
Non-Lethal Detection of Ranavirus in Fish
 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
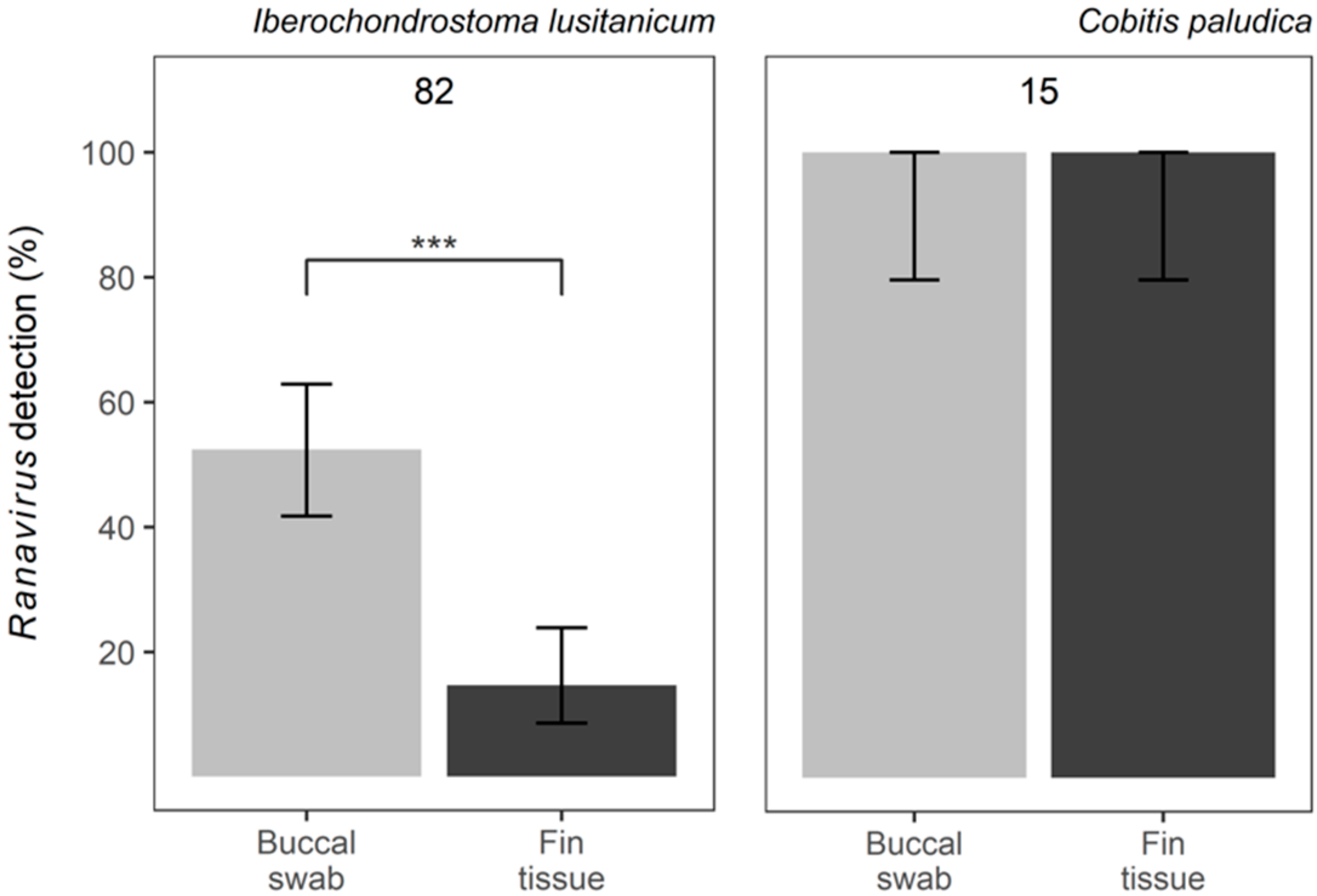
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Balcombe, J.P.; Barnard, N.D.; Sandusky, C. Laboratory routines cause animal stress. J. Am. Assoc. Lab. Anim. Sci. 2004, 43, 42–51. [Google Scholar]
- Davis, A.K.; Maerz, J.C. Assessing stress levels of captive-reared amphibians with hematological data: Implications for conservation initiatives. J. Herpetol. 2011, 45, 40–44. [Google Scholar] [CrossRef]
- Harikrishnan, V.; Hansen, A.K.; Abelson, K.S.; Sørensen, D.B. A comparison of various methods of blood sampling in mice and rats: Effects on animal welfare. Lab. Anim. 2018, 52, 253–264. [Google Scholar] [CrossRef]
- Sneddon, L.U.; Halsey, L.G.; Bury, N.R. Considering aspects of the 3Rs principles within experimental animal biology. J. Exp. Biol. 2017, 220, 3007–3016. [Google Scholar] [CrossRef] [PubMed]
- WOAH. Chapter 2.1.3. Infection with Ranavirus. In Manual of Diagnostic Tests for Aquatic Animals 2021; WOAH: Paris, France, 2021; pp. 69–88. [Google Scholar]
- Price, S.J.; Garner, T.W.J.; Nichols, R.A.; Balloux, F.; Ayres, C.; Mora-Cabello de Alba, A.; Bosch, J. Collapse of amphibian communities due to an introduced ranavirus. Curr. Biol. 2014, 24, 2586–2591. [Google Scholar] [CrossRef]
- Rosa, G.M.; Sabino-Pinto, J.; Laurentino, T.G.; Martel, A.; Pasmans, F.; Rebelo, R.; Griffiths, R.A.; Stöhr, A.C.; Marschang, R.E.; Price, S.J.; et al. Impact of asynchronous emergence of two lethal pathogens on amphibian assemblages. Sci. Rep. 2017, 7, 43260. [Google Scholar] [CrossRef]
- Rosa, G.M.; Botto, G.A.; Mitra, A.T.; de Almeida, J.S.; Hofmann, M.; Leung, W.T.M.; de Matos, A.P.A.; Caeiro, M.F.; Froufe, E.; Loureiro, A.; et al. Invasive fish disrupt host-pathogen dynamics leading to amphibian declines. Biol. Conserv. 2022, 276, 109785. [Google Scholar] [CrossRef]
- Mao, J.; Green, D.E.; Fellers, G.; Chinchar, V.G. Molecular characterization of iridoviruses isolated from sympatric amphibians and fish. Virus. Res. 1999, 63, 45–52. [Google Scholar] [CrossRef]
- Teacher, A.G.F.; Cunningham, A.A.; Garner, T.W.J. Assessing the long-term impact of Ranavirus infection in wild common frog populations. Anim. Conserv. 2010, 13, 514–522. [Google Scholar] [CrossRef]
- Waltzek, T.B.; Miller, D.L.; Gray, M.J.; Drecktrah, B.; Briggler, J.T.; MacConnell, B.; Hudson, C.; Hopper, L.; Friary, J.; Yun, S.C.; et al. New disease records for hatchery-reared sturgeon, I, Expansion of frog virus 3 host range into Scaphirhynchus albus. Dis. Aquat. Organ. 2014, 111, 219–227. [Google Scholar] [CrossRef]
- Duffus, A.L.J.; Waltzek, T.B.; Stöhr, A.C.; Allender, M.C.; Gotesman, M.; Whittington, R.J.; Hick, P.; Hines, M.K.; Marschang, R.E. Distribution and host range of ranaviruses. In Ranaviruses: Lethal Pathogens of Ectothermic Vertebrates; Gray, M.J., Chinchar, V.G., Eds.; Springer: Cham, Switzerland, 2015; pp. 9–57. [Google Scholar]
- Adamovicz, L.; Allender, M.C.; Archer, G.; Rzadkowska, M.; Boers, K.; Phillips, C.; Driskell, E.; Kinsel, M.J.; Chu, C. Investigation of multiple mortality events in eastern box turtles (Terrapene carolina carolina). PLoS ONE 2018, 13, e0195617. [Google Scholar] [CrossRef]
- McKenzie, C.M.; Piczak, M.L.; Snyman, H.N.; Joseph, T.; Theijin, C.; Chow-Fraser, P.; Jardineet, C.M. First report of ranavirus mortality in a common snapping turtle Chelydra serpentina. Dis. Aquat. Organ 2019, 132, 221–227. [Google Scholar] [CrossRef]
- Whittington, R.J.; Philbey, A.; Reddacliff, G.L.; Macgown, A.R. Epidemiology of epizootic haematopoietic necrosis virus (EHNV) infection in farmed rainbow trout, Oncorhynchus mykiss (Walbaum): Findings based on virus isolation, antigen capture ELISA and serology. J. Fish. Dis. 1994, 17, 205–218. [Google Scholar] [CrossRef]
- Whittington, R.J.; Becker, J.A.; Dennis, M.M. Iridovirus infections in finfish-critical review with emphasis on ranaviruses. J. Fish. Dis. 2010, 33, 95–122. [Google Scholar] [CrossRef] [PubMed]
- George, M.R.; John, K.R.; Mansoor, M.M.; Saravanakumar, R.; Sundar, P.; Pradeep, V. Isolation and characterization of a ranavirus from koi, Cyprinus carpio L., experiencing mass mortalities in India. J. Fish. Dis 2015, 38, 389–403. [Google Scholar] [CrossRef] [PubMed]
- Picco, A.M.; Brunner, J.L.; Collins, J.P. Susceptibility of the Endangered California Tiger Salamander, Ambystoma californiense, to Ranavirus Infection. J. Wildl. Dis. 2007, 43, 286–290. [Google Scholar] [CrossRef]
- St-Amour, V.; Garner, T.W.J.; Schulte-Hostedde, A.I.; Lesbarrères, D. Effects of Two Amphibian Pathogens on the Developmental Stability of Green Frogs. Conserv. Biol. 2010, 24, 788–794. [Google Scholar] [CrossRef]
- Gray, M.J.; Duffus, A.L.J.; Haman, K.H.; Harris, R.N.; Allender, M.C.; Thompson, T.A.; Christman, M.R.; Sacerdote-Velat, A.; Sprague, L.A.; Williams, J.M.; et al. Pathogen surveillance in herpetofaunal populations: Guidance on study design, sample collection, biosecurity, and intervention strategies. Herpetol. Rev. 2017, 48, 334–351. [Google Scholar]
- Price, S.J.; Wadia, A.; Wright, O.N.; Leung, W.T.M.; Cunningham, A.A.; Lawson, B. Screening of a long-term sample set reveals two Ranavirus lineages in British herpetofauna. PLoS ONE 2017, 12, e0184768. [Google Scholar] [CrossRef]
- St-Amour, V.; Lesbarrères, D. Genetic evidence of Ranavirus in toe clips: An alternative to lethal sampling methods. Conserv. Genet. 2007, 8, 1247–1250. [Google Scholar] [CrossRef]
- Kurobe, T.; Kwak, K.T.; MacConnell, E.; McDowell, T.S.; Mardones, F.O.; Hedrick, R.P. Development of PCR assays to detect iridovirus infections among captive and wild populations of Missouri River sturgeon. Dis. Aquat. Organ. 2010, 93, 31–42. [Google Scholar] [CrossRef]
- Greer, A.L.; Collins, J.P. Sensitivity of a diagnostic test for amphibian ranavirus varies with sampling protocol. J. Wildl. Dis. 2007, 43, 525–532. [Google Scholar] [CrossRef]
- Gray, M.J.; Miller, D.L.; Hoverman, J.T. Reliability of non-lethal surveillance methods for detecting ranavirus infection. Dis. Aquat. Organ 2012, 99, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Deakin, A.G.; Buckley, J.; AlZu’bi, H.S.; Cossins, A.R.; Spencer, J.W.; Al’Nuaimy, W.; Young, I.S.; Thomson, J.S.; Sneddon, L.U. Automated monitoring of behaviour in zebrafish after invasive procedures. Sci. Rep. 2019, 9, 9042. [Google Scholar] [CrossRef]
- Tilley, C.A.; Carreño Gutierrez, H.; Sebire, M.; Obasaju, O.; Reichmann, F.; Katsiadaki, I.; Barber, I.; Norton, W.H.J. Skin swabbing is a refined technique to collect DNA from model fish species. Sci. Rep. 2020, 10, 18212. [Google Scholar] [CrossRef] [PubMed]
- Kidd, A.; Reid, S.; Wilson, C. Non-invasive DNA sampling from small-bodied species at risk. Ontario Ministry of Natural Resources, 2014; 6 pp + appendices. [Google Scholar]
- Breacker, C.; Barber, I.; Norton, W.H.J.; McDearmid, J.R.; Tilley, C.A. A low-cost method of skin swabbing for the collection of DNA samples from small laboratory fish. Zebrafish 2017, 14, 35–41. [Google Scholar] [CrossRef]
- Colussi, S.; Campia, V.; Righetti, M.; Scanzio, T.; Riina, M.V.; Burioli, E.A.V.; Foglini, C.; Ingravalle, F.; Prearo, M.; Acutis, P.L. Buccal swab: A tissue sampling method for refinement of experimental procedures involving rainbow trout. J. Appl. Ichthyol. 2017, 33, 515–519. [Google Scholar] [CrossRef]
- LaPatra, S.E.; Rohovec, J.S.; Fryer, J.L. Detection of infectious Hematopoietic Necrosis Virus in fish mucus. Fish. Pathol. 1989, 24, 197–202. [Google Scholar] [CrossRef]
- Drolet, B.S.; Rohovec, J.S.; Leong, J.C. The route of entry and progression of infectious haematopoietic necrosis virus in Oncorhynchus mykiss (Walbaum): A sequential immunohistochemical study. J. Fish. Dis. 1994, 17, 337–344. [Google Scholar] [CrossRef]
- Ford, C.E.; Brookes, L.M.; Skelly, E.; Sergeant, C.; Jordine, T.; Balloux, F.; Nichols, R.A.; Garner, T.W.J. Non-lethal detection of Frog Virus 3-Like (RUK13) and Common Midwife Toad Virus-Like (PDE18) Ranaviruses in two UK-native amphibian species. Viruses 2022, 14, 2635. [Google Scholar] [CrossRef]
- Gui, L.; Li, X.; Lin, S.; Zhao, Y.; Lin, P.; Wang, B.; Tang, R.; Guo, J.; Zu, Y.; Zhou, Y.; et al. Low-cost and rapid method of DNA extraction from scaled fish blood and skin mucus. Viruses 2022, 14, 840. [Google Scholar] [CrossRef]
- Crivelli, A.J. Iucn Iberochondrostoma lusitanicus; The IUCN Red List of Threatened Species; IUCN (International Union for Conservation of Nature): Gland, Switzerland, 2006. [Google Scholar] [CrossRef]
- Coelho, M.M.; Mesquita, N.; Collares-Pereira, M.J. Chondrostoma almacai, a new cyprinid species from the southwest of Portugal, Iberian Peninsula. Folia. Zool. 2005, 54, 201–212. [Google Scholar]
- Crivelli, A.J. Iucn Cobitis paludica; The IUCN Red List of Threatened Species; IUCN (International Union for Conservation of Nature): Gland, Switzerland, 2006. [Google Scholar] [CrossRef]
- Soriguer, M.C.; Vallespín, C.; Gomez-Cama, C.; Hernando, J.A. Age, diet, growth and reproduction of a population of Cobitis paludica (de Buen, 1930) in the Palancar Stream (southwest of Europe, Spain) (Pisces: Cobitidae). Hydrobiologia 2000, 436, 51–58. [Google Scholar] [CrossRef]
- Reid, C.H.; Vandergoot, C.S.; Midwood, J.D.; Stevens, E.D.; Bowker, J.; Cooke, S.J. On the electroimmobilization of fishes for research and practice: Opportunities, challenges, and research needs. Fisheries 2019, 44, 576–585. [Google Scholar] [CrossRef]
- Phillott, A.D.; Speare, R.; Hines, H.B.; Skerratt, L.F.; Meyer, E.; McDonald, K.R.; Cashins, S.D.; Mendez, D.; Berger, L. Minimising exposure of amphibians to pathogens during field studies. Dis. Aquat. Organ. 2010, 92, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Leung, W.T.M.; Thomas-Walters, L.; Garner, T.W.J.; Balloux, F.; Durrant, C.; Price, S.J. A quantitative-PCR based method to estimate ranavirus viral load following normalisation by reference to an ultraconserved vertebrate target. J. Virol. Methods 2017, 249, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Brooks, M.E.; Kristensen, K.; van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Mächler, M.; Bolker, B.M. glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 2017, 9, 378–400. [Google Scholar] [CrossRef]
- Team, R. Rstudio: Integrated Development for r. Rstudio; PBC: Boston, MA, USA, 2020; Available online: http://www.Rstudio.Com (accessed on 9 September 2022).
- Holopainen, R.; Tapiovaara, H.; Honkanen, J. Expression analysis of immune response genes in fish epithelial cells following ranavirus infection. Fish. Shellfish. Immunol. 2012, 32, 1095–1105. [Google Scholar] [CrossRef]
- Becerra, J.; Montes, G.S.; Bexiga, S.R.; Junqueira, L.C. Structure of the tail fin in teleosts. Cell. Tissue. Res 1983, 230, 127–137. [Google Scholar] [CrossRef]
- Saucedo, B.; Garner, T.W.J.; Kruithof, N.; Allain, S.J.R.; Goodman, M.J.; Cranfield, R.J.; Sergeant, C.; Vergara, D.A.; Kik, M.J.L.; Forzán, M.J.; et al. Common midwife toad ranaviruses replicate first in the oral cavity of smooth newts (Lissotriton vulgaris) and show distinct strain-associated pathogenicity. Sci. Rep. 2019, 9, 4453. [Google Scholar] [CrossRef]
- Brunner, J.L.; Schock, D.M.; Collins, J.P. Transmission dynamics of the amphibian ranavirus Ambystoma tigrinum virus. Dis. Aquat. Organ. 2007, 77, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Gray, M.J.; Miller, D.L.; Hoverman, J.T. Ecology and pathology of amphibian ranaviruses. Dis. Aquat. Organ 2009, 87, 243–266. [Google Scholar] [CrossRef] [PubMed]
- Hoverman, J.T.; Gray, M.J.; Miller, D.L. Anuran susceptibilities to ranaviruses: Role of species identity, exposure route, and a novel virus isolate. Dis. Aquat. Organ. 2010, 89, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Robert, J.; George, E.; Andino, F.D.J.; Chen, G. Waterborne infectivity of the Ranavirus Frog Virus 3 in Xenopus laevis. Virology 2011, 417, 410–417. [Google Scholar] [CrossRef]
- Button, K.; Ioannidis, J.; Mokrysz, C.; Nosek, B.A.; Flint, J.; Robinson, E.S.J.; Munafò, M.R. Power failure: Why small sample size undermines the reliability of neuroscience. Nat. Rev. Neurosci. 2013, 14, 365–376. [Google Scholar] [CrossRef]
- Goodman, R.M.; Miller, D.L.; Ararso, Y.T. Prevalence of Ranavirus in Virginia turtles as detected by tail-clip sampling versus oral-cloacal swabbing. Northeast. Nat. 2013, 20, 325–332. [Google Scholar] [CrossRef]
- Das, A.; Spackman, E.; Pantin-Jackwood, M.J.; Suarez, D.L. Removal of real-time reverse transcription polymerase chain reaction (RT-PCR) inhibitors associated with cloacal swab samples and tissues for improved diagnosis of Avian Influenza Virus by RT-PCR. J. Vet. Diagn. Investig. 2009, 21, 771–778. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coutinho, C.D.; Ford, C.E.; Trafford, J.D.; Duarte, A.; Rebelo, R.; Rosa, G.M. Non-Lethal Detection of Ranavirus in Fish. Viruses 2023, 15, 471. https://doi.org/10.3390/v15020471
Coutinho CD, Ford CE, Trafford JD, Duarte A, Rebelo R, Rosa GM. Non-Lethal Detection of Ranavirus in Fish. Viruses. 2023; 15(2):471. https://doi.org/10.3390/v15020471
Chicago/Turabian StyleCoutinho, Catarina D., Charlotte E. Ford, Joseph D. Trafford, Ana Duarte, Rui Rebelo, and Gonçalo M. Rosa. 2023. "Non-Lethal Detection of Ranavirus in Fish" Viruses 15, no. 2: 471. https://doi.org/10.3390/v15020471
APA StyleCoutinho, C. D., Ford, C. E., Trafford, J. D., Duarte, A., Rebelo, R., & Rosa, G. M. (2023). Non-Lethal Detection of Ranavirus in Fish. Viruses, 15(2), 471. https://doi.org/10.3390/v15020471