

Transcriptomic Responses Underlying the High Virulence of Black Queen Cell Virus and Sacbrood Virus following a Change in Their Mode of Transmission in Honey Bees (Apis mellifera)


Abstract
1. Introduction
2. Materials and Methods
2.1. Honey Bees
2.2. Pesticide
2.3. Viral Inoculum and Infection
2.4. Exposure to FPF Insecticide and Virus
2.5. RNA Extraction and Sequencing
2.6. RNA-Seq Data Analysis
Differential Gene Expression Analysis
2.7. Statistical Analysis
3. Results
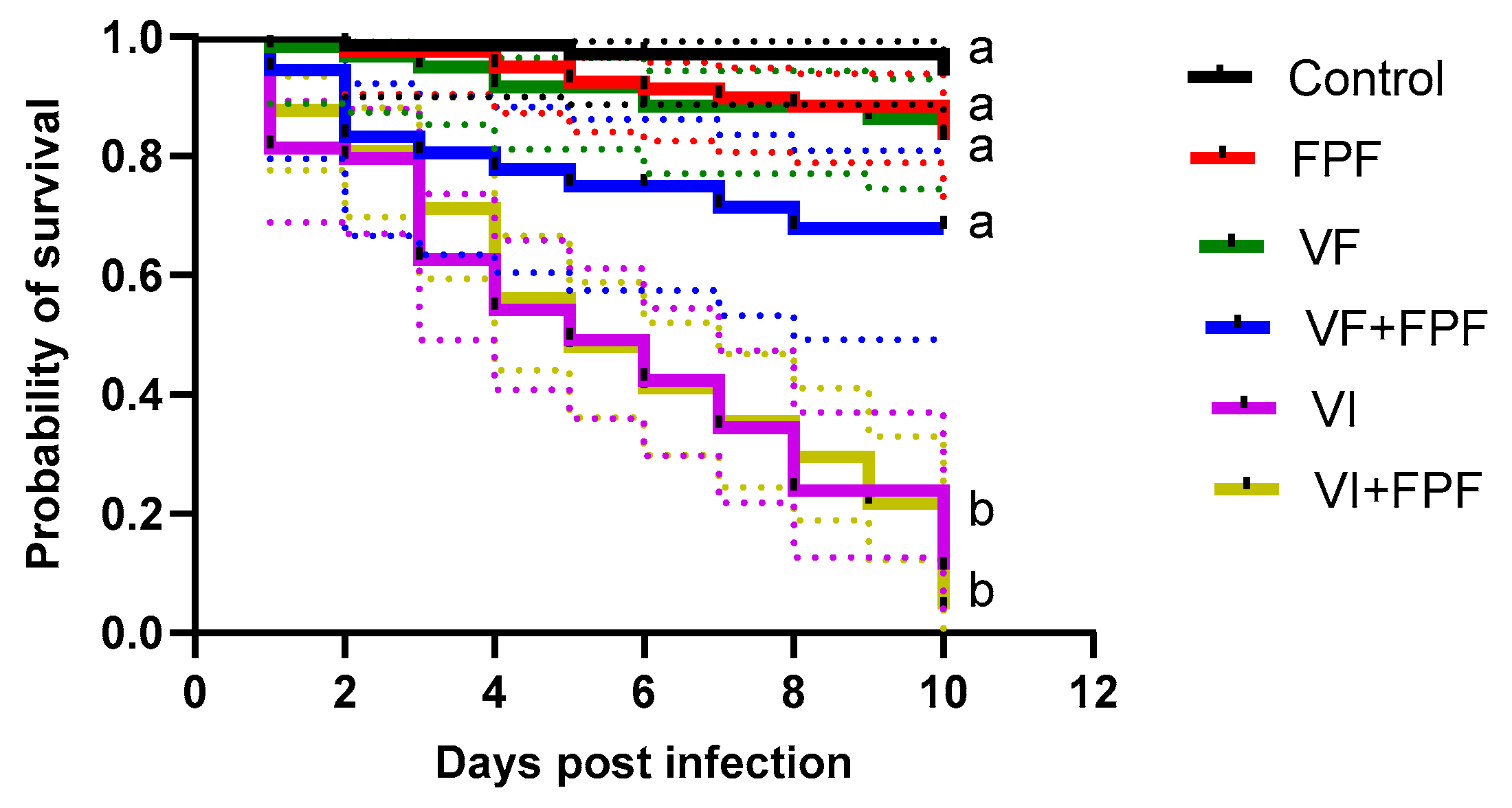
3.1. Effects on Survival
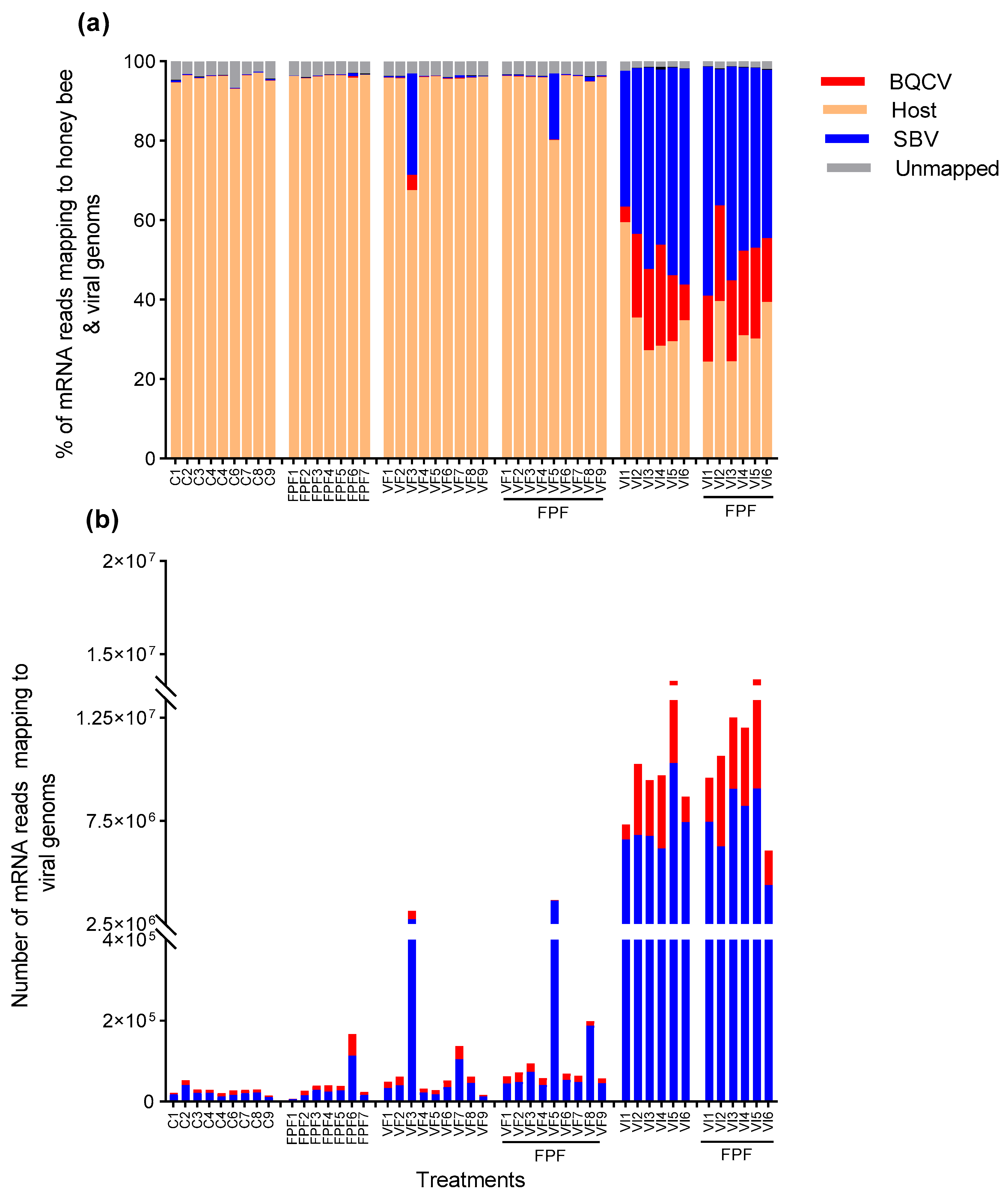
3.2. RNA Sequencing and Mapping Rates
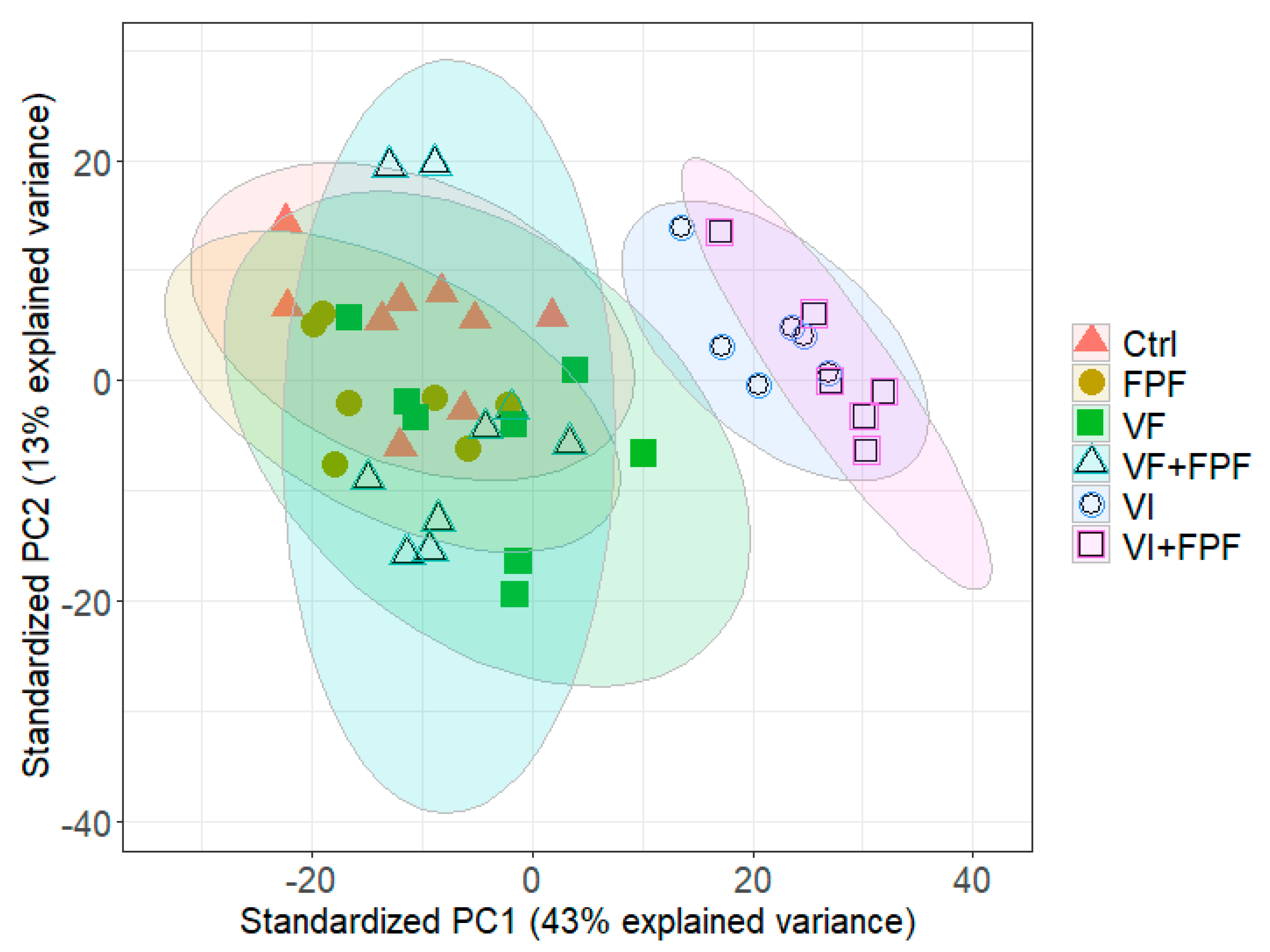
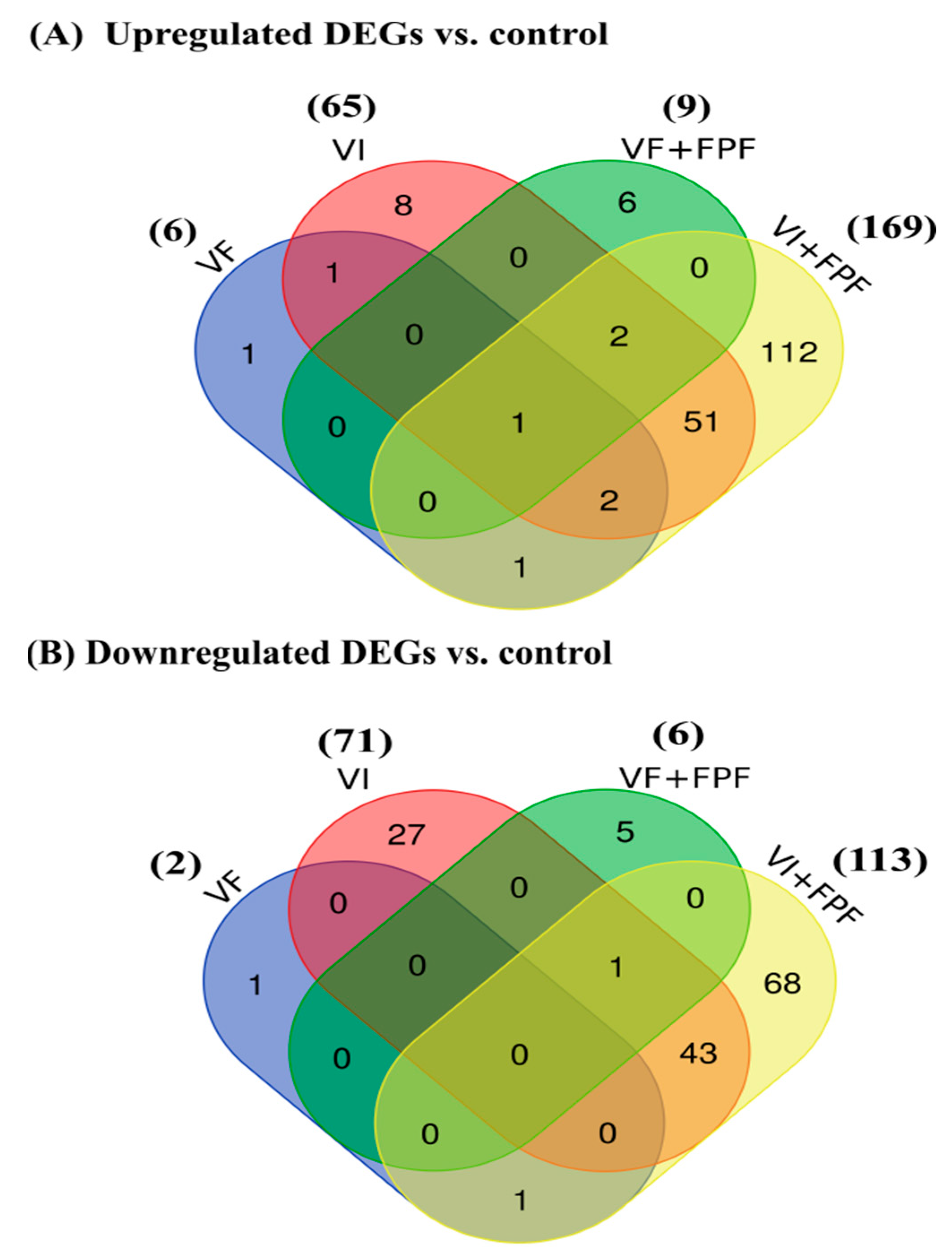
3.3. Differentially Expressed Genes (DEGs)
3.4. Functional Annotation and Classification of the DEGs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khalifa, S.A.M.; Elshafiey, E.H.; Shetaia, A.A.; El-Wahed, A.A.A.; Algethami, A.F.; Musharraf, S.G.; AlAjmi, M.F.; Zhao, C.; Masry, S.H.D.; Abdel-Daim, M.M.; et al. Overview of Bee Pollination and Its Economic Value for Crop Production. Insects 2021, 12, 688. [Google Scholar] [CrossRef] [PubMed]
- Potts, S.G.; Imperatriz-fonseca, V.; Ngo, H.T.; Aizen, M.A.; Biesmeijer, J.C.; Breeze, T.D. Safeguarding Pollinators and Their Values to Human Well-Being. Nature 2016, 540, 220–229. [Google Scholar] [CrossRef] [PubMed]
- Al Naggar, Y.; Codling, G.; Giesy, J.P.; Safer, A. Beekeeping and the Need for Pollination from an Agricultural Perspective in Egypt. Bee World 2018, 95, 107–112. [Google Scholar] [CrossRef]
- Osterman, J.; Aizen, M.A.; Biesmeijer, J.C.; Bosch, J.; Howlett, B.G.; Inouye, D.W.; Jung, C.; Martins, D.J.; Medel, R.; Pauw, A.; et al. Global Trends in the Number and Diversity of Managed Pollinator Species. Agric. Ecosyst. Environ. 2021, 322, 107653. [Google Scholar] [CrossRef]
- van der Zee, R.; Pisa, L.; Andonov, S.; Brodschneider, R.; Charrière, J.-D.; Chlebo, R.; Coffey, M.F.; Crailsheim, K.; Dahle, B.; Gajda, A.; et al. Managed Honey Bee Colony Losses in Canada, China, Europe, Israel and Turkey, for the Winters of 2008–9 and 2009–10. J. Apic. Res. 2012, 51, 100–114. [Google Scholar] [CrossRef]
- Bruckner, S.; Wilson, M.; Aurell, D.; Rennich, K.; VanEngelsdorp, D.; Steinhauer, N.; Williams, G.R. A National Survey of Managed Honey Bee Colony Losses in the USA: Results from the Bee Informed Partnership for 2017–18, 2018–2019, and 2019–20. J. Apic. Res. 2023, 62, 429–443. [Google Scholar] [CrossRef]
- El-Seedi, H.R.; Ahmed, H.R.; El-Wahed, A.A.A.; Saeed, A.; Algethami, A.F.; Attia, N.F.; Guo, Z.; Musharraf, S.G.; Khatib, A.; Alsharif, S.M.; et al. Bee Stressors from an Immunological Perspective and Strategies to Improve Bee Health. Vet. Sci. 2022, 9, 199. [Google Scholar] [CrossRef]
- Hristov, P.; Shumkova, R.; Palova, N.; Neov, B. Factors Associated with Honey Bee Colony Losses: A Mini-Review. Vet. Sci. 2020, 7, 166. [Google Scholar] [CrossRef]
- Insolia, L.; Molinari, R.; Rogers, S.R.; Williams, G.R.; Chiaromonte, F.; Calovi, M. Honey Bee Colony Loss Linked to Parasites, Pesticides and Extreme Weather across the United States. Sci. Rep. 2022, 12, 20787. [Google Scholar] [CrossRef]
- Beaurepaire, A.; Piot, N.; Doublet, V.; Antunez, K.; Campbell, E.; Chantawannakul, P.; Chejanovsky, N.; Gajda, A.; Heerman, M.; Panziera, D.; et al. Diversity and Global Distribution of Viruses of the Western Honey Bee, Apis mellifera. Insects 2020, 11, 239. [Google Scholar] [CrossRef]
- Chen, Y.; Evans, J.; Feldlaufer, M. Horizontal and Vertical Transmission of Viruses in the Honey Bee, Apis mellifera. J. Invertebr. Pathol. 2006, 92, 152–159. [Google Scholar] [CrossRef]
- Ryabov, E.V.; Wood, G.R.; Fannon, J.M.; Moore, J.D.; Bull, J.C.; Chandler, D.; Mead, A.; Burroughs, N.; Evans, D.J. A Virulent Strain of Deformed Wing Virus (DWV) of Honeybees (Apis mellifera) Prevails after Varroa destructor-Mediated, or In Vitro, Transmission. PLoS Pathog. 2014, 10, e1004230. [Google Scholar] [CrossRef]
- Martin, S.J.; Highfield, A.C.; Brettell, L.; Villalobos, E.M.; Budge, G.E.; Powell, M.; Nikaido, S.; Schroeder, D.C. Global Honey Bee Viral Landscape Altered by a Parasitic Mite. Science 2012, 336, 1304–1306. [Google Scholar] [CrossRef]
- Liu, T.P. Virus-like Particles in the Tracheal Mite Acarapis Woodi (Rennie). Apidologie 1991, 22, 213–219. [Google Scholar] [CrossRef]
- Cressler, C.E.; Mcleod, D.V.; Rozins, C.; van den Hoogen, J.; Day, T. The Adaptive Evolution of Virulence: A Review of Theoretical Predictions and Empirical Tests. Parasitology 2016, 143, 915–930. [Google Scholar] [CrossRef]
- Ewald, P.W. Host-Parasite Relations, Vectors, and the Evolution of Disease Severity. Annu. Rev. Ecol. Syst. 1983, 14, 465–485. [Google Scholar] [CrossRef]
- Evans, J.D.; Spivak, M. Socialized Medicine: Individual and Communal Disease Barriers in Honey Bees. J. Invertebr. Pathol. 2010, 103, S62–S72. [Google Scholar] [CrossRef]
- Le Clec’h, W.; Dittmer, J.; Raimond, M.; Bouchon, D.; Sicard, M. Phenotypic Shift in Wolbachia Virulence towards Its Native Host across Serial Horizontal Passages. Proc. R. Soc. B Biol. Sci. 2017, 284, 20171076. [Google Scholar] [CrossRef]
- Al Naggar, Y.; Paxton, R.J. Mode of Transmission Determines the Virulence of Black Queen Cell Virus in Adult Honey Bees, Posing a Future Threat to Bees and Apiculture. Viruses 2020, 12, 535. [Google Scholar] [CrossRef]
- Remnant, E.J.; Mather, N.; Gillard, T.L.; Yagound, B.; Beekman, M. Direct Transmission by Injection Affects Competition among RNA Viruses in Honeybees. Proc. R. Soc. B Biol. Sci. 2019, 286, 20182452. [Google Scholar] [CrossRef]
- Chen, G.; Wu, Y.; Deng, J.; Wen, Z.; Wang, S.; Chen, Y.; Hu, F.; Zheng, H. Seasonal Variation of Viral Infections between the Eastern Honey Bee (Apis cerana) and the Western Honey Bee (Apis mellifera). Microbiologyopen 2021, 10, e1162. [Google Scholar] [CrossRef] [PubMed]
- D’Alvise, P.; Seeburger, V.; Gihring, K.; Kieboom, M.; Hasselmann, M. Seasonal Dynamics and Co-occurrence Patterns of Honey Bee Pathogens Revealed by High-throughput RT-qPCR Analysis. Ecol. Evol. 2019, 9, 10241–10252. [Google Scholar] [CrossRef] [PubMed]
- Welch, A.; Drummond, F.; Tewari, S.; Averill, A.; Burand, J.P. Presence and Prevalence of Viruses in Local and Migratory Honeybees (Apis mellifera) in Massachusetts. Appl. Environ. Microbiol. 2009, 75, 7862–7865. [Google Scholar] [CrossRef] [PubMed]
- Kandel, M.; Paxton, R.J.; Al Naggar, Y. Nationwide Screening for Bee Viruses in Apis Mellifera Colonies in Egypt. Insects 2023, 14, 172. [Google Scholar] [CrossRef]
- Allen, M.; Ball, B. The Incidence and World Distribution of Honey Bee Viruses. Bee World 1996, 77, 141–162. [Google Scholar] [CrossRef]
- Wei, R.; Cao, L.; Feng, Y.; Chen, Y.; Chen, G.; Zheng, H. Sacbrood Virus: A Growing Threat to Honeybees and Wild Pollinators. Viruses 2022, 14, 1871. [Google Scholar] [CrossRef]
- Chen, Y.P.; Siede, R. Honey Bee Viruses. Adv. Virus Res. 2007, 70, 33–80. [Google Scholar] [CrossRef]
- Prodělalová, J.; Moutelíková, R.; Titěra, D. Multiple Virus Infections in Western Honeybee (Apis mellifera L.) Ejaculate Used for Instrumental Insemination. Viruses 2019, 11, 306. [Google Scholar] [CrossRef]
- Phokasem, P.; Liuhao, W.; Panjad, P.; Yujie, T.; Li, J.; Chantawannakul, P. Differential Viral Distribution Patterns in Reproductive Tissues of Apis mellifera and Apis cerana Drones. Front. Vet. Sci. 2021, 8, 608700. [Google Scholar] [CrossRef]
- Mondet, F.; de Miranda, J.R.; Kretzschmar, A.; Le Conte, Y.; Mercer, A.R. On the Front Line: Quantitative Virus Dynamics in Honeybee (Apis mellifera L.) Colonies along a New Expansion Front of the Parasite Varroa Destructor. PLoS Pathog. 2014, 10, e1004323. [Google Scholar] [CrossRef]
- Chantawannakul, P.; Ward, L.; Boonham, N.; Brown, M. A Scientific Note on the Detection of Honeybee Viruses Using Real-Time PCR (TaqMan) in Varroa Mites Collected from a Thai Honeybee (Apis mellifera) Apiary. J. Invertebr. Pathol. 2006, 91, 69–73. [Google Scholar] [CrossRef]
- Shen, M.; Cui, L.; Ostiguy, N.; Cox-Foster, D. Intricate Transmission Routes and Interactions between Picorna-like Viruses (Kashmir Bee Virus and Sacbrood Virus) with the Honeybee Host and the Parasitic Varroa Mite. J. Gen. Virol. 2005, 86, 2281–2289. [Google Scholar] [CrossRef]
- Wang, S.; Chen, G.; Lin, Z.; Wu, Y.; Hu, F.; Zheng, H. Occurrence of Multiple Honeybee Viruses in the Ectoparasitic Mites Varroa Spp. in Apis Cerana Colonies. J. Invertebr. Pathol. 2019, 166, 107225. [Google Scholar] [CrossRef]
- Al Naggar, Y.; Paxton, R.J. The Novel Insecticides Flupyradifurone and Sulfoxaflor Do Not Act Synergistically with Viral Pathogens in Reducing Honey Bee (Apis mellifera) Survival but Sulfoxaflor Modulates Host Immunocompetence. Microb. Biotechnol. 2021, 14, 227–240. [Google Scholar] [CrossRef]
- Poquet, Y.; Vidau, C.; Alaux, C. Modulation of Pesticide Response in Honeybees. Apidologie 2016, 47, 412–426. [Google Scholar] [CrossRef]
- Al Naggar, Y.; Baer, B. Consequences of a Short Time Exposure to a Sublethal Dose of Flupyradifurone (Sivanto) Pesticide Early in Life on Survival and Immunity in the Honeybee (Apis mellifera). Sci. Rep. 2019, 9, 19753. [Google Scholar] [CrossRef]
- Nauen, R.; Jeschke, P.; Velten, R.; Beck, M.E.; Ebbinghaus-Kintscher, U.; Thielert, W.; Wölfel, K.; Haas, M.; Kunz, K.; Raupach, G. Flupyradifurone: A Brief Profile of a New Butenolide Insecticide. Pest Manag. Sci. 2015, 71, 850–862. [Google Scholar] [CrossRef]
- Siviter, H.; Muth, F. Do Novel Insecticides Pose a Threat to Beneficial Insects? Proc. R. Soc. B Biol. Sci. 2020, 287, 20201265. [Google Scholar] [CrossRef]
- Al Naggar, Y.; Singavarapu, B.; Paxton, R.J.; Wubet, T. Bees under Interactive Stressors: The Novel Insecticides Flupyradifurone and Sulfoxaflor along with the Fungicide Azoxystrobin Disrupt the Gut Microbiota of Honey Bees and Increase Opportunistic Bacterial Pathogens. Sci. Total Environ. 2022, 849, 157941. [Google Scholar] [CrossRef]
- Tehel, A.; Vu, Q.; Bigot, D.; Gogol-döring, A.; Koch, P.; Jenkins, C.; Doublet, V.; Theodorou, P.; Paxton, R. The Two Prevalent Genotypes of an Emerging Infectious Disease, Deformed Wing Virus, Cause Equally low Pupal Mortality and Equally High Wing Deformities in Host Honey Bees. Viruses 2019, 11, 114. [Google Scholar] [CrossRef]
- De Miranda, J.R.; Bailey, L.; Ball, B.V.; Blanchard, P.; Budge, G.E.; Chejanovsky, N.; Chen, Y.-P.; Gauthier, L.; Genersch, E.; de Graaf, D.C.; et al. Standard Methods for Virus Research in Apis mellifera. J. Apic. Res. 2013, 52, 1–56. [Google Scholar] [CrossRef]
- U.S. EPA. Environmental Fate and Ecological Risk Assessment for Foliar, Soil Drench, and Seed Treatment Uses of the New Insecticide Flupyradifurone (BYI 02960). 2014. Available online: https://www.regulations.gov/document/EPA-HQ-OPP-2013-0226-0047 (accessed on 25 May 2023).
- McMahon, D.P.; Fürst, M.A.; Caspar, J.; Theodorou, P.; Brown, M.J.F.; Paxton, R.J. A Sting in the Spit: Widespread Cross-Infection of Multiple RNA Viruses across Wild and Managed Bees. J. Anim. Ecol. 2015, 84, 615–624. [Google Scholar] [CrossRef] [PubMed]
- Locke, B.; Conte, Y.L.; Crauser, D.; Fries, I. Host Adaptations Reduce the Reproductive Success of Varroa Destructor in Two Distinct European Honey Bee Populations. Ecol. Evol. 2012, 2, 1144–1150. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An Ultra-Fast All-in-One FASTQ Preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Bray, N.L.; Pimentel, H.; Melsted, P.; Pachter, L. Near-Optimal Probabilistic RNA-Seq Quantification. Nat. Biotechnol. 2016, 34, 525–527. [Google Scholar] [CrossRef]
- Soneson, C.; Love, M.I.; Robinson, M.D. Differential Analyses for RNA-Seq: Transcript-Level Estimates Improve Gene-Level Inferences. F1000Research 2016, 4, 1521. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. 2020. Available online: http://www.R-project.org/ (accessed on 25 May 2023).
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Vu, V.Q. Ggbiplot: A Ggplot2 Based Biplot (R Package Version 0.55). 2011. Available online: https://rdocumentation.org/packages/ggbiplot/versions/0.55 (accessed on 25 May 2023).
- Raudvere, U.; Kolberg, L.; Kuzmin, I.; Arak, T.; Adler, P.; Peterson, H.; Vilo, J. G:Profiler: A Web Server for Functional Enrichment Analysis and Conversions of Gene Lists (2019 Update). Nucleic Acids Res. 2019, 47, W191–W198. [Google Scholar] [CrossRef]
- Therneau, T. Coxme. R Package Version 2.2-5. 2012. Available online: http://cran.r-project.org/web/packages/coxme (accessed on 25 May 2023).
- Hothorn, T.; Bretz, F.; Westfal, P.; Heiberger, R.M.; Schuetzenmeister, A. Multcomp: Simultaneous Inference in General Parametric Models. 2013. Available online: http://multcomp.R-forge.R-project.org (accessed on 25 May 2023).
- Bailey, L.; Ball, B.V.; Perry, J.N. Association of Viruses with Two Protozoal Pathogens of the Honey Bee. Ann. Appl. Biol. 1983, 103, 13–20. [Google Scholar] [CrossRef]
- Doublet, V.; Labarussias, M.; de Miranda, J.R.; Moritz, R.F.A.; Paxton, R.J. Bees under Stress: Sublethal Doses of a Neonicotinoid Pesticide and Pathogens Interact to Elevate Honey Bee Mortality across the Life Cycle. Environ. Microbiol. 2015, 17, 969–983. [Google Scholar] [CrossRef]
- Holden, H.M.; Rayment, I.; Thoden, J.B. Structure and Function of Enzymes of the Leloir Pathway for Galactose Metabolism. J. Biol. Chem. 2003, 278, 43885–43888. [Google Scholar] [CrossRef]
- Delnoy, B.; Coelho, A.I.; Rubio-Gozalbo, M.E. Current and Future Treatments for Classic Galactosemia. J. Pers. Med. 2021, 11, 75. [Google Scholar] [CrossRef]
- Chen, P.; Lu, Y.-H.; Lin, Y.-H.; Wu, C.-P.; Tang, C.-K.; Wei, S.-C.; Wu, Y.-L. Deformed Wing Virus Infection Affects the Neurological Function of Apis mellifera by Altering Extracellular Adenosine Signaling. Insect Biochem. Mol. Biol. 2021, 139, 103674. [Google Scholar] [CrossRef]
- Merkling, S.H.; van Rij, R.P. Beyond RNAi: Antiviral Defense Strategies in Drosophila and Mosquito. J. Insect Physiol. 2013, 59, 159–170. [Google Scholar] [CrossRef]
- Grozinger, C.M.; Flenniken, M.L. Bee Viruses: Ecology, Pathogenicity, and Impacts. Ann. Rev. Entomol. 2019, 64, 205–226. [Google Scholar] [CrossRef]
- Brutscher, L.M.; McMenamin, A.J.; Flenniken, M.L. The Buzz about Honey Bee Viruses. PLoS Pathog. 2016, 12, e1005757. [Google Scholar] [CrossRef]
- Ryabov, E.V.; Fannon, J.M.; Moore, J.D.; Wood, G.R.; Evans, D.J. The Iflaviruses Sacbrood Virus and Deformed Wing Virus Evoke Different Transcriptional Responses in the Honeybee Which May Facilitate Their Horizontal or Vertical Transmission. PeerJ 2016, 4, e1591. [Google Scholar] [CrossRef]
- Feng, M.; Fei, S.; Xia, J.; Labropoulou, V.; Swevers, L.; Sun, J. Antimicrobial Peptides as Potential Antiviral Factors in Insect Antiviral Immune Response. Front. Immunol. 2020, 11, 2030. [Google Scholar] [CrossRef]
- Pizzorno, M.C.; Field, K.; Kobokovich, A.L.; Martin, P.L.; Gupta, R.A.; Mammone, R.; Rovnyak, D.; Capaldi, E.A. Transcriptomic Responses of the Honey Bee Brain to Infection with Deformed Wing Virus. Viruses 2021, 13, 287. [Google Scholar] [CrossRef]
- Galbraith, D.A.; Yang, X.; Niño, E.L.; Yi, S.; Grozinger, C. Parallel Epigenomic and Transcriptomic Responses to Viral Infection in Honey Bees (Apis mellifera). PLoS Pathog. 2015, 11, e1004713. [Google Scholar] [CrossRef] [PubMed]
- Rutter, L.; Carrillo-Tripp, J.; Bonning, B.C.; Cook, D.; Toth, A.L.; Dolezal, A.G. Transcriptomic Responses to Diet Quality and Viral Infection in Apis mellifera. BMC Genom. 2019, 20, 412. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, Á.G.; Naylor, H.; Esteves, S.S.; Pais, I.S.; Martins, N.E.; Teixeira, L. The Toll-Dorsal Pathway Is Required for Resistance to Viral Oral Infection in Drosophila. PLoS Pathog. 2014, 10, e1004507. [Google Scholar] [CrossRef] [PubMed]
- Brutscher, L.M.; Daughenbaugh, K.F.; Flenniken, M.L. Antiviral Defense Mechanisms in Honey Bees. Curr. Opin. Insect Sci. 2015, 10, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Brito, N.F.; Moreira, M.F.; Melo, A.C.A. A Look inside Odorant-Binding Proteins in Insect Chemoreception. J. Insect Physiol. 2016, 95, 51–65. [Google Scholar] [CrossRef]
- Williamson, S.M.; Baker, D.D.; Wright, G.A. Acute Exposure to a Sublethal Dose of Imidacloprid and Coumaphos Enhances Olfactory Learning and Memory in the Honeybee Apis mellifera. Invertebr. Neurosci. 2013, 13, 63–70. [Google Scholar] [CrossRef]
- Shi, J.; Yang, H.; Yu, L.; Liao, C.; Liu, Y.; Jin, M.; Yan, W.; Wu, X.B. Sublethal Acetamiprid Doses Negatively Affect the Lifespans and Foraging Behaviors of Honey Bee (Apis mellifera L.) Workers. Sci. Total Environ. 2020, 738, 139924. [Google Scholar] [CrossRef]
- Gao, J.; Jin, S.S.; He, Y.; Luo, J.H.; Xu, C.Q.; Wu, Y.Y.; Hou, C.S.; Wang, Q.; Diao, Q.Y. Physiological Analysis and Transcriptome Analysis of Asian Honey Bee (Apis cerana cerana) in Response to Sublethal Neonicotinoid Imidacloprid. Insects 2020, 11, 753. [Google Scholar] [CrossRef]
- Iqbal, J.; Mueller, U. Virus Infection Causes Specific Learning Deficits in Honeybee Foragers. Proc. R. Soc. B Biol. Sci. 2007, 274, 1517–1521. [Google Scholar] [CrossRef]
- Li, Z.; Chen, Y.; Zhang, S.; Chen, S.; Li, W.; Yan, L.; Shi, L.; Wu, L.; Sohr, A.; Su, S. Viral Infection Affects Sucrose Responsiveness and Homing Ability of Forager Honey Bees, Apis mellifera L. PLoS ONE 2013, 8, e77354. [Google Scholar] [CrossRef]
- Dittes, J.; Schäfer, M.O.; Aupperle-Lellbach, H.; Mülling, C.K.W.; Emmerich, I.U. Overt Infection with Chronic Bee Paralysis Virus (CBPV) in Two Honey Bee Colonies. Vet. Sci. 2020, 7, 142. [Google Scholar] [CrossRef]
- Baracchi, D.; Francese, S.; Turillazzi, S. Beyond the Antipredatory Defence: Honey Bee Venom Function as a Component of Social Immunity. Toxicon 2011, 58, 550–557. [Google Scholar] [CrossRef]
- Büyükgüzel, E.; Tunaz, H.; Stanley, D.; Büyükgüzel, K. Eicosanoids Mediate Galleria mellonella Cellular Immune Response to Viral Infection. J. Insect Physiol. 2007, 53, 99–105. [Google Scholar] [CrossRef]
- Stanley, D.; Kim, Y. Insect prostaglandins and other eicosanoids: From molecular to physiological actions. Adv. Insect Physiol. 2019, 56, 283–343. [Google Scholar] [CrossRef]
- Bazzo, R.; Tappin, M.J.; Pastore, A.; Harvey, T.S.; Carver, J.A.; Campbell, I.D. The Structure of Melittin. A 1H-NMR Study in Methanol. Eur. J. Biochem. 1988, 173, 139–146. [Google Scholar] [CrossRef]
- Memariani, H.; Memariani, M.; Moravvej, H.; Shahidi-Dadras, M. Melittin: A Venom-Derived Peptide with Promising Anti-Viral Properties. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 5–17. [Google Scholar] [CrossRef]
- Lee, K.S.; Kim, B.Y.; Yoon, H.J.; Choi, Y.S.; Jin, B.R. Secapin, a Bee Venom Peptide, Exhibits Anti-Fibrinolytic, Anti-Elastolytic, and Anti-Microbial Activities. Dev. Comp. Immunol. 2016, 63, 27–35. [Google Scholar] [CrossRef]
- Yang, J.; Lee, K.S.; Kim, B.Y.; Choi, Y.S.; Yoon, H.J.; Jia, J.; Jin, B.R. Anti-Fibrinolytic and Anti-Microbial Activities of a Serine Protease Inhibitor from Honeybee (Apis Cerana) Venom. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2017, 201, 11–18. [Google Scholar] [CrossRef]
- Argiolas, A.; Herring, P.; Pisano, J.J. Amino Acid Sequence of Bumblebee MCD Peptide: A New Mast Cell Degranulating Peptide from the Venom of the Bumblebee Megabombus pennsylvanicus. Peptides 1985, 6 (Suppl. S3), 431–436. [Google Scholar] [CrossRef]
- Ziai, M.R.; Russek, S.; Wang, H.C.; Beer, B.; Blume, A.J. Mast Cell Degranulating Peptide: A Multi-Functional Neurotoxin. J. Pharm. Pharmacol. 1990, 42, 457–461. [Google Scholar] [CrossRef]
- Barribeau, S.M.; Schmid-Hempel, P.; Sadd, B.M. Royal Decree: Gene Expression in Trans-Generationally Immune Primed Bumblebee Workers Mimics a Primary Immune Response. PLoS ONE 2016, 11, e0159635. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Zhang, Z.; Zhuang, M.; Wang, L.; Li, K.; Yao, J.; Yang, H.; Huang, J.; Hao, Y.; Ying, F.; et al. Transcriptome Profiling Reveals a Novel Mechanism of Antiviral Immunity Upon Sacbrood Virus Infection in Honey Bee Larvae (Apis cerana). Front. Microbiol. 2021, 12, 615893. [Google Scholar] [CrossRef] [PubMed]
- Erban, T.; Sopko, B.; Kadlikova, K.; Talacko, P.; Harant, K. Varroa Destructor Parasitism Has a Greater Effect on Proteome Changes than the Deformed Wing Virus and Activates TGF-β Signaling Pathways. Sci. Rep. 2019, 9, 9400. [Google Scholar] [CrossRef] [PubMed]
- Weaver, D.B.; Cantarel, B.L.; Elsik, C.G.; Boncristiani, D.L.; Evans, J.D. Multi-Tiered Analyses of Honey Bees That Resist or Succumb to Parasitic Mites and Viruses. BMC Genom. 2021, 22, 720. [Google Scholar] [CrossRef]
- Seehuus, S.-C.; Norberg, K.; Gimsa, U.; Krekling, T.; Amdam, G.V. Reproductive Protein Protects Functionally Sterile Honey Bee Workers from Oxidative Stress. Proc. Natl. Acad. Sci. USA 2006, 103, 962–967. [Google Scholar] [CrossRef]
- Amdam, G.V.; Norberg, K.; Hagen, A.; Omholt, S.W. Social Exploitation of Vitellogenin. Proc. Natl. Acad. Sci. USA 2003, 100, 100–1799. [Google Scholar] [CrossRef]
- Amdam, G.V.; Simões, Z.L.; Hagen, A.; Norberg, K.; Schrøder, K.; Mikkelsen, Ø.; Kirkwood, T.B.; Omholt, S.W. Hormonal Control of the Yolk Precursor Vitellogenin Regulates Immune Function and Longevity in Honeybees. Exp. Gerontol. 2004, 39, 767–773. [Google Scholar] [CrossRef]
- Ararso, Z.; Ma, C.; Qi, Y.; Feng, M.; Han, B.; Hu, H.; Meng, L.; Li, J. Proteome Comparisons between Hemolymph of Two Honeybee Strains (Apis mellifera ligustica) Reveal Divergent Molecular Basis in Driving Hemolymph Function and High Royal Jelly Secretion. J. Proteome Res. 2018, 17, 402–419. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | β | SE of β Coefficient (+/−) | Exp. β | Z | p |
|---|---|---|---|---|---|
| FPF | 1.05 | 0.64 | 2.87 | 1.64 | 1.00 |
| VF | 0.91 | 0.68 | 2.49 | 1.34 | 1.00 |
| VF+FPF | 1.95 | 0.67 | 7.03 | 2.91 | 0.05 |
| VI | 3.61 | 0.60 | 37.09 | 5.97 | <0.001 |
| VI+FPF | 3.68 | 0.60 | 39.68 | 6.12 | <0.001 |
| Interaction between viruses and FPF | |||||
| VF+FPF vs. | VF | 1.00 | |||
| FPF | 1.00 | ||||
| VI+FPF vs. | VI | 1.00 | |||
| FPF | <0.001 | ||||
| Gene Name | Gene Description | Category | (log2 Fold Change) | |||
|---|---|---|---|---|---|---|
| VF | VF+FPF | VI | VI+FPF | |||
| LOC113218760 | probable galactose-1-phosphate uridylyltransferase | Carbohydrate metabolism | 5.46 | 5.02 | 6.28 | 6.32 |
| LOC725754 | zinc finger protein castor homolog 1, transcript variant X4 | Cell communication and signaling | 2.15 | - | 2.76 | 2.44 |
| Oa1 | octopamine receptor | Signaling receptor activity | 2.41 | - | - | - |
| Mrjp1 | major royal jelly protein 1 | MRJP family | - | 2.36 | - | - |
| Mrjp5 | major royal jelly protein 5 | MRJP family | - | 3.38 | - | - |
| Mrjp6 | major royal jelly protein 6 | MRJP family | - | 2.60 | - | - |
| Apid1 | apidaecin 1 | Antimicrobial peptides (AMPs) | - | 2.4 | 3.71 | 3.8 |
| LOC406142 | hymenoptaecin | AMPs | - | 2.21 | 2.68 | 2.93 |
| LOC411577 | protein argonaute-2 | RNAi | - | - | 2.72 | 2.6 |
| LOC726766 | endoribonuclease Dicer, transcript variant X1 | RNAi | - | - | 2.17 | 2 |
| Y-e3 | yellow-e3 | MRJP family | - | - | 2.35 | 2.85 |
| CPR1 | cuticular protein 1 | Structural constituent of cuticle | - | - | 2.56 | 3.19 |
| nAChRb2 | nicotinic acetylcholine receptor beta2 subunit | Signaling receptor activity | - | - | 2.83 | 2.56 |
| LOC107964791 | titin-like | Hypertrophic cardiomyopathy (HCM) | - | - | 2.62 | 3.42 |
| Obp9 | odorant binding protein 9 | Odorant receptor | - | - | 3.6 | 4.25 |
| LOC406144 | abaecin | AMPs | - | - | 2.26 | - |
| nAChRa9 | nicotinic acetylcholine receptor alpha9 subunit | Signaling receptor activity | - | - | 2.12 | - |
| LOC100578352 | ionotropic receptor 75a-like | - | - | 3.19 | 3.36 | |
| LOC725629 | DNA-binding protein D-ETS-6-like | - | - | 4.50 | 4.03 | |
| 5-ht7 | serotonin receptor 7 | Signaling receptor activity | - | - | - | 2.42 |
| CPR17 | cuticular protein 17 | Structural constituent of cuticle | - | - | - | 3.12 |
| LOC100576212 | odorant receptor 13a | Odorant receptor | - | - | - | 2.97 |
| LOC410235 | toll-like receptor Tollo | Toll | - | - | - | 2.43 |
| LOC410495 | cytochrome P450 307a1 | Detoxification | - | - | - | 2.69 |
| LOC413908 | cytochrome P450 6A1, transcript variant X1 | Detoxification | - | - | - | 5.71 |
| LOC727431 | gustatory receptor for sugar taste 64f, transcript variant X3 | chemosensation | - | - | - | 2.36 |
| Or12 | odorant receptor 12 | Odorant receptor | - | - | - | 2.34 |
| SP34 | serine protease 34 | Toll/TLR | - | - | - | 3.83 |
| TyHyd | tyrosine hydroxylase | Tyrosine metabolism | - | - | - | 2.04 |
| LOC107964335 | melatonin receptor type 1B-B-like | Signaling receptor activity | - | - | - | 5.14 |
| LOC410624 | serine proteinase stubble, transcript variant X4 | Toll/TLR | - | - | - | 2.81 |
| Gene Name | Gene Description | Category | (log2 Fold Change) | |||
|---|---|---|---|---|---|---|
| VF | VF+FPF | VI | VI+FPF | |||
| Obp13 | odorant binding protein 13 | odorant-binding protein (OBP) | - | −2.35 | −2.85 | −2.33 |
| Apamin | apamin protein | - | - | - | −5.51 | −5.64 |
| Apd-3 | apidermin 3 | innate immune | - | - | −2.95 | −2.25 |
| Est-6 | venom carboxylesterase-6 | - | - | - | −2.30 | - |
| Eth | ecdysis triggering hormone | - | - | - | −2.71 | −2.26 |
| LOC102653899 | probable cytochrome P450 6a13 | - | - | - | −3.59 | −3.05 |
| LOC102654530 | odorant receptor 67a-like, transcript variant X2 | Odorant receptor (OR) | - | - | −2.43 | - |
| LOC724308 | serine protease 53 | Toll/TLR | - | - | −2.14 | −2.58 |
| LOC724436 | phospholipase A2-like | Eicosanoid | - | - | −4.63 | −3.76 |
| Pla2 | phospholipase A2 | Eicosanoid | - | - | −5.82 | −6.47 |
| Vg | vitellogenin | vitellogenin | - | - | −3.29 | −2.99 |
| LOC406145 | secapin | innate immune response | - | - | −6.10 | −5.33 |
| LOC678674 | venom allergen Api m 6 | - | - | - | −5.27 | −7.50 |
| LOC406114 | alpha-amylase | - | - | - | −3.44 | −4.00 |
| Melt | melittin | innate immune | - | - | −5.46 | −4.43 |
| LOC100576797 | acyl-CoA Delta (11) desaturase, transcript variant X2 | lipid metabolism | - | - | −5.21 | −4.89 |
| Obp17 | odorant binding protein 17 | OBP | - | - | −2.56 | −3.02 |
| Obp3 | odorant binding protein 3 | OBP | - | - | −2.41 | −3.50 |
| LOC724175 | probable cytochrome P450 304a1 | - | - | - | −2.81 | −3.13 |
| Mcdp | mast cell-degranulating peptide | - | - | - | −6.30 | −6.13 |
| LOC408603 | glucose dehydrogenase [FAD, quinone] | - | - | - | −2.96 | −4.68 |
| LOC727037 | phospholipase A1 member A, transcript variant X3 | Eicosanoid | - | - | −2.34 | −2.73 |
| Obp14 | odorant binding protein 14 | OBP | - | - | −3.71 | −2.82 |
| COX3 | Cytochrome c oxidase subunit 3 | mitochondrial energy metabolism | - | - | - | −2.16 |
| CSP6 | chemosensory protein 6 | - | - | - | −2.22 | |
| Obp21 | odorant binding protein 21 | OBP | - | - | - | −2.69 |
| LOC725922 | mitochondrial basic amino acids transporter | Transport | - | - | - | −2.88 |
| LOC724211 | cytochrome P450 9e2 | - | - | - | - | −2.31 |
| LOC551197 | probable cytochrome P450 6a13 | - | - | - | - | −2.03 |
| LOC410894 | chymotrypsin-1 | proteolysis | - | - | - | −2.21 |
| LOC411307 | mitochondrial enolase superfamily member 1 | cellular amino acid catabolic process | - | - | - | −2.02 |
| GO: Category | GO: Biological Process Term | p Value 1 | DEGs 2 | Category 3 | Treatment |
| GO:0019320 | Hexose catabolic process | 1.37 × 10−3 | 1 | 3 | VI, VF, VI+FPF, VF+FPF |
| GO:0019388 | Galactose catabolic process | 1.37 × 10−3 | 1 | 3 | |
| GO:0033499 | Galactose catabolic process via UDP-galactose | 1.37 × 10−3 | 1 | 3 | |
| GO:0046365 | Monosaccharide catabolic process | 1.37 × 10−3 | 1 | 4 | |
| GO:0006012 | Galactose metabolic process | 2.74 × 10−3 | 1 | 10 | |
| GO:0005996 | Monosaccharide metabolic process | 6.17 × 10−3 | 1 | 36 | |
| GO:0016052 | Carbohydrate catabolic process | 6.17 × 10−3 | 1 | 27 | |
| GO:0019318 | Hexose metabolic process | 6.17 × 10−3 | 1 | 34 | |
| GO:0044282 | Small molecule catabolic process | 7.62 × 10−3 | 1 | 50 | |
| GO:0005975 | Carbohydrate metabolic process | 2.15 × 10−2 | 1 | 157 | |
| GO:0009617 | Response to bacterium | 3.51 × 10−6 | 2 | 9 | VI, VF+FPF, VI+FPF |
| GO:0042742 | Defense response to bacterium | 3.51 × 10−6 | 2 | 9 | |
| GO:0006952 | Defense response | 5.39 × 10−6 | 2 | 22 | |
| GO:0006955 | Immune response | 5.39 × 10−6 | 2 | 24 | |
| GO:0009607 | Response to biotic stimulus | 5.39 × 10−6 | 2 | 21 | |
| GO:0043207 | Response to external biotic stimulus | 5.39 × 10−6 | 2 | 21 | |
| GO:0044419 | Biological process involved in interspecies interaction between organisms | 5.39 × 10−6 | 2 | 24 | |
| GO:0045087 | Innate immune response | 5.39 × 10−6 | 2 | 17 | |
| GO:0051707 | Response to other organism | 5.39 × 10−6 | 2 | 21 | |
| GO:0098542 | Defense response to other organism | 5.39 × 10−6 | 2 | 21 | |
| GO:0050896 | Response to stimulus | 5.67 × 10−3 | 2 | 902 | |
| GO:0050832 | Defense response to fungus | 1.81 × 10−3 | 1 | 1 | VF+FPF |
| GO:0050830 | Defense response to Gram-positive bacterium | 1.81 × 10−3 | 1 | 1 | |
| GO:0050829 | Defense response to Gram-negative bacterium | 1.81 × 10−3 | 1 | 1 | |
| GO:0048651 | Polyphenic determination, influence by environmental factors | 1.81 × 10−3 | 1 | 1 | |
| GO:0048650 | Caste determination, influence by environmental factors | 1.81 × 10−3 | 1 | 1 | |
| GO:0009620 | Response to fungus | 1.81 × 10−3 | 1 | 1 | |
| GO:0048648 | Caste determination | 1.81 × 10−3 | 1 | 1 | |
| GO:0048647 | Polyphenic determination | 1.81 × 10−3 | 1 | 1 | |
| GO:0001906 | Cell killing | 2.90 × 10−3 | 1 | 2 | |
| GO:0031640 | Killing of cells of other organisms | 2.90 × 10−3 | 1 | 2 |
| GO: Category | GO: Biological Process Term | p Value 1 | DEG 2 | Category 3 | Treatment |
| GO:0035821 | Modulation of the processes of other organisms | 2.96 × 10−5 | 3 | 4 | VI, VI+FPF |
| GO:0055114 | Obsolete oxidation–reduction process | 1.55 × 10−3 | 8 | 334 | |
| GO:0010876 | Lipid localization | 1.93 × 10−3 | 4 | 56 | |
| GO:0006869 | Lipid transport | 1.93 × 10−3 | 4 | 54 | |
| GO:0044419 | Biological process involved in interspecies interaction between organisms | 2.86 × 10−3 | 3 | 24 | |
| GO:0015909 | Long-chain fatty acid transport | 4.23 × 10−3 | 2 | 7 | |
| GO:0071715 | Icosanoid transport | 4.23 × 10−3 | 2 | 7 | |
| GO:0050482 | Arachidonic acid secretion | 4.23 × 10−3 | 2 | 7 | |
| GO:0044255 | Cellular lipid metabolic process | 4.23 × 10−3 | 5 | 167 | |
| GO:0032309 | Icosanoid secretion | 4.23 × 10−3 | 2 | 7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al Naggar, Y.; Shafiey, H.; Paxton, R.J. Transcriptomic Responses Underlying the High Virulence of Black Queen Cell Virus and Sacbrood Virus following a Change in Their Mode of Transmission in Honey Bees (Apis mellifera). Viruses 2023, 15, 1284. https://doi.org/10.3390/v15061284
Al Naggar Y, Shafiey H, Paxton RJ. Transcriptomic Responses Underlying the High Virulence of Black Queen Cell Virus and Sacbrood Virus following a Change in Their Mode of Transmission in Honey Bees (Apis mellifera). Viruses. 2023; 15(6):1284. https://doi.org/10.3390/v15061284
Chicago/Turabian StyleAl Naggar, Yahya, Hassan Shafiey, and Robert J. Paxton. 2023. "Transcriptomic Responses Underlying the High Virulence of Black Queen Cell Virus and Sacbrood Virus following a Change in Their Mode of Transmission in Honey Bees (Apis mellifera)" Viruses 15, no. 6: 1284. https://doi.org/10.3390/v15061284
APA StyleAl Naggar, Y., Shafiey, H., & Paxton, R. J. (2023). Transcriptomic Responses Underlying the High Virulence of Black Queen Cell Virus and Sacbrood Virus following a Change in Their Mode of Transmission in Honey Bees (Apis mellifera). Viruses, 15(6), 1284. https://doi.org/10.3390/v15061284