii. Glycobiology 2016, 26, 745–756. [Google Scholar] [CrossRef]
- Kohda, D. Structural Basis of Protein Asn-Glycosylation by Oligosaccharyltransferases. In Glycobiophysics; Yamaguchi, Y., Kato, K., Eds.; Springer: Singapore, 2018; pp. 171–199. [Google Scholar] [CrossRef]
- Cohen-Rosenzweig, C.; Yurist-Doutsch, S.; Eichler, J. AglS, a Novel Component of the Haloferax volcanii N-glycosylation Pathway, Is a Dolichol Phosphate-Mannose Mannosyltransferase. J. Bacteriol. 2012, 194, 6909–6916. [Google Scholar] [CrossRef]
- Kaminski, L.; Guan, Z.; Abu-Qarn, M.; Konrad, Z.; Eichler, J. AglR is required for addition of the final mannose residue of the N-linked glycan decorating the Haloferax volcanii S-layer glycoprotein. Biochim. Et Biophys. Acta (BBA) Gen. Subj. 2012, 1820, 1664–1670. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Pfeiffer, F.; Losensky, G.; Marchfelder, A.; Habermann, B.; Dyall-Smith, M. Whole-genome comparison between the type strain of Halobacterium salinarum (DSM 3754T) and the laboratory strains R1 and NRC-1. Microbiologyopen 2020, 9, e974. [Google Scholar] [CrossRef] [PubMed]
- Bignon, E.A.; Chou, K.R.; Roine, E.; Tischler, N.D. Halorubrum pleomorphic virus-6 Membrane Fusion Is Triggered by an S-Layer Component of Its Haloarchaeal Host. Viruses 2022, 14, 254. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Demina, T.A.; Roux, S.; Aiewsakun, P.; Kazlauskas, D.; Simmonds, P.; Prangishvili, D.; Oksanen, H.M.; Krupovic, M. Diversity, taxonomy, and evolution of archaeal viruses of the class Caudoviricetes. PLoS Biol. 2021, 19, e3001442. [Google Scholar] [CrossRef]
- Mercier, C.; Thies, D.; Zhong, L.; Raftery, M.J.; Cavicchioli, R.; Erdmann, S. In depth characterization of an archaeal virus-host system reveals numerous virus exclusion mechanisms. bioRxiv 2022. [Google Scholar] [CrossRef]
- Schwarzer, S.; Hackl, T.; Oksanen Hanna, M.; Quax Tessa, E.F. Archaeal Host Cell Recognition and Viral Binding of HFTV1 to Its Haloferax Host. mBio 2023, 14, e01833-22. [Google Scholar] [CrossRef]
- Zink, I.A.; Pfeifer, K.; Wimmer, E.; Sleytr, U.B.; Schuster, B.; Schleper, C. CRISPR-mediated gene silencing reveals involvement of the archaeal S-layer in cell division and virus infection. Nat. Commun. 2019, 10, 4797. [Google Scholar] [CrossRef]
- Quemin, E.R.J.; Chlanda, P.; Sachse, M.; Forterre, P.; Prangishvili, D.; Krupovic, M. Eukaryotic-Like Virus Budding in Archaea. mBio 2016, 7, e01439-16. [Google Scholar] [CrossRef]
- Quemin, E.R.J.; Pietilä, M.K.; Oksanen, H.M.; Forterre, P.; Rijpstra, W.I.C.; Schouten, S.; Bamford, D.H.; Prangishvili, D.; Krupovic, M. Sulfolobus Spindle-Shaped Virus 1 Contains Glycosylated Capsid Proteins, a Cellular Chromatin Protein, and Host-Derived Lipids. J. Virol. 2015, 89, 11681–11691. [Google Scholar] [CrossRef]
- Han, Z.; Yuan, W.; Xiao, H.; Wang, L.; Zhang, J.; Peng, Y.; Cheng, L.; Liu, H.; Huang, L. Structural insights into a spindle-shaped archaeal virus with a sevenfold symmetrical tail. Proc. Natl. Acad. Sci. USA 2022, 119, e2119439119. [Google Scholar] [CrossRef]
- Hartman, R.; Biewenga, L.; Munson-McGee, J.; Refai, M.; Boyd, E.S.; Bothner, B.; Lawrence, C.M.; Young, M. Discovery and Characterization of Thermoproteus Spherical Piliferous Virus 1: A Spherical Archaeal Virus Decorated with Unusual Filaments. J. Virol. 2020, 94, e00036-20. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-G.; Kim, S.-J.; Cvirkaite-Krupovic, V.; Yu, W.-J.; Gwak, J.-H.; López-Pérez, M.; Rodriguez-Valera, F.; Krupovic, M.; Cho, J.-C.; Rhee, S.-K. Spindle-shaped viruses infect marine ammonia-oxidizing thaumarchaea. Proc. Natl. Acad. Sci. USA 2019, 116, 15645–15650. [Google Scholar] [CrossRef] [PubMed]
- Larson, E.T.; Reiter, D.; Young, M.; Lawrence, C.M. Structure of A197 from Sulfolobus Turreted Icosahedral Virus: A Crenarchaeal Viral Glycosyltransferase Exhibiting the GT-A Fold. J. Virol. 2006, 80, 7636–7644. [Google Scholar] [CrossRef]
- Bath, C.; Cukalac, T.; Porter, K.; Dyall-Smith, M.L. His1 and His2 are distantly related, spindle-shaped haloviruses belonging to the novel virus group, Salterprovirus. Virology 2006, 350, 228–239. [Google Scholar] [CrossRef]
- Iranzo, J.; Koonin Eugene, V.; Prangishvili, D.; Krupovic, M. Bipartite Network Analysis of the Archaeal Virosphere: Evolutionary Connections between Viruses and Capsidless Mobile Elements. J. Virol. 2016, 90, 11043–11055. [Google Scholar] [CrossRef] [PubMed]
- Munson-McGee, J.H.; Rooney, C.; Young, M.J. An Uncultivated Virus Infecting a Nanoarchaeal Parasite in the Hot Springs of Yellowstone National Park. J. Virol. 2020, 94, e01213-19. [Google Scholar] [CrossRef]
- Rambo, I.M.; Langwig, M.V.; Leão, P.; De Anda, V.; Baker, B.J. Genomes of six viruses that infect Asgard archaea from deep-sea sediments. Nat. Microbiol. 2022, 7, 953–961. [Google Scholar] [CrossRef]
- Alarcón-Schumacher, T.; Naor, A.; Gophna, U.; Erdmann, S. Isolation of a virus causing a chronic infection in the archaeal model organism Haloferax volcanii reveals antiviral activities of a provirus. Proc. Natl. Acad. Sci. USA 2022, 119, e2205037119. [Google Scholar] [CrossRef] [PubMed]
- Pietilä, M.K.; Roine, E.; Sencilo, A.; Bamford, D.H.; Oksanen, H.M. Pleolipoviridae, a newly proposed family comprising archaeal pleomorphic viruses with single-stranded or double-stranded DNA genomes. Arch. Virol. 2016, 161, 249–256. [Google Scholar] [CrossRef]
- Erdmann, S.; Tschitschko, B.; Zhong, L.; Raftery, M.J.; Cavicchioli, R. A plasmid from an Antarctic haloarchaeon uses specialized membrane vesicles to disseminate and infect plasmid-free cells. Nat. Microbiol. 2017, 2, 1446. [Google Scholar] [CrossRef]
- Forterre, P.; Da Cunha, V.; Catchpole, R. Plasmid vesicles mimicking virions. Nat. Microbiol. 2017, 2, 1340. [Google Scholar] [CrossRef]
- Mills, J.; Gebhard, L.J.; Schubotz, F.; Shevchenko, A.; Speth, D.R.; Liao, Y.; Duggin, I.G.; Marchfelder, A.; Erdmann, S. Extracellular vesicles of Euryarchaeida: Precursor to eukaryotic membrane trafficking. bioRxiv 2023. [Google Scholar] [CrossRef]
- Koonin, E.V.; Dolja, V.V.; Krupovic, M.; Kuhn, J.H. Viruses Defined by the Position of the Virosphere within the Replicator Space. Microbiol. Mol. Biol. Rev. 2021, 85, e00193-20. [Google Scholar] [CrossRef] [PubMed]
- Burns, D.; Dyall-Smith, M. 22 Cultivation of haloarchaea. In Methods in Microbiology; Elsevier: Amsterdam, The Netherlands, 2006; Volume 35, pp. 535–552. [Google Scholar]
- Gebhard, L.J.; Duggin, I.G.; Erdmann, S. Improving the genetic system for Halorubrum lacusprofundi to allow in-frame deletions. Front. Microbiol. 2023, 14, 5621. [Google Scholar] [CrossRef] [PubMed]
- Dyall-Smith, M. The Halohandbook: Protocols for Halobacterial Genetics, Version 7.3. 2009. Available online: https://haloarchaea.com/wp-content/uploads/2018/10/Halohandbook_2009_v7.3mds.pdf (accessed on 1 November 2022).
- Franzmann, P.; Stackebrandt, E.; Sanderson, K.; Volkman, J.; Cameron, D.; Stevenson, P.; McMeekin, T.; Burton, H. Halobacterium lacusprofundi sp. nov., a halophilic bacterium isolated from Deep Lake, Antarctica. Syst. Appl. Microbiol. 1988, 11, 20–27. [Google Scholar] [CrossRef]
- Allers, T.; Ngo, H.-P.; Mevarech, M.; Lloyd, R.G. Development of Additional Selectable Markers for the Halophilic Archaeon Haloferax volcanii Based on the leuB and trpA Genes. Appl. Environ. Microbiol. 2004, 70, 943–953. [Google Scholar] [CrossRef]
- Snedecor, G.; Cochran, W. Statistical Methods, 7th ed.; The Iowa State University: Ames, IA, USA, 1989. [Google Scholar]
- Levene, H. Robust tests for equality of variances. In Contributions to Probability and Statistics; Olkin, I., Ed.; Stanford University Press: Palo Alto, CA, USA, 1960; pp. 278–292. [Google Scholar]
- Fligner, M.A.; Killeen, T.J. Distribution-Free Two-Sample Tests for Scale. J. Am. Stat. Assoc. 1976, 71, 210–213. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing, Version 4.1.2; R Foundation for Statistical Computing: Vienna, Austria, 2021.
- RStudio TeamRStudio: Integrated Development Environment for R, Version 2023.3.0.386; Posit Software, PBC: Boston, MA, USA, 2023. Available online: http://www.rstudio.com/ (accessed on 1 November 2022).
- Dubray, G.; Bezard, G. A highly sensitive periodic acid-silver stain for 1,2-diol groups of glycoproteins and polysaccharides in polyacrylamide gels. Anal. Biochem. 1982, 119, 325–329. [Google Scholar] [CrossRef]
- Deatherage, B.L.; Cookson, B.T. Membrane vesicle release in bacteria, eukaryotes, and archaea: A conserved yet underappreciated aspect of microbial life. Infect. Immun. 2012, 80, 1948–1957. [Google Scholar] [CrossRef]
- Schatz, D.; Vardi, A. Extracellular vesicles—New players in cell–cell communication in aquatic environments. Curr. Opin. Microbiol. 2018, 43, 148–154. [Google Scholar] [CrossRef]
- Gill, S.; Catchpole, R.; Forterre, P. Extracellular membrane vesicles in the three domains of life and beyond. FEMS Microbiol. Rev. 2019, 43, 273–303. [Google Scholar] [CrossRef] [PubMed]
- Gavel, Y.; von Heijne, G. Sequence differences between glycosylated and non-glycosylated Asn-X-Thr/Ser acceptor sites: Implications for protein engineering. Protein Eng. 1990, 3, 433–442. [Google Scholar] [CrossRef]
- Mäntynen, S.; Laanto, E.; Oksanen, H.M.; Poranen, M.M.; Díaz-Muñoz, S.L. Black box of phage–bacterium interactions: Exploring alternative phage infection strategies. Open Biol. 2021, 11, 210188. [Google Scholar] [CrossRef] [PubMed]
- Zaretsky, M.; Darnell, C.L.; Schmid, A.K.; Eichler, J. N-glycosylation Is Important for Halobacterium salinarum Archaellin Expression, Archaellum Assembly and Cell Motility. Front. Microbiol. 2019, 10, 1367. [Google Scholar] [CrossRef] [PubMed]
- Chaban, B.; Voisin, S.; Kelly, J.; Logan, S.M.; Jarrell, K.F. Identification of genes involved in the biosynthesis and attachment of Methanococcus voltae N-linked glycans: Insight into N-linked glycosylation pathways in Archaea. Mol. Microbiol. 2006, 61, 259–268. [Google Scholar] [CrossRef]
- Guan, Z.; Naparstek, S.; Calo, D.; Eichler, J. Protein glycosylation as an adaptive response in Archaea: Growth at different salt concentrations leads to alterations in Haloferax volcanii S-layer glycoprotein N-glycosylation. Environ. Microbiol. 2012, 14, 743–753. [Google Scholar] [CrossRef]
- Kaminski, L.; Guan, Z.; Yurist-Doutsch, S.; Eichler, J. Two Distinct N-glycosylation Pathways Process the Haloferax volcanii S-Layer Glycoprotein upon Changes in Environmental Salinity. mBio 2013, 4, e00716-13. [Google Scholar] [CrossRef]
- Mescher, M.F.; Strominger, J.L. Purification and characterization of a prokaryotic glycoprotein from the cell envelope of Halobacterium salinarium. J. Biol. Chem. 1976, 251, 2005–2014. [Google Scholar] [CrossRef]
- Sumper, M.; Berg, E.; Mengele, R.; Strobel, I. Primary structure and glycosylation of the S-layer protein of Haloferax volcanii. J. Bacteriol. 1990, 172, 7111–7118. [Google Scholar] [CrossRef]
- Lu, H.; Pei, C.; Zhou, H.; Lü, Y.; He, Y.; Li, Y.; Han, J.; Xiang, H.; Eichler, J.; Jin, C. Agl22 and Agl23 are involved in the synthesis and utilization of the lipid-linked intermediates in the glycosylation pathways of the halophilic archaeaon Haloarcula hispanica. Mol. Microbiol. 2020, 114, 762–774. [Google Scholar] [CrossRef]
- Kelly, J.; Vinogradov, E.; Robotham, A.; Tessier, L.; Logan, S.M.; Jarrell, K.F. Characterizing the N- and O-linked glycans of the PGF-CTERM sorting domain-containing S-layer protein of Methanoculleus marisnigri. Glycobiology 2022, 32, 629–644. [Google Scholar] [CrossRef] [PubMed]
- Tschitschko, B.; Williams, T.J.; Allen, M.A.; Páez-Espino, D.; Kyrpides, N.; Zhong, L.; Raftery, M.J.; Cavicchioli, R. Antarctic archaea–virus interactions: Metaproteome-led analysis of invasion, evasion and adaptation. ISME J. 2015, 9, 2094–2107. [Google Scholar] [CrossRef] [PubMed]
- Legrand, P.; Collins, B.; Blangy, S.; Murphy, J.; Spinelli, S.; Gutierrez, C.; Richet, N.; Kellenberger, C.; Desmyter, A.; Mahony, J.; et al. The Atomic Structure of the Phage Tuc2009 Baseplate Tripod Suggests that Host Recognition Involves Two Different Carbohydrate Binding Modules. mBio 2016, 7, e01781-15. [Google Scholar] [CrossRef]
- Hayes, S.; Vincentelli, R.; Mahony, J.; Nauta, A.; Ramond, L.; Lugli, G.A.; Ventura, M.; van Sinderen, D.; Cambillau, C. Functional carbohydrate binding modules identified in evolved dits from siphophages infecting various Gram-positive bacteria. Mol. Microbiol. 2018, 110, 777–795. [Google Scholar] [CrossRef] [PubMed]
- Lavelle, K.; Goulet, A.; McDonnell, B.; Spinelli, S.; van Sinderen, D.; Mahony, J.; Cambillau, C. Revisiting the host adhesion determinants of Streptococcus thermophilus siphophages. Microb. Biotechnol. 2020, 13, 1765–1779. [Google Scholar] [CrossRef] [PubMed]
- Beamud, B.; García-González, N.; Gómez-Ortega, M.; González-Candelas, F.; Domingo-Calap, P.; Sanjuan, R. Genetic determinants of host tropism in Klebsiella phages. Cell Rep. 2023, 42, 112048. [Google Scholar] [CrossRef]
- Atanasova, N.S.; Roine, E.; Oren, A.; Bamford, D.H.; Oksanen, H.M. Global network of specific virus–host interactions in hypersaline environments. Environ. Microbiol. 2012, 14, 426–440. [Google Scholar] [CrossRef]
- Atanasova, N.S.; Demina, T.A.; Buivydas, A.; Bamford, D.H.; Oksanen, H.M. Archaeal viruses multiply: Temporal screening in a solar saltern. Viruses 2015, 7, 1902–1926. [Google Scholar] [CrossRef]
- Mizuno, C.M.; Prajapati, B.; Lucas-Staat, S.; Sime-Ngando, T.; Forterre, P.; Bamford, D.H.; Prangishvili, D.; Krupovic, M.; Oksanen, H.M. Novel haloarchaeal viruses from Lake Retba infecting Haloferax and Halorubrum species. Environ. Microbiol. 2019, 21, 2129–2147. [Google Scholar] [CrossRef]
- Quemin, E.R.J.; Quax, T.E.F. Archaeal viruses at the cell envelope: Entry and egress. Front. Microbiol. 2015, 6, 552. [Google Scholar] [CrossRef]
- Konrad, Z.; Eichler, J. Lipid modification of proteins in Archaea: Attachment of a mevalonic acid-based lipid moiety to the surface-layer glycoprotein of Haloferax volcanii follows protein translocation. Biochem. J. 2002, 366, 959–964. [Google Scholar] [CrossRef] [PubMed]
- Abdul Halim, M.F.; Pfeiffer, F.; Zou, J.; Frisch, A.; Haft, D.; Wu, S.; Tolić, N.; Brewer, H.; Payne, S.H.; Paša-Tolić, L.; et al. Haloferax volcanii archaeosortase is required for motility, mating, and C-terminal processing of the S-layer glycoprotein. Mol. Microbiol. 2013, 88, 1164–1175. [Google Scholar] [CrossRef] [PubMed]
- Kandiba, L.; Guan, Z.; Eichler, J. Lipid modification gives rise to two distinct Haloferax volcanii S-layer glycoprotein populations. Biochim. Biophys. Acta 2013, 1828, 938–943. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, M.; Malla, S.; Blythe, M.J.; Nieduszynski, C.A.; Allers, T. Accelerated growth in the absence of DNA replication origins. Nature 2013, 503, 544–547. [Google Scholar] [CrossRef]
- Liu, X.; Miao, D.; Zhang, F.; Wu, Z.; Liu, J.; Xiang, H. Characterization of the minimal replicon of pHM300 and independent copy number control of major and minor chromosomes of Haloferax mediterranei. FEMS Microbiol. Lett. 2013, 339, 66–74. [Google Scholar] [CrossRef]
- Chen, S.; Wang, C.; Xiang, H. Sequence analysis and minimal replicon determination of a new haloarchaeal plasmid pHF2 isolated from Haloferax sp. strain Q22. Plasmid 2016, 83, 1–7. [Google Scholar] [CrossRef]
- Charlebois, R.L.; Lam, W.L.; Cline, S.W.; Doolittle, W.F. Characterization of pHV2 from Halobacterium volcanii and its use in demonstrating transformation of an archaebacterium. Proc. Natl. Acad. Sci. USA 1987, 84, 8530–8534. [Google Scholar] [CrossRef]
- Schleper, C.; Holz, I.; Janekovic, D.; Murphy, J.; Zillig, W. A multicopy plasmid of the extremely thermophilic archaeon Sulfolobus effects its transfer to recipients by mating. J. Bacteriol. 1995, 177, 4417–4426. [Google Scholar] [CrossRef]
- Prangishvili, D.; Albers, S.-V.; Holz, I.; Arnold, H.P.; Stedman, K.; Klein, T.; Singh, H.; Hiort, J.; Schweier, A.; Kristjansson, J.K.; et al. Conjugation in Archaea: Frequent Occurrence of Conjugative Plasmids in Sulfolobus. Plasmid 1998, 40, 190–202. [Google Scholar] [CrossRef]
- Erauso, G.; Stedman, K.M.; van de Werken, H.J.G.; Zillig, W.; van der Oost, J. Two novel conjugative plasmids from a single strain of Sulfolobus. Microbiology 2006, 152, 1951–1968. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HRTV-DL1 | HFPV-1 | PVs (pR1SE) | |
|---|---|---|---|
| Genome size | 37.7 kb | 8 kb | 37.8 kb (pR1SEDL18 = 109 kb) |
| Particle | head-tailed virus, non-contractile tail | pleomorphic, membrane enveloped | undescribed morphology, membrane enveloped |
| Life cycle | lytic | chronic | chronic |
| Predicted glycosylated virus/plasmid proteins | ORF12 (major capsid protein) ORF24 (host attachment) [26] | ORF4 (spike protein) [39] | ORF6 (structural protein), ORF9 (structural protein) [41] |
| S-layer interactions | binds S-layer as primary receptor [26] | not known | not known |
| Glycosylation-related genes on genome | ORF26 (predicted glycosyl hydrolase [26]) | none | ORF89 (predicted glycosyltransferase/glycogen phosphorylase), ORF90 (predicted glycosyltransferase) [41] from plasmid pR1SEDL18 |
| Effect of host N-glycosylation on life cycle of infectious agent | not significant | significant increase in HFPV-1 extracellular titers and intracellular virus:host genome ratios in ∆aglB cells | significant decrease in extracellular pR1SE/PV titers in ∆aglB cells |
| Effect of N-glycosylation of particles on particle stability in varying NaCl concentrations | changes in stability in varying NaCl concentrations are similar to parent strain | particle stability of ∆aglB-produced particles does not increase when comparing low-salt to physiological salt conditions, as observed for particles produced in the parent strain, indicating altered particle stability | not tested |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gebhard, L.J.; Vershinin, Z.; Alarcón-Schumacher, T.; Eichler, J.; Erdmann, S. Influence of N-Glycosylation on Virus–Host Interactions in Halorubrum lacusprofundi. Viruses 2023, 15, 1469. https://doi.org/10.3390/v15071469
Gebhard LJ, Vershinin Z, Alarcón-Schumacher T, Eichler J, Erdmann S. Influence of N-Glycosylation on Virus–Host Interactions in Halorubrum lacusprofundi. Viruses. 2023; 15(7):1469. https://doi.org/10.3390/v15071469
Chicago/Turabian StyleGebhard, L. Johanna, Zlata Vershinin, Tomás Alarcón-Schumacher, Jerry Eichler, and Susanne Erdmann. 2023. "Influence of N-Glycosylation on Virus–Host Interactions in Halorubrum lacusprofundi" Viruses 15, no. 7: 1469. https://doi.org/10.3390/v15071469
APA StyleGebhard, L. J., Vershinin, Z., Alarcón-Schumacher, T., Eichler, J., & Erdmann, S. (2023). Influence of N-Glycosylation on Virus–Host Interactions in Halorubrum lacusprofundi. Viruses, 15(7), 1469. https://doi.org/10.3390/v15071469