
Piperidine CD4-Mimetic Compounds Expose Vulnerable Env Epitopes Sensitizing HIV-1-Infected Cells to ADCC

 , , ,
, , ,
Abstract
1. Introduction
2. Results
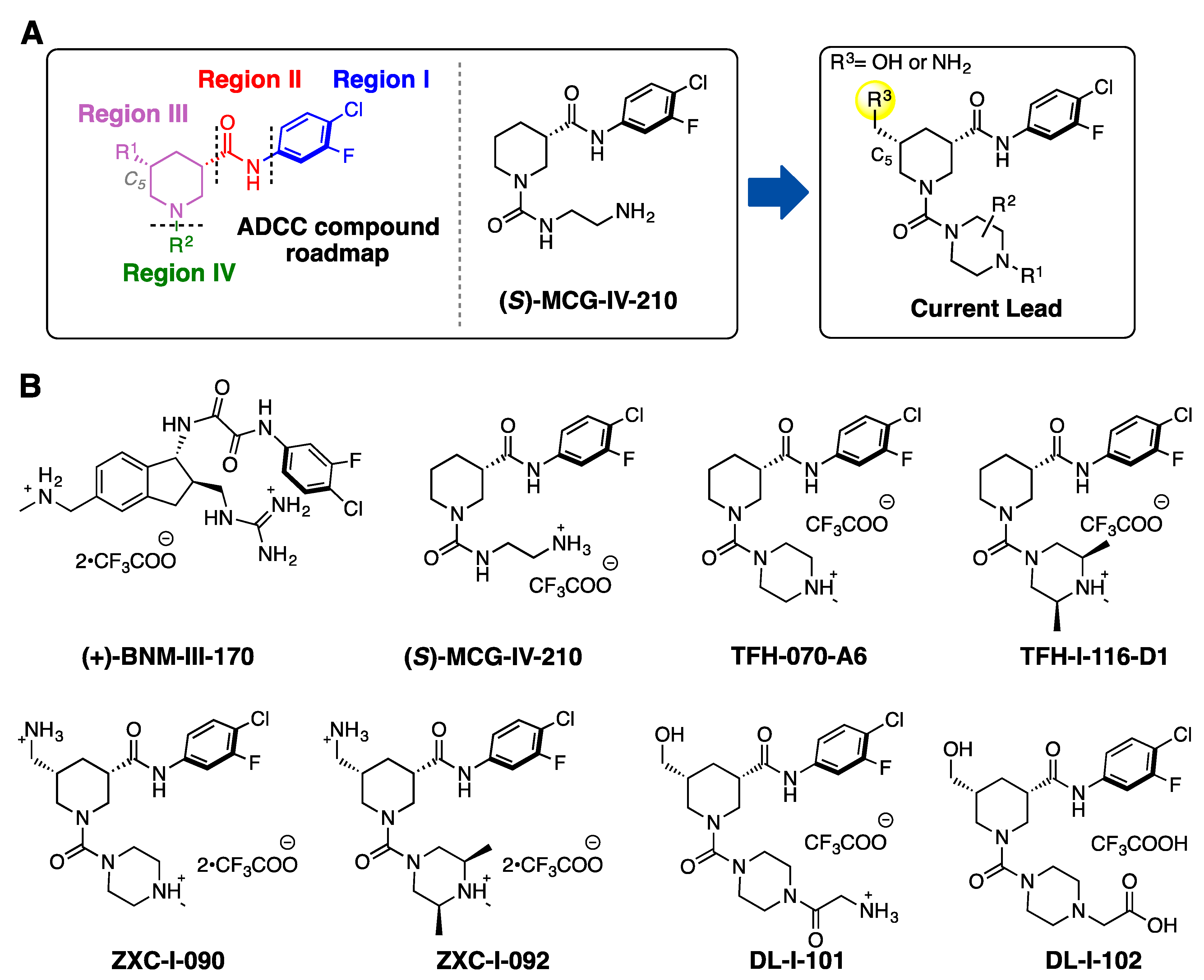
2.1. New Piperidine Analogs
2.2. The Design and Synthesis of New CD4mc Piperidine Analogs
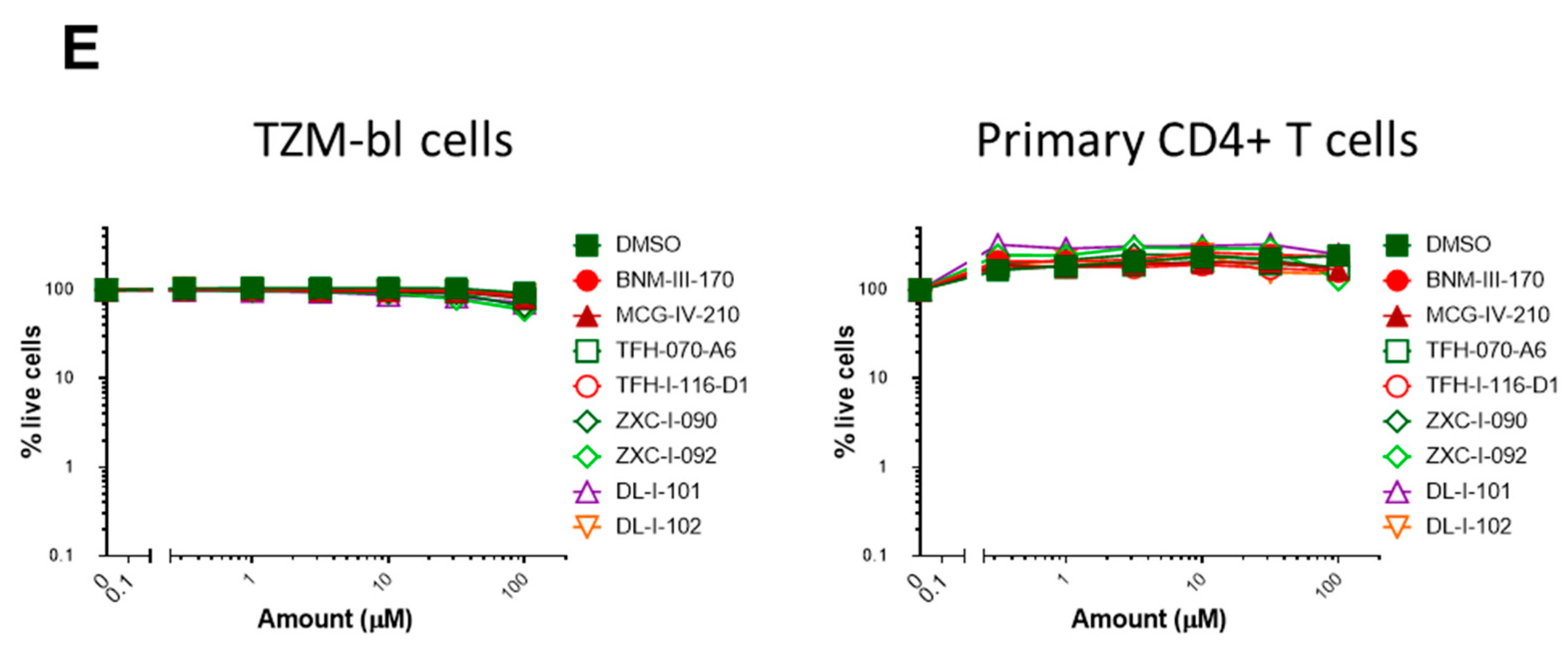
2.3. Biological Testing
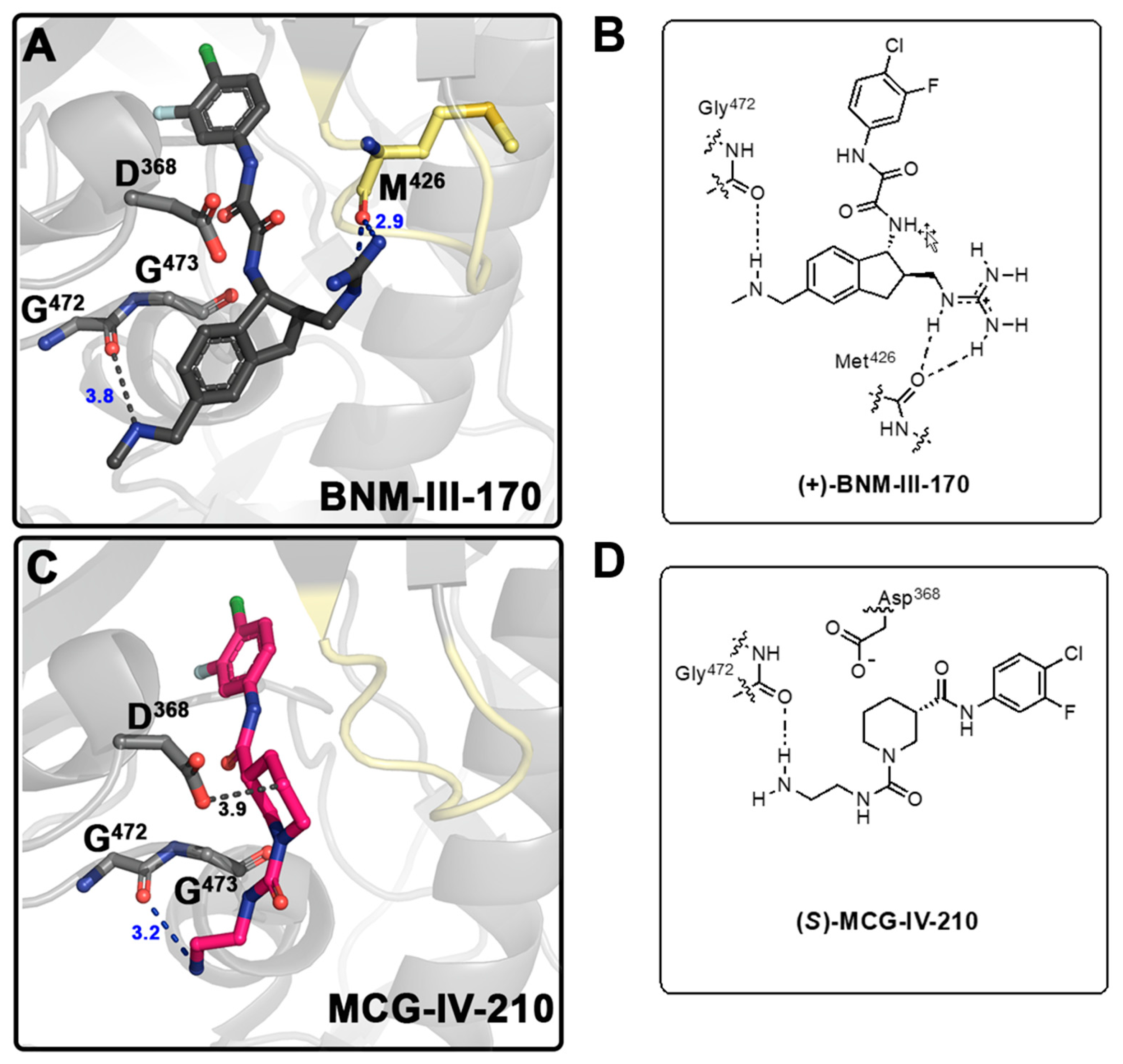
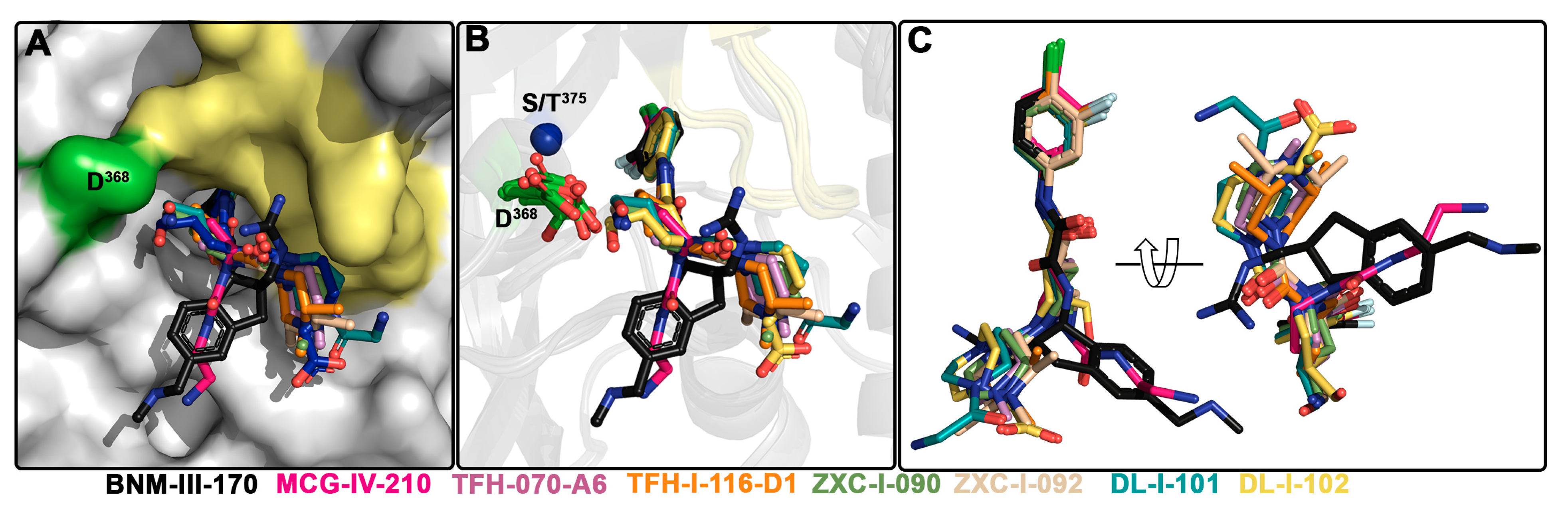
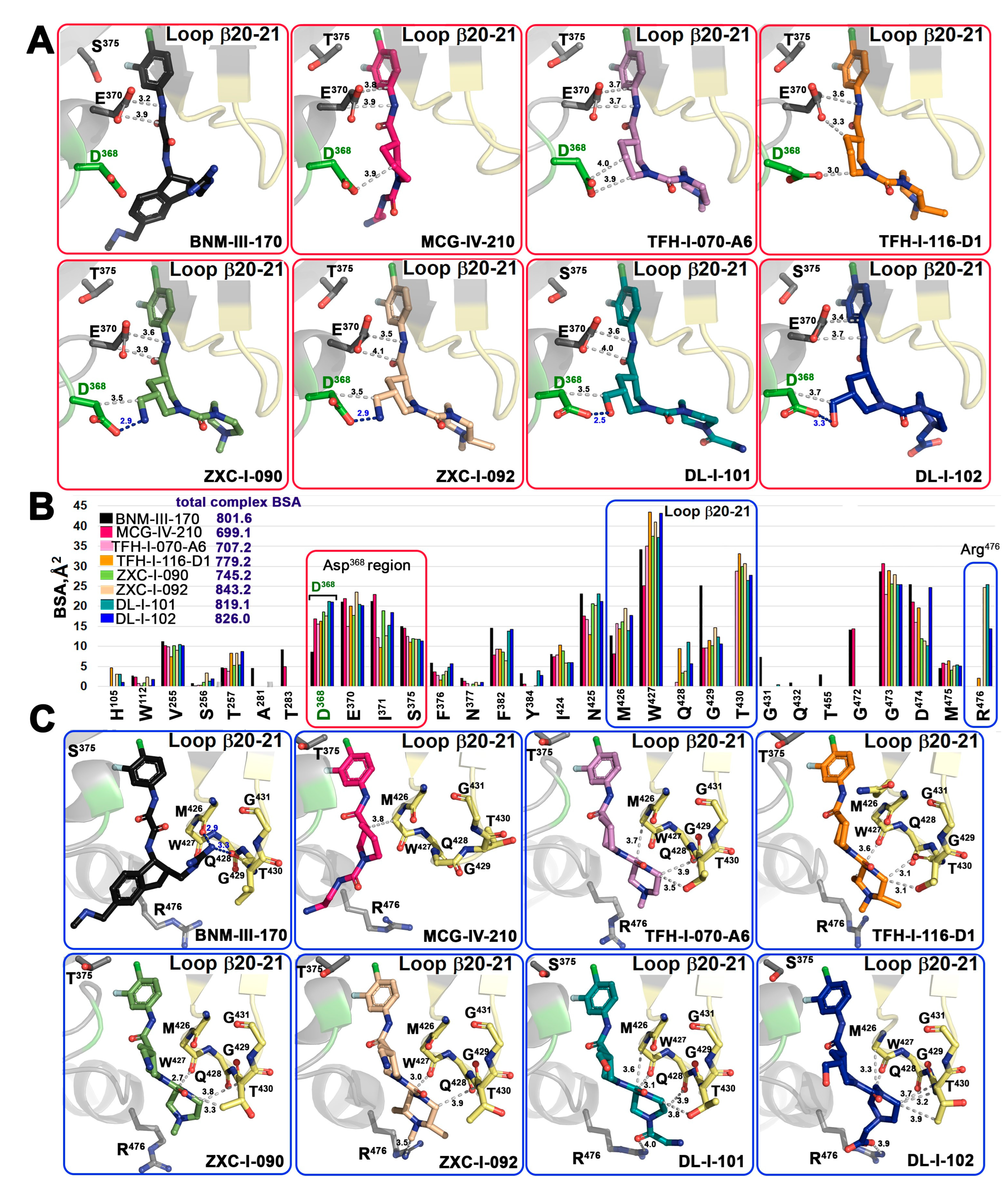
2.4. Structural Basis of Interaction of (S)-MCG-IV-210 Derivatives with a gp120 Core
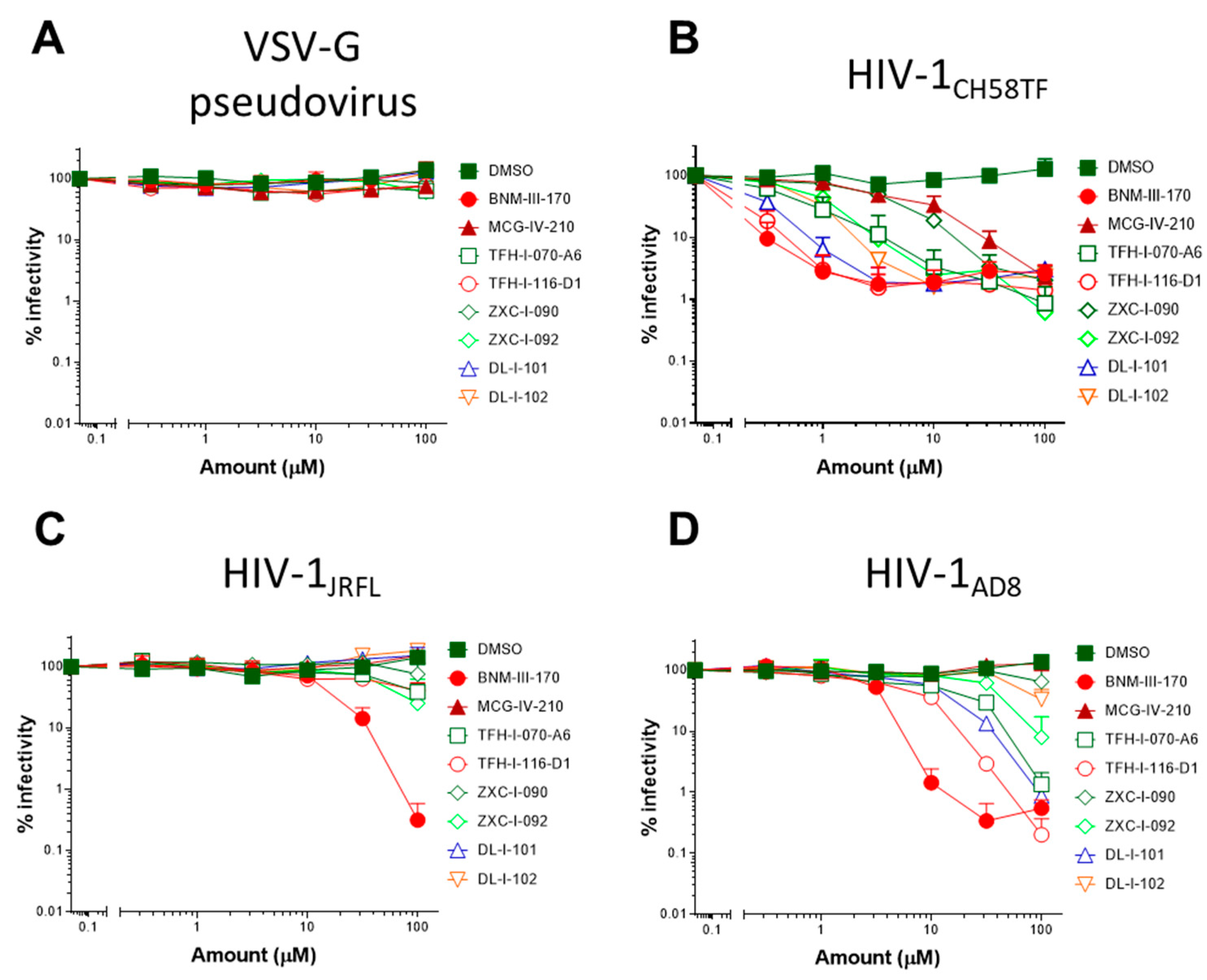
2.5. (S)-MCG-IV-210 Derivatives Inhibit Viral Infection
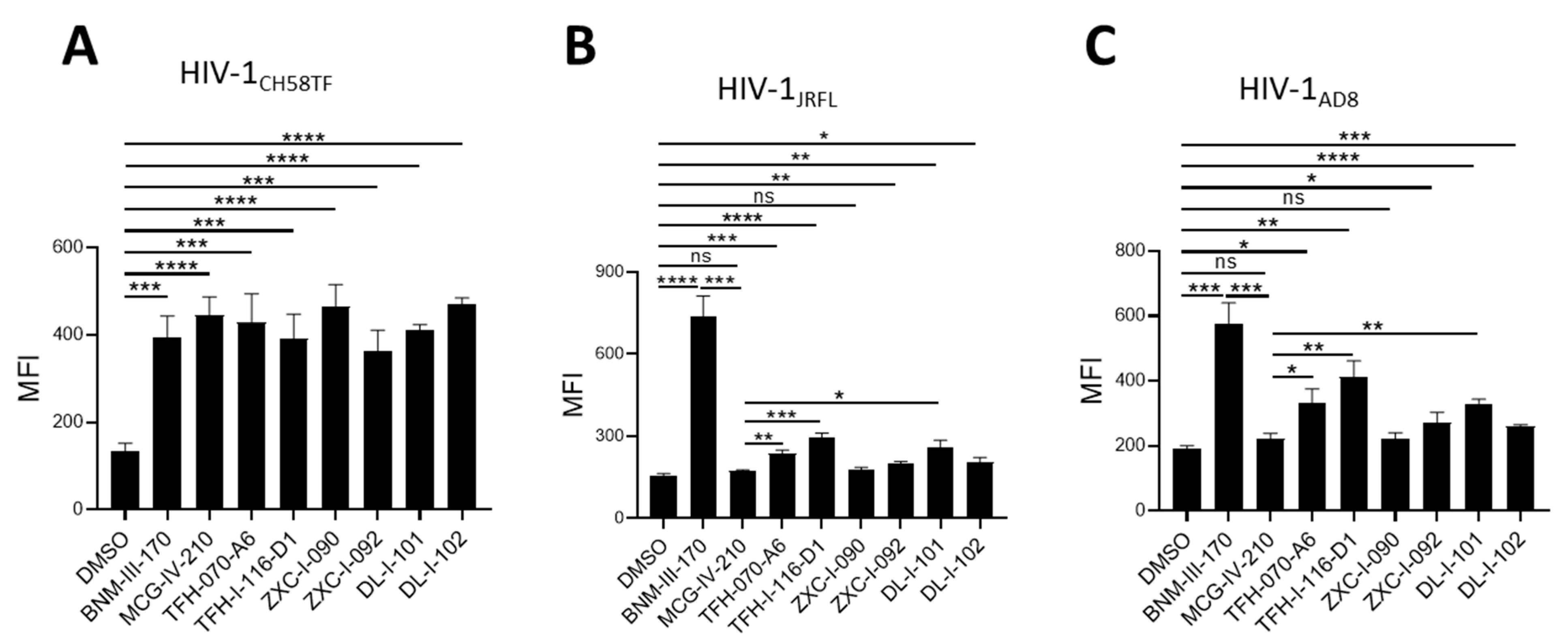
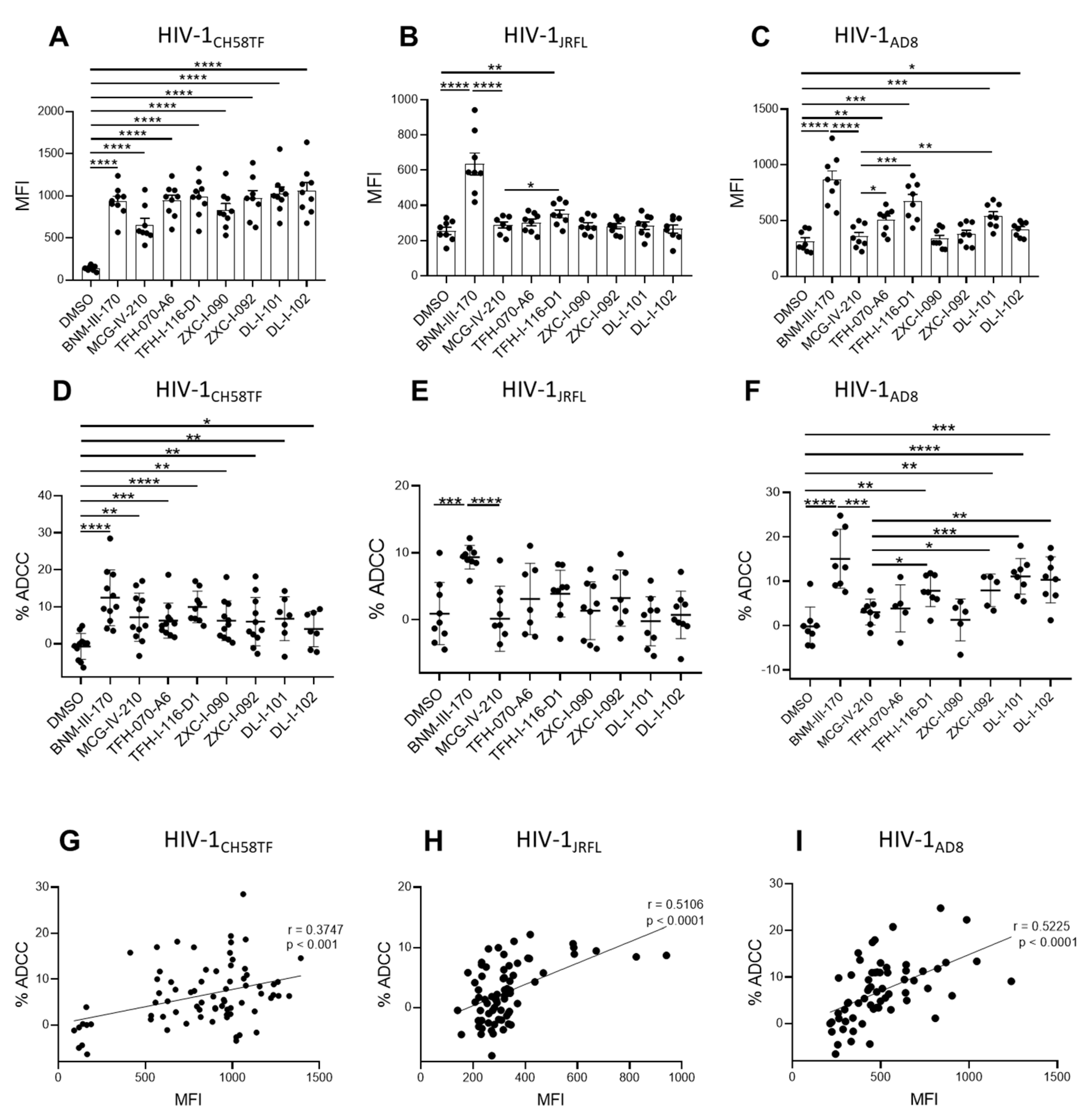
2.6. Sensitization of HIV-1-Infected Cells to ADCC
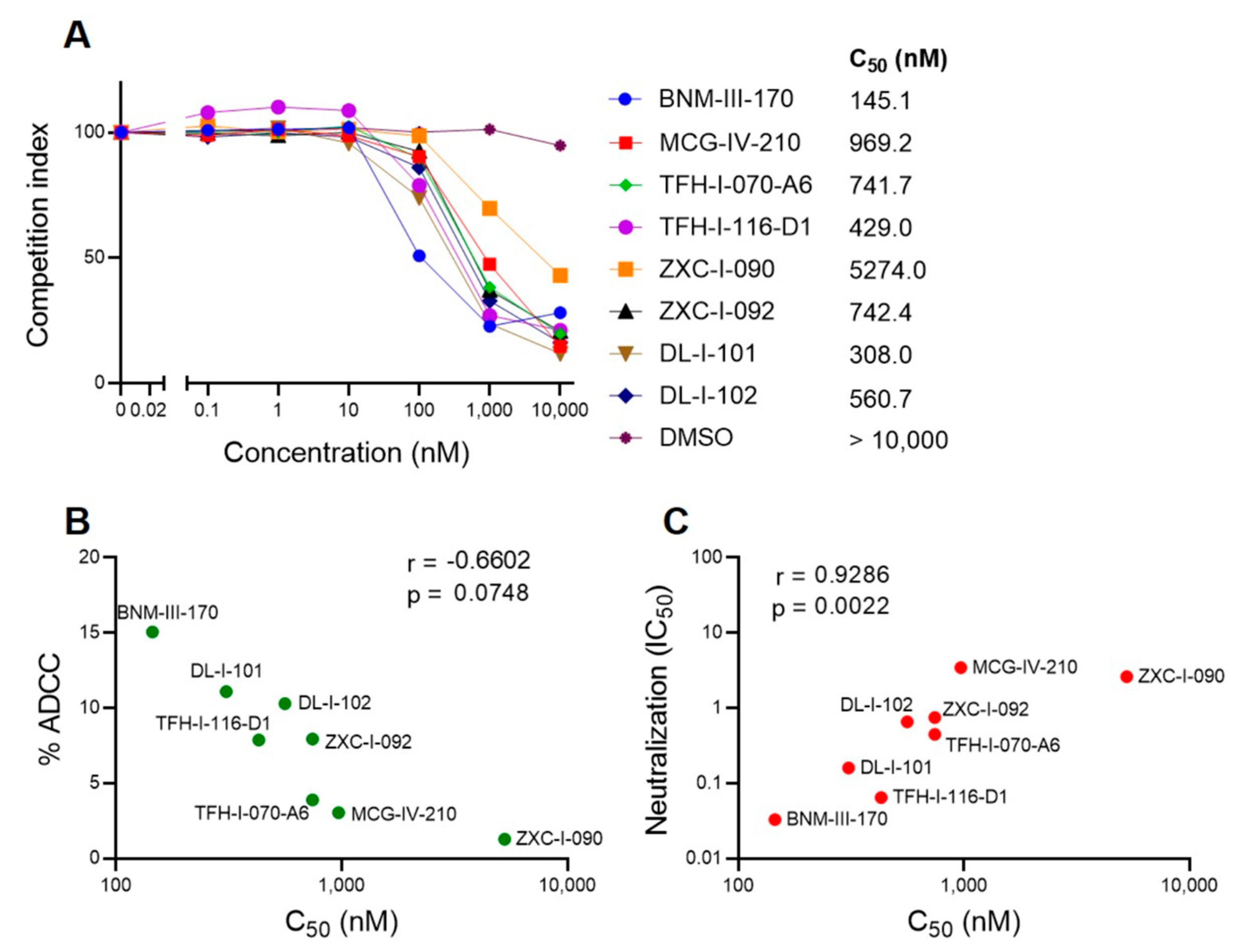
2.7. BLI Competition Assay
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Cell Lines and Isolation of Primary Cells
4.3. Chemical Synthesis: General Considerations
4.4. Viral Production and Infection of Primary CD4+ T Cells
4.5. Viral Neutralization
4.6. Antibodies and Plasma
4.7. Flow Cytometry Analysis of Cell-Surface Staining
4.8. Cell Viability Test
4.9. ADCC FACS-Based Assay
4.10. Statistical Analysis
4.11. CRF01_AE Core e Expression and Purification
4.12. Biolayer Interferometry (BLI) Competition Assay
4.13. Crystallization of gp120 Cores Complex with CD4mc
4.14. Data Collection, Structure Solution and Refinement
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Disclaimer
References
- Allan, J.S.; Coligan, J.E.; Barin, F.; McLane, M.F.; Sodroski, J.G.; Rosen, C.A.; Haseltine, W.A.; Lee, T.H.; Essex, M. Major glycoprotein antigens that induce antibodies in AIDS patients are encoded by HTLV-III. Science 1985, 228, 1091–1094. [Google Scholar] [CrossRef] [PubMed]
- Robey, W.G.; Safai, B.; Oroszlan, S.; Arthur, L.O.; Gonda, M.A.; Gallo, R.C.; Fischinger, P.J. Characterization of envelope and core structural gene products of HTLV-III with sera from AIDS patients. Science 1985, 228, 593–595. [Google Scholar] [CrossRef] [PubMed]
- Wyatt, R.; Sodroski, J. The HIV-1 envelope glycoproteins: Fusogens, antigens, and immunogens. Science 1998, 280, 1884–1888. [Google Scholar] [CrossRef]
- Furuta, R.A.; Wild, C.T.; Weng, Y.; Weiss, C.D. Capture of an early fusion-active conformation of HIV-1 gp41. Nat. Struct. Biol. 1998, 5, 276–279. [Google Scholar] [CrossRef] [PubMed]
- Decker, J.M.; Bibollet-Ruche, F.; Wei, X.; Wang, S.; Levy, D.N.; Wang, W.; Delaporte, E.; Peeters, M.; Derdeyn, C.A.; Allen, S.; et al. Antigenic conservation and immunogenicity of the HIV coreceptor binding site. J. Exp. Med. 2005, 201, 1407–1419. [Google Scholar] [CrossRef]
- Veillette, M.; Coutu, M.; Richard, J.; Batraville, L.A.; Dagher, O.; Bernard, N.; Tremblay, C.; Kaufmann, D.E.; Roger, M.; Finzi, A. The HIV-1 gp120 CD4-bound conformation is preferentially targeted by antibody-dependent cellular cytotoxicity-mediating antibodies in sera from HIV-1-infected individuals. J. Virol. 2015, 89, 545–551. [Google Scholar] [CrossRef]
- Veillette, M.; Desormeaux, A.; Medjahed, H.; Gharsallah, N.E.; Coutu, M.; Baalwa, J.; Guan, Y.; Lewis, G.; Ferrari, G.; Hahn, B.H.; et al. Interaction with cellular CD4 exposes HIV-1 envelope epitopes targeted by antibody-dependent cell-mediated cytotoxicity. J. Virol. 2014, 88, 2633–2644. [Google Scholar] [CrossRef]
- Madani, N.; Schon, A.; Princiotto, A.M.; Lalonde, J.M.; Courter, J.R.; Soeta, T.; Ng, D.; Wang, L.; Brower, E.T.; Xiang, S.H.; et al. Small-molecule CD4 mimics interact with a highly conserved pocket on HIV-1 gp120. Structure 2008, 16, 1689–1701. [Google Scholar] [CrossRef]
- Madani, N.; Princiotto, A.M.; Schon, A.; LaLonde, J.; Feng, Y.; Freire, E.; Park, J.; Courter, J.R.; Jones, D.M.; Robinson, J.; et al. CD4-mimetic small molecules sensitize human immunodeficiency virus to vaccine-elicited antibodies. J. Virol. 2014, 88, 6542–6555. [Google Scholar] [CrossRef]
- Prevost, J.; Tolbert, W.D.; Medjahed, H.; Sherburn, R.T.; Madani, N.; Zoubchenok, D.; Gendron-Lepage, G.; Gaffney, A.E.; Grenier, M.C.; Kirk, S.; et al. The HIV-1 Env gp120 Inner Domain Shapes the Phe43 Cavity and the CD4 Binding Site. mBio 2020, 11, e00280-20. [Google Scholar] [CrossRef]
- Kwong, P.D.; Wyatt, R.; Robinson, J.; Sweet, R.W.; Sodroski, J.; Hendrickson, W.A. Structure of an HIV gp120 envelope glycoprotein in complex with the CD4 receptor and a neutralizing human antibody. Nature 1998, 393, 648–659. [Google Scholar] [CrossRef]
- Zhao, Q.; Ma, L.; Jiang, S.; Lu, H.; Liu, S.; He, Y.; Strick, N.; Neamati, N.; Debnath, A.K. Identification of N-phenyl-N′-(2,2,6,6-tetramethyl-piperidin-4-yl)-oxalamides as a new class of HIV-1 entry inhibitors that prevent gp120 binding to CD4. Virology 2005, 339, 213–225. [Google Scholar] [CrossRef]
- Madani, N.; Princiotto, A.M.; Mach, L.; Ding, S.; Prevost, J.; Richard, J.; Hora, B.; Sutherland, L.; Zhao, C.A.; Conn, B.P.; et al. A CD4-mimetic compound enhances vaccine efficacy against stringent immunodeficiency virus challenge. Nat. Commun. 2018, 9, 2363. [Google Scholar] [CrossRef] [PubMed]
- Madani, N.; Princiotto, A.M.; Zhao, C.; Jahanbakhshsefidi, F.; Mertens, M.; Herschhorn, A.; Melillo, B.; Smith, A.B., III; Sodroski, J. Activation and Inactivation of Primary Human Immunodeficiency Virus Envelope Glycoprotein Trimers by CD4-Mimetic Compounds. J. Virol. 2017, 91, e01880-16. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.; Verly, M.M.; Princiotto, A.; Melillo, B.; Moody, T.; Bradley, T.; Easterhoff, D.; Roger, M.; Hahn, B.H.; Madani, N.; et al. Small Molecule CD4-Mimetics Sensitize HIV-1-infected Cells to ADCC by Antibodies Elicited by Multiple Envelope Glycoprotein Immunogens in Non-Human Primates. AIDS Res. Hum. Retrovir. 2016, 33, 428–431. [Google Scholar] [CrossRef]
- Richard, J.; Prevost, J.; von Bredow, B.; Ding, S.; Brassard, N.; Medjahed, H.; Coutu, M.; Melillo, B.; Bibollet-Ruche, F.; Hahn, B.H.; et al. BST-2 Expression Modulates Small CD4-Mimetic Sensitization of HIV-1-Infected Cells to Antibody-Dependent Cellular Cytotoxicity. J. Virol. 2017, 91, e00219-17. [Google Scholar] [CrossRef] [PubMed]
- Richard, J.; Veillette, M.; Brassard, N.; Iyer, S.S.; Roger, M.; Martin, L.; Pazgier, M.; Schon, A.; Freire, E.; Routy, J.P.; et al. CD4 mimetics sensitize HIV-1-infected cells to ADCC. Proc. Natl. Acad. Sci. USA 2015, 112, E2687–E2694. [Google Scholar] [CrossRef]
- Ding, S.; Grenier, M.C.; Tolbert, W.D.; Vezina, D.; Sherburn, R.; Richard, J.; Prevost, J.; Chapleau, J.P.; Gendron-Lepage, G.; Medjahed, H.; et al. A New Family of Small-Molecule CD4-Mimetic Compounds Contacts Highly Conserved Aspartic Acid 368 of HIV-1 gp120 and Mediates Antibody-Dependent Cellular Cytotoxicity. J. Virol. 2019, 93, e01325-19. [Google Scholar] [CrossRef]
- Kwon, Y.D.; Finzi, A.; Wu, X.; Dogo-Isonagie, C.; Lee, L.K.; Moore, L.R.; Schmidt, S.D.; Stuckey, J.; Yang, Y.; Zhou, T.; et al. Unliganded HIV-1 gp120 core structures assume the CD4-bound conformation with regulation by quaternary interactions and variable loops. Proc. Natl. Acad. Sci. USA 2012, 109, 5663–5668. [Google Scholar] [CrossRef]
- Zoubchenok, D.; Veillette, M.; Prevost, J.; Sanders-Buell, E.; Wagh, K.; Korber, B.; Chenine, A.L.; Finzi, A. Histidine 375 Modulates CD4 Binding in HIV-1 CRF01_AE Envelope Glycoproteins. J. Virol. 2017, 91, e02151-16. [Google Scholar] [CrossRef]
- Grenier, M.C.; Ding, S.; Vezina, D.; Chapleau, J.P.; Tolbert, W.D.; Sherburn, R.; Schon, A.; Somisetti, S.; Abrams, C.F.; Pazgier, M.; et al. Optimization of Small Molecules That Sensitize HIV-1 Infected Cells to Antibody-Dependent Cellular Cytotoxicity. ACS Med. Chem. Lett. 2020, 11, 371–378. [Google Scholar] [CrossRef]
- Mascola, J.R.; D’Souza, P.; Gilbert, P.; Hahn, B.H.; Haigwood, N.L.; Morris, L.; Petropoulos, C.J.; Polonis, V.R.; Sarzotti, M.; Montefiori, D.C. Recommendations for the design and use of standard virus panels to assess neutralizing antibody responses elicited by candidate human immunodeficiency virus type 1 vaccines. J. Virol. 2005, 79, 10103–10107. [Google Scholar] [CrossRef]
- Seaman, M.S.; Janes, H.; Hawkins, N.; Grandpre, L.E.; Devoy, C.; Giri, A.; Coffey, R.T.; Harris, L.; Wood, B.; Daniels, M.G.; et al. Tiered categorization of a diverse panel of HIV-1 Env pseudoviruses for assessment of neutralizing antibodies. J. Virol. 2010, 84, 1439–1452. [Google Scholar] [CrossRef] [PubMed]
- Hraber, P.; Korber, B.; Wagh, K.; Montefiori, D.; Roederer, M. A single, continuous metric to define tiered serum neutralization potency against HIV. eLife 2018, 7, e31805. [Google Scholar] [CrossRef] [PubMed]
- Montefiori, D.C.; Roederer, M.; Morris, L.; Seaman, M.S. Neutralization tiers of HIV-1. Curr. Opin. HIV AIDS 2018, 13, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Weiss, M.S. Global indicators of X-ray data quality. J. Appl. Cryst. 2001, 34, 130–135. [Google Scholar] [CrossRef]
- Karplus, P.A.; Diederichs, K. Linking crystallographic model and data quality. Science 2012, 336, 1030–1033. [Google Scholar] [CrossRef]
- Brunger, A.T. Free R value: Cross-validation in crystallography. In Methods in Enzymology; Academic Press: New York, NY, USA, 1997; Volume 277, pp. 366–396. [Google Scholar]
- Melillo, B.; Liang, S.; Park, J.; Schon, A.; Courter, J.R.; LaLonde, J.M.; Wendler, D.J.; Princiotto, A.M.; Seaman, M.S.; Freire, E.; et al. Small-Molecule CD4-Mimics: Structure-Based Optimization of HIV-1 Entry Inhibition. ACS Med. Chem. Lett. 2016, 7, 330–334. [Google Scholar] [CrossRef]
- Richard, J.; Pacheco, B.; Gohain, N.; Veillette, M.; Ding, S.; Alsahafi, N.; Tolbert, W.D.; Prevost, J.; Chapleau, J.P.; Coutu, M.; et al. Co-receptor Binding Site Antibodies Enable CD4-Mimetics to Expose Conserved Anti-cluster A ADCC Epitopes on HIV-1 Envelope Glycoproteins. EBioMedicine 2016, 12, 208–218. [Google Scholar] [CrossRef]
- Alsahafi, N.; Bakouche, N.; Kazemi, M.; Richard, J.; Ding, S.; Bhattacharyya, S.; Das, D.; Anand, S.P.; Prevost, J.; Tolbert, W.D.; et al. An Asymmetric Opening of HIV-1 Envelope Mediates Antibody-Dependent Cellular Cytotoxicity. Cell Host Microbe 2019, 25, 578–587.e5. [Google Scholar] [CrossRef]
- Laumaea, A.; Marchitto, L.; Ding, S.; Beaudoin-Bussieres, G.; Prevost, J.; Gasser, R.; Chatterjee, D.; Gendron-Lepage, G.; Medjahed, H.; Chen, H.C.; et al. Small CD4 mimetics sensitize HIV-1-infected macrophages to antibody-dependent cellular cytotoxicity. Cell Rep. 2023, 42, 111983. [Google Scholar] [CrossRef] [PubMed]
- Prevost, J.; Anand, S.P.; Rajashekar, J.K.; Zhu, L.; Richard, J.; Goyette, G.; Medjahed, H.; Gendron-Lepage, G.; Chen, H.C.; Chen, Y.; et al. HIV-1 Vpu restricts Fc-mediated effector functions in vivo. Cell Rep. 2022, 41, 111624. [Google Scholar] [CrossRef] [PubMed]
- Rajashekar, J.K.; Richard, J.; Beloor, J.; Prevost, J.; Anand, S.P.; Beaudoin-Bussieres, G.; Shan, L.; Herndler-Brandstetter, D.; Gendron-Lepage, G.; Medjahed, H.; et al. Modulating HIV-1 envelope glycoprotein conformation to decrease the HIV-1 reservoir. Cell Host Microbe 2021, 29, 904–916 e6. [Google Scholar] [CrossRef] [PubMed]
- Richard, J.; Prevost, J.; Baxter, A.E.; von Bredow, B.; Ding, S.; Medjahed, H.; Delgado, G.G.; Brassard, N.; Sturzel, C.M.; Kirchhoff, F.; et al. Uninfected Bystander Cells Impact the Measurement of HIV-Specific Antibody-Dependent Cellular Cytotoxicity Responses. mBio 2018, 9, e00358-18. [Google Scholar] [CrossRef]
- Ding, S.; Veillette, M.; Coutu, M.; Prevost, J.; Scharf, L.; Bjorkman, P.J.; Ferrari, G.; Robinson, J.E.; Sturzel, C.; Hahn, B.H.; et al. A Highly Conserved Residue of the HIV-1 gp120 Inner Domain Is Important for Antibody-Dependent Cellular Cytotoxicity Responses Mediated by Anti-cluster A Antibodies. J. Virol. 2016, 90, 2127–2134. [Google Scholar] [CrossRef] [PubMed]
- Madani, N.; Princiotto, A.M.; Easterhoff, D.; Bradley, T.; Luo, K.; Williams, W.B.; Liao, H.X.; Moody, M.A.; Phad, G.E.; Vazquez Bernat, N.; et al. Antibodies Elicited by Multiple Envelope Glycoprotein Immunogens in Primates Neutralize Primary Human Immunodeficiency Viruses (HIV-1) Sensitized by CD4-Mimetic Compounds. J. Virol. 2016, 90, 5031–5046. [Google Scholar] [CrossRef]
- Finzi, A.; Xiang, S.H.; Pacheco, B.; Wang, L.; Haight, J.; Kassa, A.; Danek, B.; Pancera, M.; Kwong, P.D.; Sodroski, J. Topological layers in the HIV-1 gp120 inner domain regulate gp41 interaction and CD4-triggered conformational transitions. Mol. Cell 2010, 37, 656–667. [Google Scholar] [CrossRef]
- Richard, J.; Veillette, M.; Batraville, L.A.; Coutu, M.; Chapleau, J.P.; Bonsignori, M.; Bernard, N.; Tremblay, C.; Roger, M.; Kaufmann, D.E.; et al. Flow cytometry-based assay to study HIV-1 gp120 specific antibody-dependent cellular cytotoxicity responses. J. Virol. Methods 2014, 208, 107–114. [Google Scholar] [CrossRef]
- Otwinowski, Z.; Minor, W. Processing of X-ray diffraction data collected in oscillation mode. In Meth Enzymol; Carter, C.W.J., Sweet, R.M., Eds.; Academic Press: New York, NY, USA, 1997; Volume 276, pp. 307–326. [Google Scholar]
- Collaborative Computational Project. The CCP4 suite: Programs for protein crystallography. Acta Crystallogr. Sect. D Biol. Crystallogr. 1994, 50 Pt 5, 760–763. [Google Scholar] [CrossRef]
- Adams, P.D.; Afonine, P.V.; Bunkoczi, G.; Chen, V.B.; Davis, I.W.; Echols, N.; Headd, J.J.; Hung, L.W.; Kapral, G.J.; Grosse-Kunstleve, R.W.; et al. PHENIX: A comprehensive Python-based system for macromolecular structure solution. Acta Cryst. D Biol Cryst. 2010, D66, 213–221. [Google Scholar] [CrossRef]
- Laskowski, R.A.; Swindells, M.B. LigPlot+: Multiple ligand-protein interaction diagrams for drug discovery. J. Chem. Inf. Model. 2011, 51, 2778–2786. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LM/HT gp120CRF01_AE Coree—TFH-I-070-A6 | LM/HT gp120CRF01_AE Coree—TFH-I-116-D1 | LM/HT gp120CRF01_AE Coree—ZXC-I-090 | LM/HT gp120CRF01_AE Coree—ZXC-I-092 | LM/HS gp120CRF01_AE Coree—DL-I-101 | LM/HS gp120CRF01_AE Coree—DL-I-102 | |
|---|---|---|---|---|---|---|
| Data collection | ||||||
| Wavelength, Ǻ | 0.979 | 0.979 | 0.979 | 0.979 | 0.979 | 0.979 |
| Space group | P212121 | P212121 | P212121 | P212121 | P212121 | P212121 |
| Cell parameters | ||||||
| a, b, c, Å | 63.6, 66.3, 91.4 | 64.7, 66.0, 87.8 | 61.8, 65.9, 91.2 | 60.0, 65.2, 87.3 | 67.7, 66.9, 92.2 | 194.5, 86.6, 57.7 |
| α, β, γ, ° | 90, 90, 90 | 90, 90, 90 | 90, 90, 90 | 90, 909, 90 | 90, 90, 90 | 90, 90, 90 |
| Molecules/a.u. | 1 | 1 | 1 | 1 | 1 | 1 |
| Resolution, (Å) | 50–2.4 (2.44–2.4) | 50–3.5 (3.69–3.5) | 50–2.4 (2.44–2.4) | 50–2.85 (2.9–2.85) | 50–2.19 (2.23–2.19) | 50–2.6 (2.64–2.6) |
| # of reflections | ||||||
| Total | 49,666 | 11,650 | 61,106 | 34,488 | 86,920 | 39,768 |
| Unique | 13,796 | 4021 | 14,904 | 7185 | 20,214 | 10,748 |
| Rmerge a, % | 9.1 (57.9) | 22.3 (113) | 10.6 (80.1) | 12.1 (104) | 17.2 (66.5) | 15.4 (98.3) |
| Rpim b, % | 5.1 (32.3) | 14.7 (72.9) | 5.8 (71.1) | 8.2 (77.4) | 9.7 (36.4) | 9.0 (65.0) |
| CC1/2 c | 0.98 (0.71) | 0.97 (0.33) | 1.0 (0.50) | 1.0 (0.42) | 1.0 (0.65) | 0.94 (0.60) |
| I/σ | 12.3 (1.3) | 2.8 (0.6) | 14.2 (0.8) | 12.5 (0.8) | 25.5 (2.1) | 17.3 (1.1) |
| Completeness, % | 88.3 (78.2) | 80.6 (83.9) | 97.8 (86.4) | 83.3 (78.0) | 97.4 (85.0) | 86.1 (90.2) |
| Redundancy | 3.6 (3.7) | 2.9 (3.0) | 4.1 (2.7) | 4.8 (4.2) | 4.3 (3.8) | 3.7 (3.1) |
| Refinement Statistics | ||||||
| Resolution, Å | 50.0–2.4 | 50.0–3.5 | 50.0–2.4 | 50.0–2.85 | 50.0–2.19 | 50.0–2.6 |
| R d % | 22.1 | 25.6 | 24.0 | 24.2 | 20.8 | 22.1 |
| Rfree e, % | 27.4 | 30.7 | 26.7 | 29.6 | 25.5 | 27.4 |
| # of atoms | ||||||
| Protein | 2,668 | 2,640 | 2,665 | 2,675 | 2,683 | 2,668 |
| Water | 32 | – | 23 | – | 64 | 2 |
| Ligand/Ion | 212 | 184 | 211 | 186 | 193 | 186 |
| Overall B value (Å)2 | ||||||
| Protein | 52 | 82 | 62 | 73 | 53 | 85 |
| Water | 48 | – | 56 | – | 49 | 74 |
| Ligand/Ion | 70 | 96 | 74 | 86 | 63 | 102 |
| RMSD f | ||||||
| Bond lengths, Å | 0.005 | 0.004 | 0.006 | 0.009 | 0.007 | 0.012 |
| Bond angles, ° | 1.0 | 0.79 | 0.98 | 1.3 | 1.0 | 1.4 |
| Ramachandran g | ||||||
| Favored, % | 94.0 | 90.2 | 94.0 | 89.5 | 95.0 | 92.5 |
| Allowed, % | 4.2 | 7.4 | 5.4 | 9.0 | 4.7 | 7.2 |
| Outliers, % | 1.8 | 2.4 | 0.6 | 1.5 | 0.3 | 0.3 |
| PDB ID | 8GD0 | 8GJT | 8GCZ | 8GD1 | 8GD3 | 8GD5 |
| VSV-G Pseudovirus | HIV-1CH58TF | HIV-1JRFL | HIV-1AD8 | |
|---|---|---|---|---|
| DMSO | >100 | >100 | >100 | >100 |
| BNM-III-170 | >100 | 0.033 ± 0.01 | 13.49 ± 6.65 | 3.75 ± 1.3 |
| MCG-IV-210 | >100 | 3.46 ± 1.06 | >100 | >100 |
| TFH-I-070-A6 | >100 | 0.45 ± 0.09 | 67.24 ± 30 | 9.59 ± 3.4 |
| TFH-I-116-D1 | >100 | 0.065 ± 0.01 | 43.82 ± 16.6 | 4.8 ± 1.7 |
| ZXC-I-090 | >100 | 2.63 ± 0.5 | >100 | >100 |
| ZXC-I-092 | >100 | 0.75 ± 0.12 | 54.22 ± 22.8 | 34.05 ± 13.8 |
| DL-I-101 | >100 | 0.16 ± 0.03 | >100 | 10.21 ± 3.1 |
| DL-I-102 | >100 | 0.66 ± 0.25 | >100 | 87.02 ± 39 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, S.; Tolbert, W.D.; Zhu, H.; Lee, D.; Marchitto, L.; Higgins, T.; Zhao, X.; Nguyen, D.; Sherburn, R.; Richard, J.; et al. Piperidine CD4-Mimetic Compounds Expose Vulnerable Env Epitopes Sensitizing HIV-1-Infected Cells to ADCC. Viruses 2023, 15, 1185. https://doi.org/10.3390/v15051185
Ding S, Tolbert WD, Zhu H, Lee D, Marchitto L, Higgins T, Zhao X, Nguyen D, Sherburn R, Richard J, et al. Piperidine CD4-Mimetic Compounds Expose Vulnerable Env Epitopes Sensitizing HIV-1-Infected Cells to ADCC. Viruses. 2023; 15(5):1185. https://doi.org/10.3390/v15051185
Chicago/Turabian StyleDing, Shilei, William D. Tolbert, Huile Zhu, Daniel Lee, Lorie Marchitto, Tyler Higgins, Xuchen Zhao, Dung Nguyen, Rebekah Sherburn, Jonathan Richard, and et al. 2023. "Piperidine CD4-Mimetic Compounds Expose Vulnerable Env Epitopes Sensitizing HIV-1-Infected Cells to ADCC" Viruses 15, no. 5: 1185. https://doi.org/10.3390/v15051185
APA StyleDing, S., Tolbert, W. D., Zhu, H., Lee, D., Marchitto, L., Higgins, T., Zhao, X., Nguyen, D., Sherburn, R., Richard, J., Gendron-Lepage, G., Medjahed, H., Mohammadi, M., Abrams, C., Pazgier, M., Smith, A. B., III, & Finzi, A. (2023). Piperidine CD4-Mimetic Compounds Expose Vulnerable Env Epitopes Sensitizing HIV-1-Infected Cells to ADCC. Viruses, 15(5), 1185. https://doi.org/10.3390/v15051185