

Introductions of Human-Origin Seasonal H3N2, H1N2 and Pre-2009 H1N1 Influenza Viruses to Swine in Brazil

 , , , , and
, , , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Preparation and Sequencing
2.2. Data Set Construction
2.3. Phylogenetic Analysis
3. Results
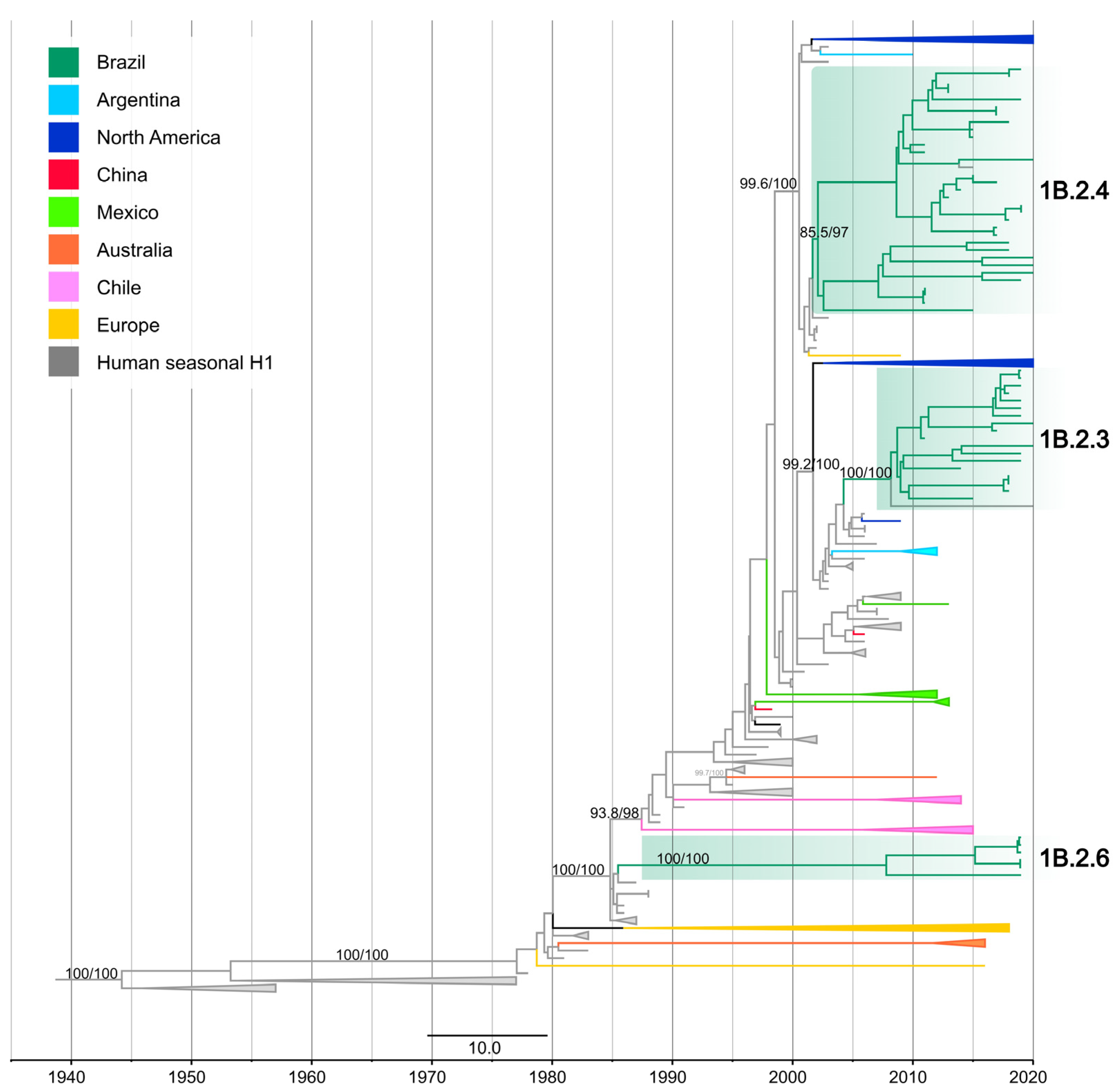
3.1. New Human-Origin H1N2/H1N1 in Swine
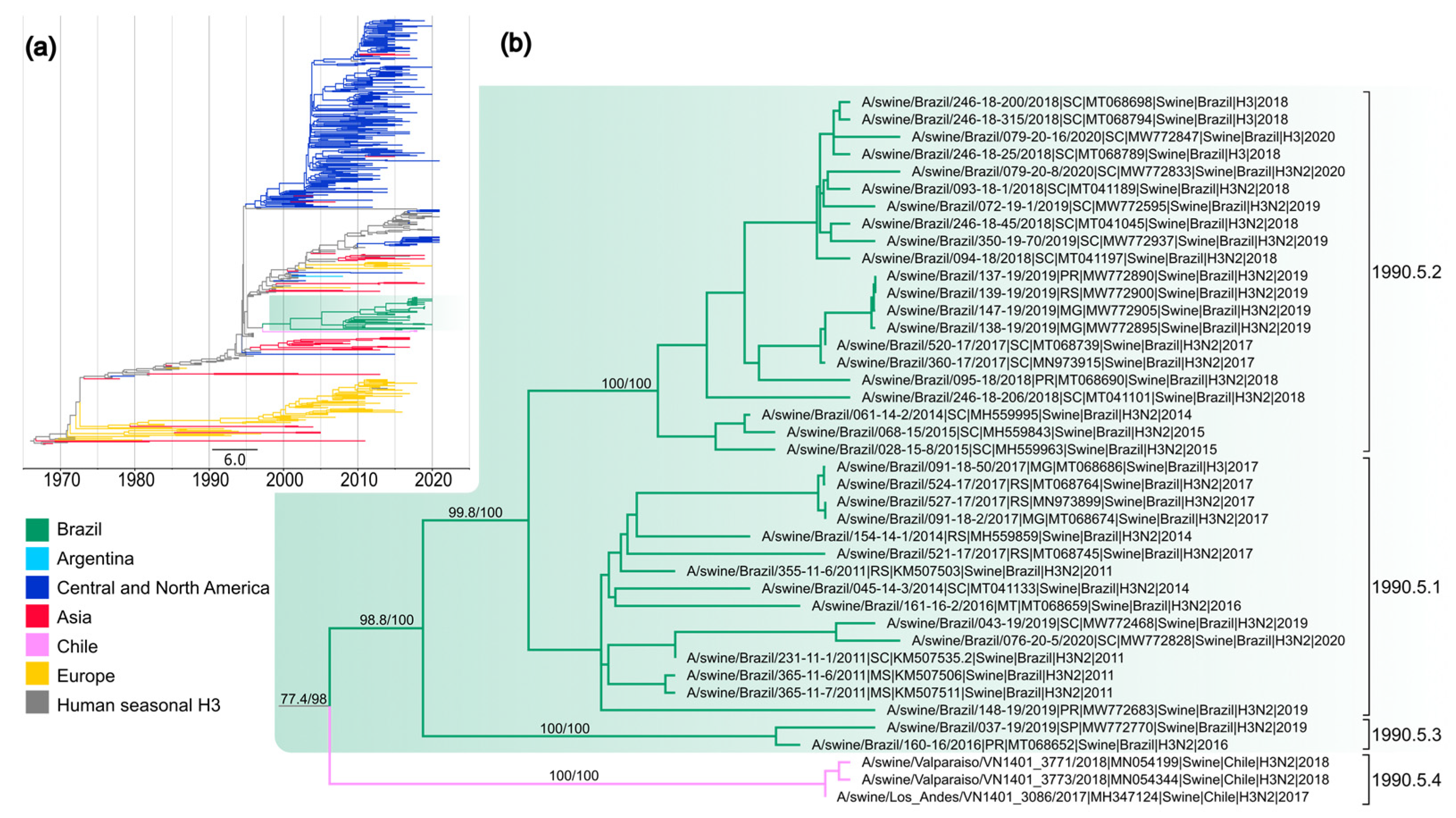
3.2. Sustained Transmission in Swine after a Single Introduction of Human H3 Segment
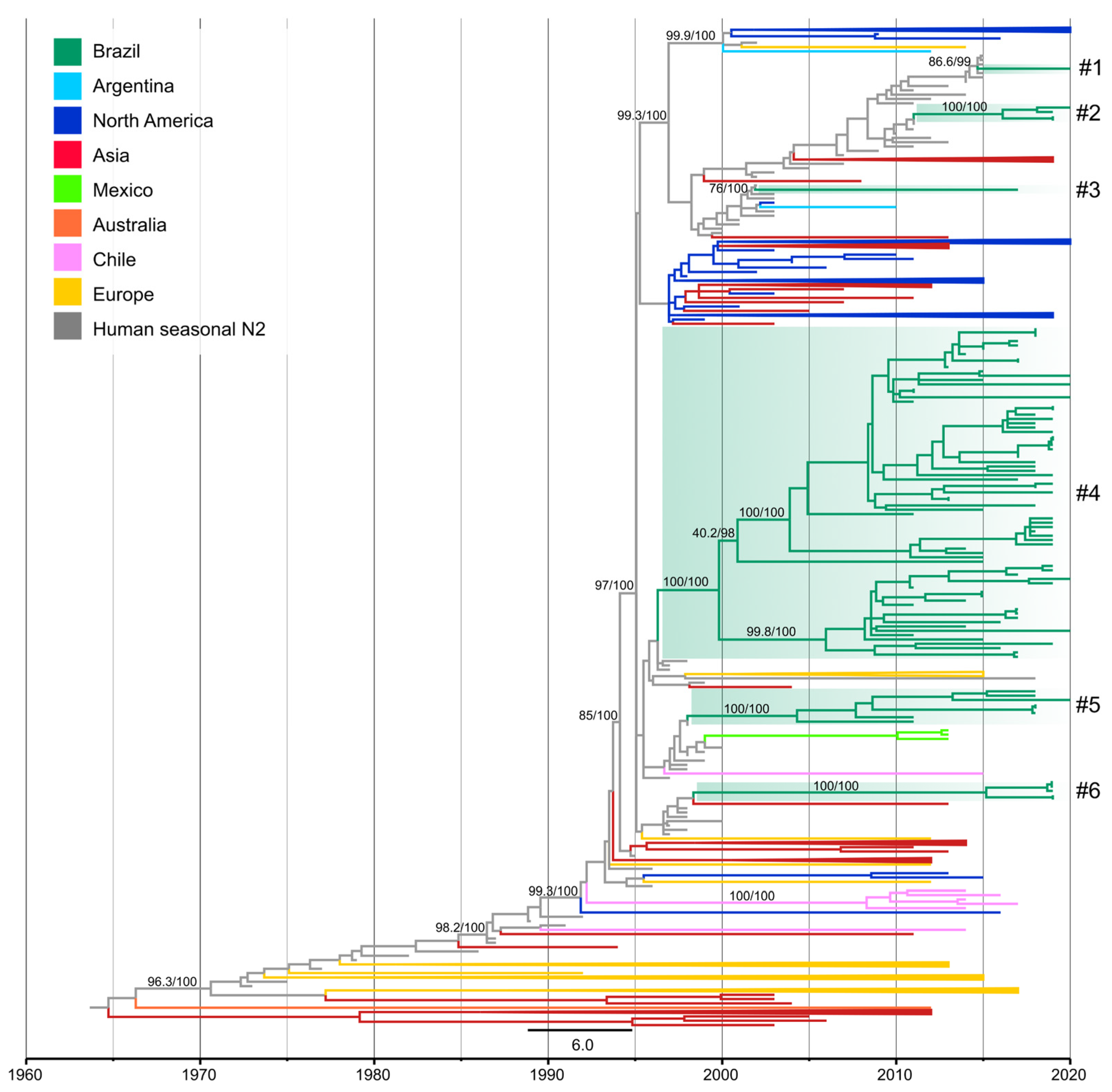
3.3. Novel Introductions of Human N2 Segments in Swine
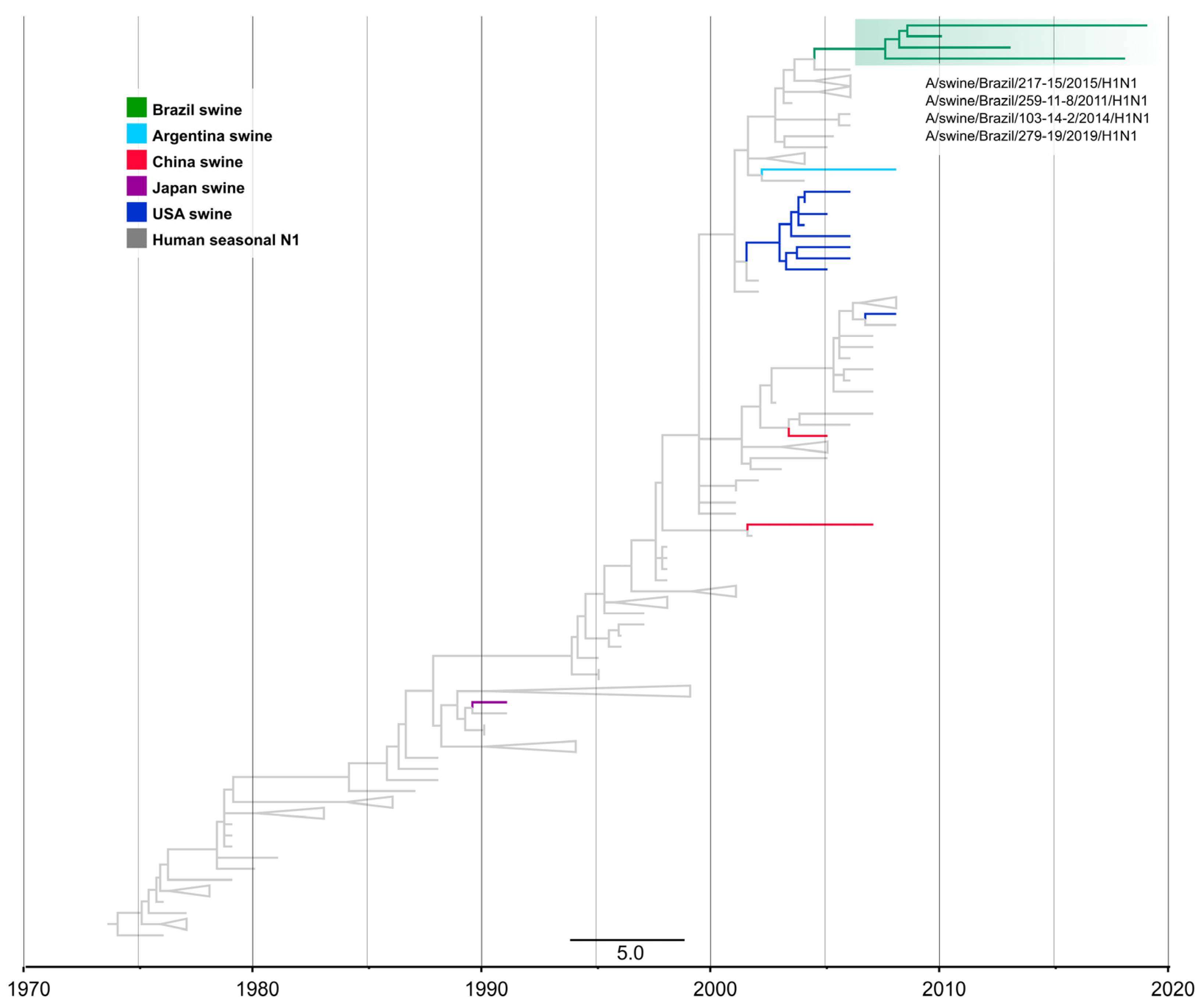
3.4. One Introduction of Pre-2009 Human IAV N1 Segment Paired with 1B HA Genes in Swine in Brazil
3.5. Reassortment with H1N1pdm09 Viruses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Garten, R.J.; Davis, C.T.; Russell, C.A.; Shu, B.; Lindstrom, S.; Balish, A.; Sessions, W.M.; Xu, X.; Skepner, E.; Deyde, V.; et al. Antigenic and Genetic Characteristics of Swine-Origin 2009 A(H1N1) Influenza Viruses Circulating in Humans. Science 2009, 325, 197–201. [Google Scholar] [CrossRef]
- Anderson, T.K.; Chang, J.; Arendsee, Z.W.; Venkatesh, D.; Souza, C.K.; Kimble, J.B.; Lewis, N.S.; Davis, C.T.; Vincent, A.L. Swine Influenza a Viruses and the Tangled Relationship with Humans. Cold Spring Harb. Perspect Med. 2021, 11, a038737. [Google Scholar] [CrossRef]
- Powell, J.D.; Abente, E.J.; Chang, J.; Souza, C.K.; Rajao, D.S.; Anderson, T.K.; Zeller, M.A.; Gauger, P.C.; Lewis, N.S.; Vincent, A.L. Characterization of Contemporary 2010.1 H3N2 Swine Influenza A Viruses Circulating in United States Pigs. Virology 2021, 553, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Nelson, M.I.; Gramer, M.R.; Vincent, A.L.; Holmes, E.C. Global Transmission of Influenza Viruses from Humans to Swine. J. Gen. Virol. 2012, 93, 2195–2203. [Google Scholar] [CrossRef]
- Nelson, M.I.; Vincent, A.L. Reverse Zoonosis of Influenza to Swine: New Perspectives on the Human-Animal Interface. Trends. Microbiol. 2015, 23, 142–153. [Google Scholar] [CrossRef]
- Sharma, A.; Zeller, M.A.; Souza, C.K.; Anderson, T.K.; Vincent, A.L.; Harmon, K.; Li, G.; Zhang, J.; Gauger, P.C. Characterization of a 2016-2017 Human Seasonal H3 Influenza A Virus Spillover Now Endemic to U.S. Swine. mSphere 2022, 7, e00809-21. [Google Scholar] [CrossRef]
- Rajao, D.S.; Vincent, A.L.; Perez, D.R. Adaptation of Human Influenza Viruses to Swine. Front. Vet. Sci. 2019, 5, 347. [Google Scholar] [CrossRef] [PubMed]
- Anderson, T.K.; Macken, C.A.; Lewis, N.S.; Scheuermann, R.H.; van Reeth, K.; Brown, I.H.; Swenson, S.L.; Simon, G.; Saito, T.; Berhane, Y.; et al. A Phylogeny-Based Global Nomenclature System and Automated Annotation Tool for H1 Hemagglutinin Genes from Swine Influenza A Viruses. mSphere 2016, 1, e00275-16. [Google Scholar] [CrossRef]
- ABPA. Relatório Anual, Associação Brasileira de Proteína Animal (ABPA). 2022. Available online: https://abpa-br.org/mercados/#relatorios (accessed on 30 November 2022).
- Cunha, R.G.; Vinha, V.R.; Passos, W.D. Isolation of a Strain of Myxovirus Influenzae-A Suis from Swine Slaughtered in Rio de Janeiro. Rev. Bras. Biol. 1978, 38, 13–17. [Google Scholar]
- Brentano, L.; Reis Ciacci-Zanella, J.; Mores, N.; Piffer, I.A. Levantamento Soroepidemiológico Para Coronavírus Respiratório e Da Gastroenterite Transmissível e Dos Vírus de Influenza H3N2 e H1N1 Em Rebanhos Suínos No Brasil. Comunicado Técnico 2002, 1, 1–6. [Google Scholar]
- Caron, L.F.; Joineau, M.E.G.; Santin, E.; Richartz, R.R.T.; Patricio, M.A.C.; Soccol, V.T. Seroprevalence of H3N2 Influenza a Virus in Pigs from Paraná (South Brazil): Interference of the Animal Management and Climatic Conditions. Virus Rev. Res. 2010, 15, 3. [Google Scholar] [CrossRef][Green Version]
- Rajão, D.S.; Alves, F.; del Puerto, H.L.; Braz, G.F.; Oliveira, F.G.; Ciacci-Zanella, J.R.; Schaefer, R.; dos Reis, J.K.P.; Guedes, R.M.C.; Lia, Z.; et al. Serological Evidence of Swine Influenza in Brazil. Influenza Other Respir. Viruses 2012, 7, 109–112. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, R.; Zanella, J.R.C.; Brentano, L.; Vincent, A.L.; Ritterbusch, G.A.; Silveira, S.; Caron, L.; Mores, N. Isolation and Characterization of a Pandemic H1N1 Influenza Virus in Pigs in Brazil. Pesq. Vet. Bras. 2011, 31, 761–767. [Google Scholar] [CrossRef]
- Ciacci-Zanella, J.R.; Schaefer, R.; Gava, D.; Haach, V.; Cantão, M.E.; Coldebella, A. Influenza A Virus Infection in Brazilian Swine Herds Following the Introduction of Pandemic 2009 H1N1. Vet. Microbiol. 2015, 180, 118–122. [Google Scholar] [CrossRef]
- Schaefer, R.; Rech, R.R.; Gava, D.; Cantão, M.E.; da Silva, M.C.; Silveira, S.; Zanella, J.R.C. A Human-like H1N2 Influenza Virus Detected during an Outbreak of Acute Respiratory Disease in Swine in Brazil. Arch. Virol. 2015, 160, 29–38. [Google Scholar] [CrossRef]
- Nelson, M.I.; Schaefer, R.; Gava, D.; Cantão, M.E.; Ciacci-Zanella, J.R. Influenza a Viruses of Human Origin in Swine, Brazil. Emerg. Infect. Dis. 2015, 21, 1339–1347. [Google Scholar] [CrossRef]
- Haach, V.; Gava, D.; Mauricio, E.C.; Franco, A.C.; Schaefer, R. One-Step Multiplex RT-QPCR for the Detection and Subtyping of Influenza A Virus in Swine in Brazil. J. Virol. Methods 2019, 269, 43–48. [Google Scholar] [CrossRef]
- Haach, V.; Gava, D.; Cantão, M.E.; Schaefer, R. Evaluation of Two Multiplex RT-PCR Assays for Detection and Subtype Differentiation of Brazilian Swine Influenza Viruses. Braz. J. Microbiol. 2020, 51, 1447–1451. [Google Scholar] [CrossRef]
- Rech, R.R.; Gava, D.; Silva, M.C.; Fernandes, L.T.; Haach, V.; Ciacci-Zanella, J.R.; Schaefer, R. Porcine Respiratory Disease Complex after the Introduction of H1N1/2009 Influenza Virus in Brazil. Zoonoses Public Health 2018, 65, e155–e161. [Google Scholar] [CrossRef]
- Zhang, J.; Harmon, K.M. RNA Extraction from Swine Samples and Detection of Influenza A Virus in Swine by Real-Time RT-PCR. In Animal Influenza Virus. Methods in Molecular Biology; Spackman, E., Ed.; Springer: New York, NY, USA, 2020; Volume 2123, pp. 295–310. [Google Scholar]
- Zhang, J.; Gauger, P.C. Isolation of Swine Influenza A Virus in Cell Cultures and Embryonated Chicken Eggs. In Animal Influenza Virus. Methods in Molecular Biology; Spackman, E., Ed.; Springer: New York, NY, USA, 2020; Volume 2123, pp. 281–294. [Google Scholar]
- Zhang, Y.; Aevermann, B.D.; Anderson, T.K.; Burke, D.F.; Dauphin, G.; Gu, Z.; He, S.; Kumar, S.; Larsen, C.N.; Lee, A.J.; et al. Influenza Research Database: An Integrated Bioinformatics Resource for Influenza Virus Research. Nucleic. Acids Res. 2017, 45, D466–D474. [Google Scholar] [CrossRef]
- Elbe, S.; Buckland-Merrett, G. Data, Disease and Diplomacy: GISAID’s Innovative Contribution to Global Health. Global Challenges 2017, 1, 33–46. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Godzik, A. Cd-Hit: A Fast Program for Clustering and Comparing Large Sets of Protein or Nucleotide Sequences. Bioinformatics 2006, 22, 1658–1659. [Google Scholar] [CrossRef] [PubMed]
- Junqueira, D.M.; Tochetto, C.; Anderson, T.K.; Gava, D.; Haach, V.; Cantão, M.E.; Vincent Baker, A.L.; Schaefer, R. Human-to-Swine Spillover and Onward Transmission of H1N1pdm09 in Brazil; Embrapa Suínos e Aves: Concórdia, SC, Brazil, 2023; manuscript in preparation. [Google Scholar]
- Kaplan, B.S.; Anderson, T.K.; Chang, J.; Santos, J.; Perez, D.; Lewis, N.; Vincent, A.L. Evolution and Antigenic Advancement of N2 Neuraminidase of Swine Influenza A Viruses Circulating in the United States Following Two Separate Introductions from Human Seasonal Viruses. J. Virol. 2021, 95, 632–653. [Google Scholar] [CrossRef] [PubMed]
- Zeller, M.A.; Chang, J.; Vincent, A.L.; Gauger, P.C.; Anderson, T.K. Spatial and Temporal Coevolution of N2 Neuraminidase and H1 and H3 Hemagglutinin Genes of Influenza A Virus in US Swine. Virus Evol. 2021, 7, veab090. [Google Scholar] [CrossRef]
- Rambaut, A.; Lam, T.T.; Carvalho, L.M.; Pybus, O.G. Exploring the Temporal Structure of Heterochronous Sequences Using TempEst (Formerly Path-O-Gen). Virus Evol. 2016, 2, vew007. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Larsson, A. AliView: A Fast and Lightweight Alignment Viewer and Editor for Large Datasets. Bioinformatics 2014, 30, 3276–3278. [Google Scholar] [CrossRef]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New Algorithms and Methods to Estimate Maximum-Likelihood Phylogenies: Assessing the Performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Sagulenko, P.; Puller, V.; Neher, R.A. TreeTime: Maximum-Likelihood Phylodynamic Analysis. Virus Evol. 2018, 4, vex042. [Google Scholar] [CrossRef] [PubMed]
- Inkscape Draw Freely. Available online: https://inkscape.org/ (accessed on 8 February 2023).
- Nelson, M.I.; Wentworth, D.E.; Culhane, M.R.; Vincent, A.L.; Viboud, C.; LaPointe, M.P.; Lin, X.; Holmes, E.C.; Detmer, S.E. Introductions and Evolution of Human-Origin Seasonal Influenza A Viruses in Multinational Swine Populations. J. Virol. 2014, 88, 10110–10119. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.; Anderson, T.K.; Zeller, M.A.; Gauger, P.C.; Vincent, A.L. OctoFLU: Automated Classification for the Evolutionary Origin of Influenza A Virus Gene Sequences Detected in U.S. Swine. Microbiol. Resour. Announc. 2019, 8, e00673-19. [Google Scholar] [CrossRef]
- Biondo, N.; Schaefer, R.; Gava, D.; Cantão, M.E.; Silveira, S.; Mores, M.A.Z.; Ciacci-Zanella, J.R.; Barcellos, D.E.S.N. Genomic Analysis of Influenza A Virus from Captive Wild Boars in Brazil Reveals a Human-like H1N2 Influenza Virus. Vet Microbiol 2014, 168, 34–40. [Google Scholar] [CrossRef]
- Schmidt, C.; Cibulski, S.P.; Muterle Varela, A.P.; Mengue Scheffer, C.; Wendlant, A.; Quoos Mayer, F.; Lopes de Almeida, L.; Franco, A.C.; Roehe, P.M. Full-Genome Sequence of a Reassortant H1N2 Influenza A Virus Isolated from Pigs in Brazil. Genome Announc 2014, 2, e01319-14. [Google Scholar] [CrossRef]
- Lopes, S.; Anderson, T.K.; Schaefer, R.; Tochetto, C.; Gava, D.; Cantão, M.E.; Ciacci-Zanella, J.R.; Vincent, A.L.; Lewis, N.S. Antigenic and Genetic Diversity of H1 and H3 Influenza A Viruses in Swine in Brazil; Department of Pathobiology and Population Sciences, The Royal Veterinary College, University of London: Hertfordshire, UK, 2023; manuscript in preparation. [Google Scholar]
- Nelson, M.I.; Lemey, P.; Tan, Y.; Vincent, A.; Lam, T.T.-Y.; Detmer, S.; Viboud, C.; Suchard, M.A.; Rambaut, A.; Holmes, E.C.; et al. Spatial Dynamics of Human-Origin H1 Influenza A Virus in North American Swine. PLoS Pathog. 2011, 7, e1002077. [Google Scholar] [CrossRef]
- Lewis, N.S.; Russell, C.A.; Langat, P.; Anderson, T.K.; Berger, K.; Bielejec, F.; Burke, D.F.; Dudas, G.; Fonville, J.M.; Fouchier, R.A.; et al. The Global Antigenic Diversity of Swine Influenza A Viruses. Elife 2016, 5, e12217. [Google Scholar] [CrossRef]
- Bowman, A.S.; Nelson, S.W.; Page, S.L.; Nolting, J.M.; Killian, M.L.; Sreevatsan, S.; Slemons, R.D. Swine-to-Human Transmission of Influenza A(H3N2) Virus at Agricultural Fairs, Ohio, USA, 2012. Emerg. Infect Dis. 2014, 20, 1472–1480. [Google Scholar] [CrossRef]
- Bastien, N.; Antonishyn, N.A.; Brandt, K.; Wong, C.E.; Chokani, K.; Vegh, N.; Horsman, G.B.; Tyler, S.; Graham, M.R.; Plummer, F.A.; et al. Human Infection with a Triple-Reassortant Swine Influenza A(H1N1) Virus Containing the Hemagglutinin and Neuraminidase Genes of Seasonal Influenza Virus. J. Infect Dis. 2010, 201, 1178–1182. [Google Scholar] [CrossRef]
- Xie, J.-F.; Zhang, Y.-H.; Zhao, L.; Xiu, W.-Q.; Chen, H.-B.; Lin, Q.; Weng, Y.-W.; Zheng, K.-C. Emergence of Eurasian Avian-Like Swine Influenza A (H1N1) Virus from an Adult Case in Fujian Province, China. Virol. Sin. 2018, 33, 282–286. [Google Scholar] [CrossRef]
- Lu, J.; Yi, L.; Jing, Y.; Tan, H.; Mai, W.; Song, Y.; Zou, L.; Liang, L.; Xiao, H.; Kang, M.; et al. A Human Infection with a Novel Reassortant H3N2 Swine Virus in China. J. Infect. 2019, 79, 174–187. [Google Scholar] [CrossRef]
- Parys, A.; Vandoorn, E.; King, J.; Graaf, A.; Pohlmann, A.; Beer, M.; Harder, T.; van Reeth, K. Human Infection with Eurasian Avian-Like Swine Influenza A(H1N1) Virus, the Netherlands, September 2019. Emerg. Infect Dis. 2021, 27, 939–943. [Google Scholar] [CrossRef]
- Resende, P.C.; Born, P.S.; Matos, A.R.; Motta, F.C.; Caetano, B.C.; Debur, M.d.C.; Riediger, I.N.; Brown, D.; Siqueira, M.M. Whole-Genome Characterization of a Novel Human Influenza A(H1N2) Virus Variant, Brazil. Emerg. Infect Dis. 2017, 23, 152–154. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Influenza A(H1N2)—Brazil. Available online: https://www.who.int/emergencies/disease-outbreak-news/item/2020-DON286 (accessed on 8 February 2023).
- Chepkwony, S.; Parys, A.; Vandoorn, E.; Stadejek, W.; Xie, J.; King, J.; Graaf, A.; Pohlmann, A.; Beer, M.; Harder, T.; et al. Genetic and Antigenic Evolution of H1 Swine Influenza A Viruses Isolated in Belgium and the Netherlands from 2014 through 2019. Sci. Rep. 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Komadina, N.; McVernon, J.; Hall, R.; Leder, K. A Historical Perspective of Influenza A(H1N2) Virus. Emerg. Infect Dis. 2014, 20, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Hufnagel, D.E.; Young, K.M.; Arendsee, Z.; Claire Gay, L.; Joaquin, C.; Rajão, D.S.; Perez, D.R.; Vincent Baker, A.L.; Anderson, T.K. Characterizing a Century of Genetic Diversity and Contemporary Antigenic Diversity of N1 Neuraminidase in IAV from North American Swine. Biorxiv 2022. [Google Scholar] [CrossRef]
- Haach, V.; Bastos, A.P.A.; Gava, D.; Fonseca, F.N.; Morés, M.A.Z.; Coldebella, A.; Franco, A.C.; Schaefer, R. Immunogenicity of a Trivalent Virosome-Based Influenza Vaccine in Pigs. Vaccine 2023. manuscript submmited. [Google Scholar]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tochetto, C.; Junqueira, D.M.; Anderson, T.K.; Gava, D.; Haach, V.; Cantão, M.E.; Vincent Baker, A.L.; Schaefer, R. Introductions of Human-Origin Seasonal H3N2, H1N2 and Pre-2009 H1N1 Influenza Viruses to Swine in Brazil. Viruses 2023, 15, 576. https://doi.org/10.3390/v15020576
Tochetto C, Junqueira DM, Anderson TK, Gava D, Haach V, Cantão ME, Vincent Baker AL, Schaefer R. Introductions of Human-Origin Seasonal H3N2, H1N2 and Pre-2009 H1N1 Influenza Viruses to Swine in Brazil. Viruses. 2023; 15(2):576. https://doi.org/10.3390/v15020576
Chicago/Turabian StyleTochetto, Caroline, Dennis M. Junqueira, Tavis K. Anderson, Danielle Gava, Vanessa Haach, Mauricio E. Cantão, Amy L. Vincent Baker, and Rejane Schaefer. 2023. "Introductions of Human-Origin Seasonal H3N2, H1N2 and Pre-2009 H1N1 Influenza Viruses to Swine in Brazil" Viruses 15, no. 2: 576. https://doi.org/10.3390/v15020576
APA StyleTochetto, C., Junqueira, D. M., Anderson, T. K., Gava, D., Haach, V., Cantão, M. E., Vincent Baker, A. L., & Schaefer, R. (2023). Introductions of Human-Origin Seasonal H3N2, H1N2 and Pre-2009 H1N1 Influenza Viruses to Swine in Brazil. Viruses, 15(2), 576. https://doi.org/10.3390/v15020576