

Non-Lethal Detection of Frog Virus 3-Like (RUK13) and Common Midwife Toad Virus-Like (PDE18) Ranaviruses in Two UK-Native Amphibian Species
 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Biosecurity
2.2. Collection and Rearing
2.3. Experimental Preparation
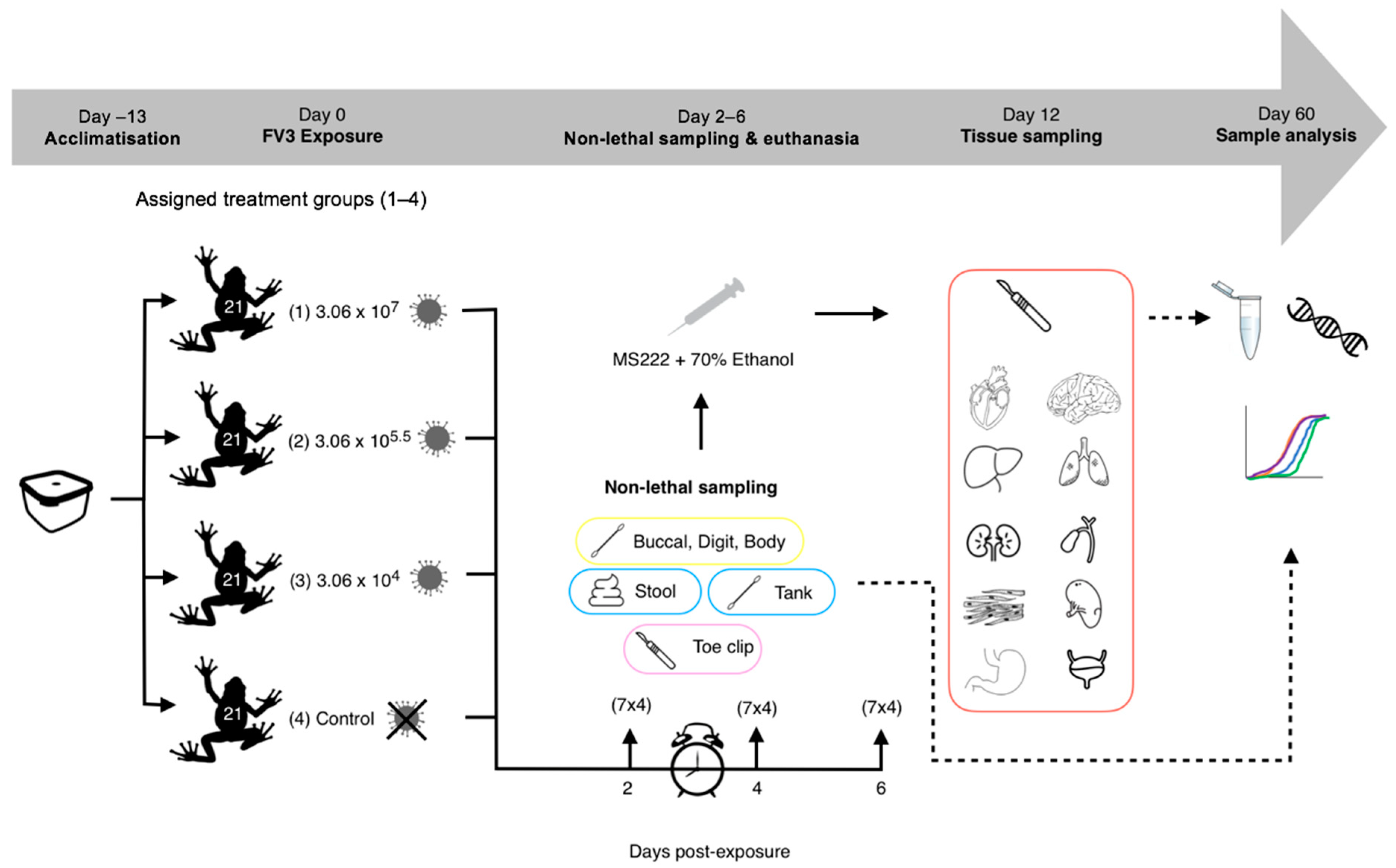
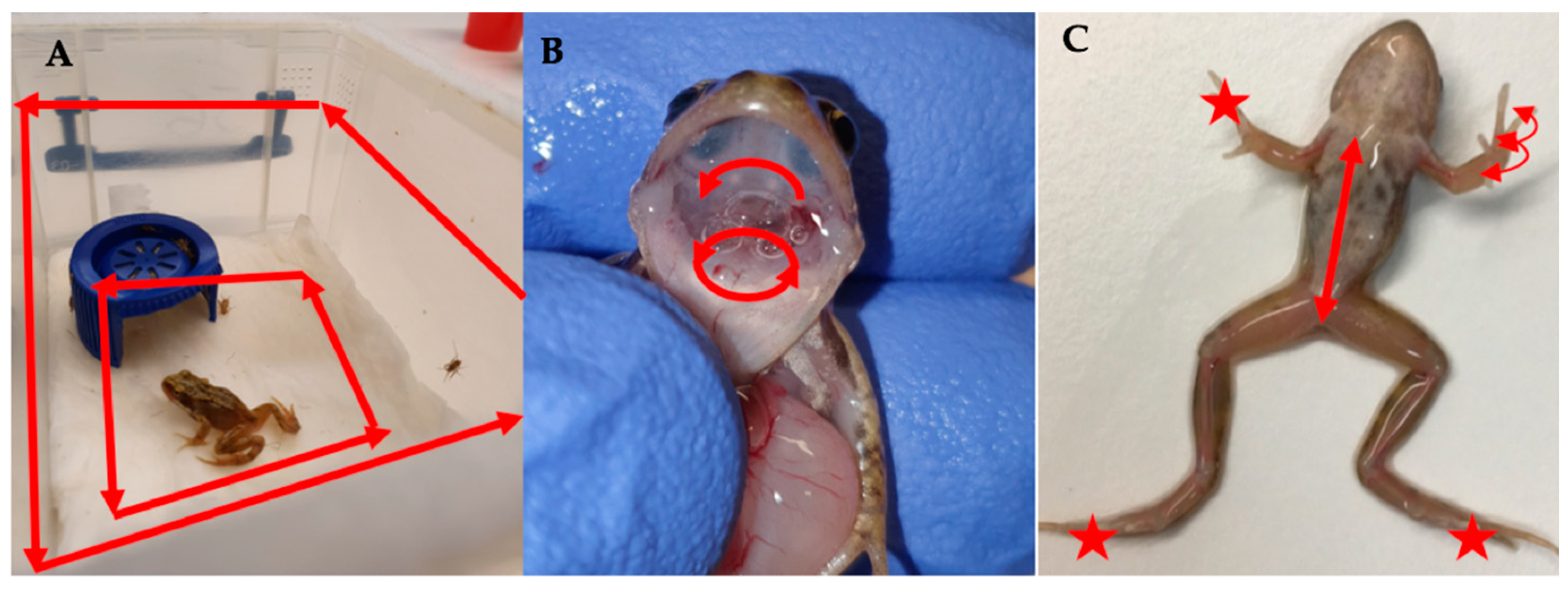
2.4. Experiment 1: Detectability of RUK13 in Post-Metamorphic R. temporaria
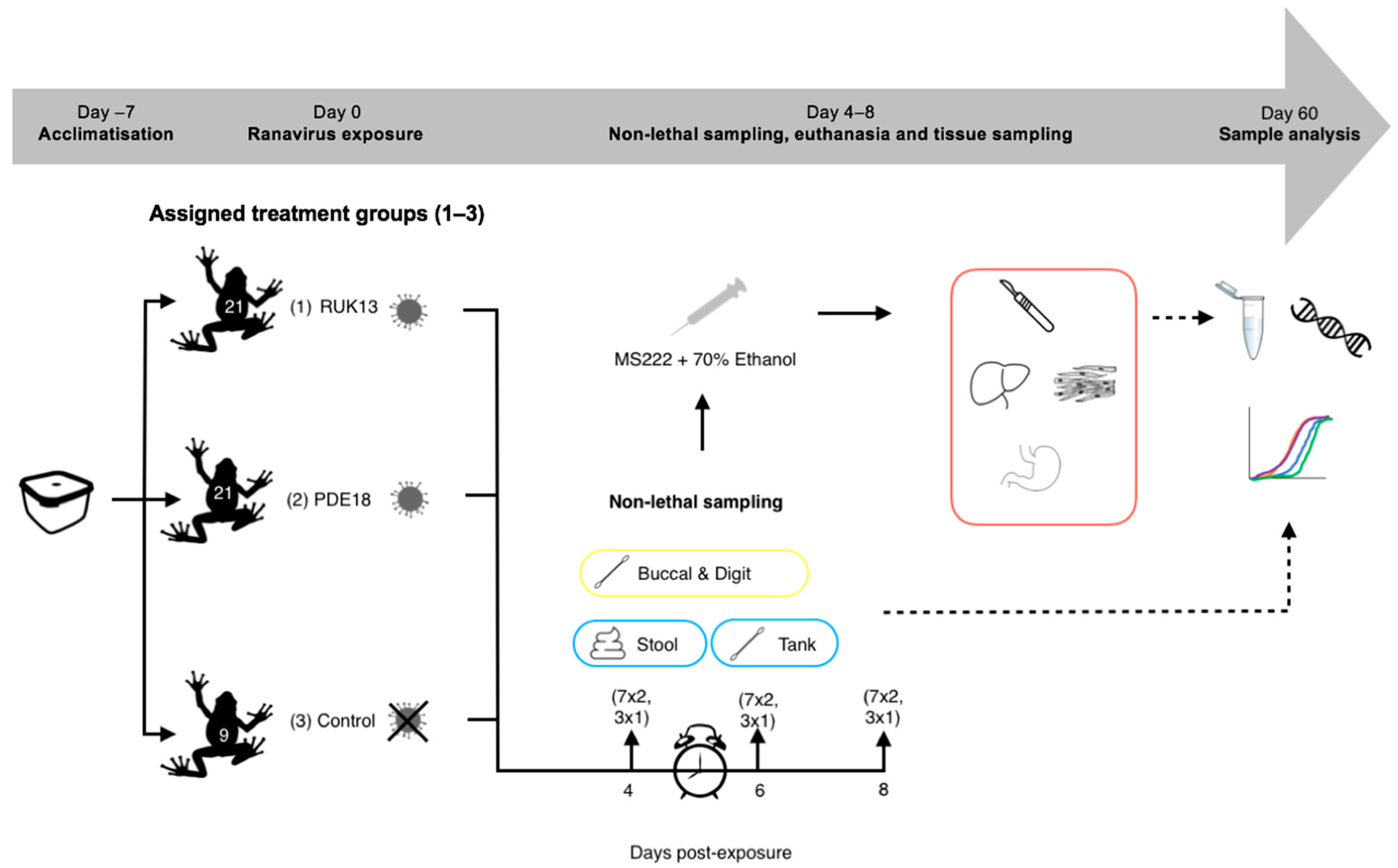
2.5. Experiment 2 (Toads): Detectability of RUK13 and PDE18 in Adult B. bufo
2.6. Extraction and Amplification of Viral DNA
2.7. Analysis
3. Results
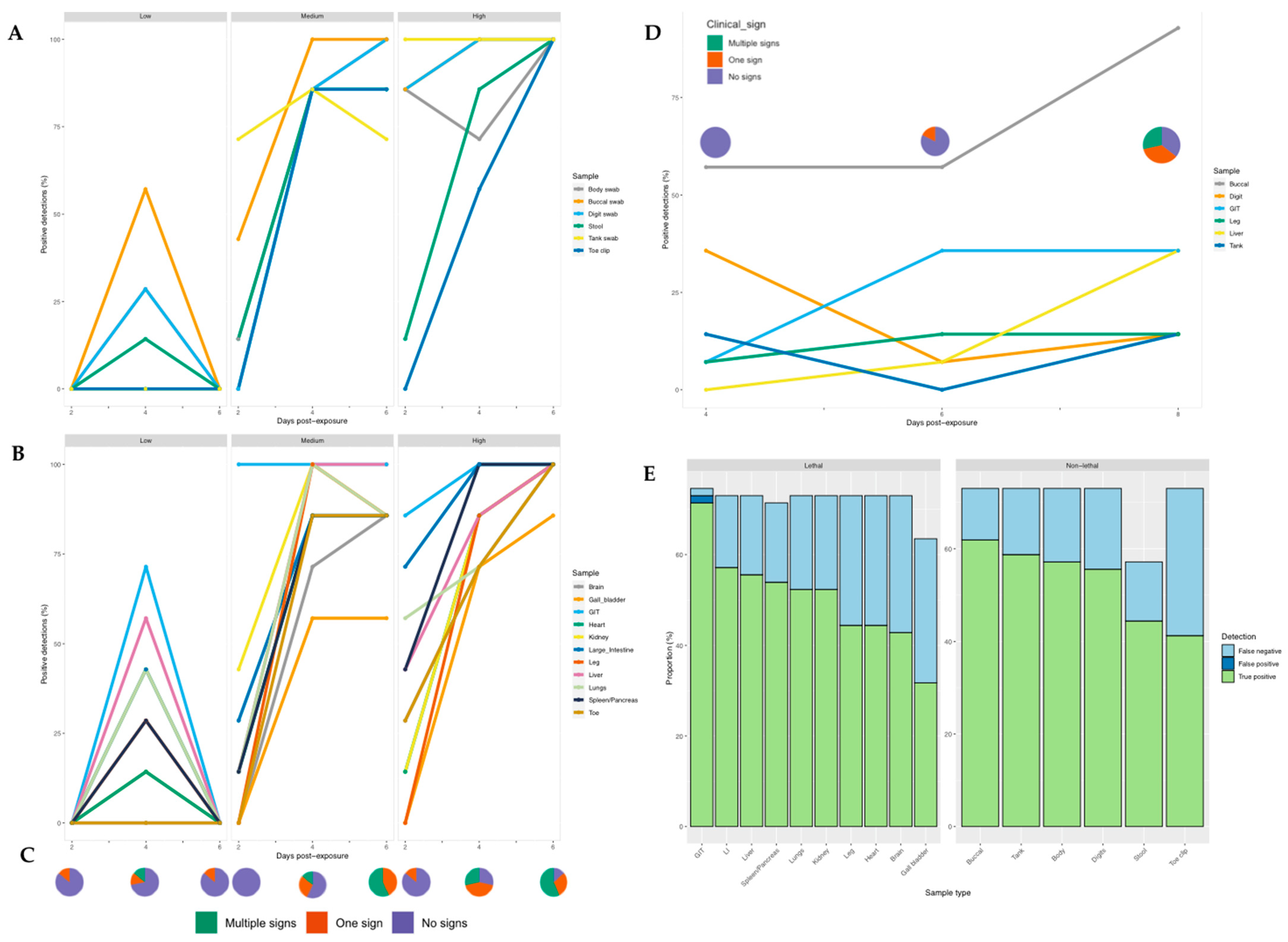
3.1. Infection Dynamics in R. temporaria: Screening for Ranavirus Using Tissue
3.2. Non-Lethal Screening for Ranavirus in R. temporaria
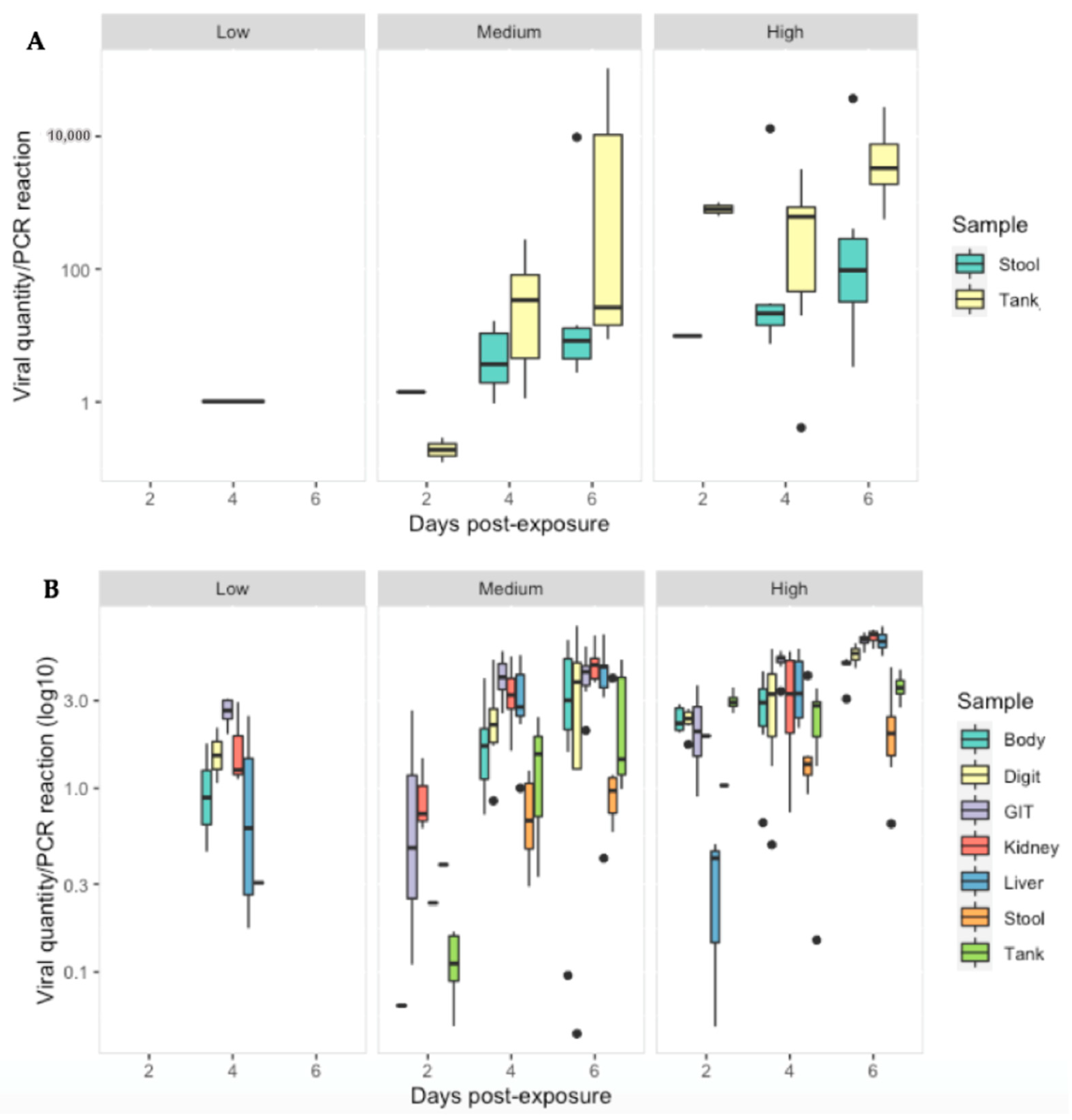
3.3. Ranavirus Shedding in R. temporaria
3.4. Screening for RUK13 and PDE18 in Toads
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Gray, M.J.; Chinchar, V.G. Ranaviruses Lethal Pathogens of Ectothermic Vertebrates; Springer International Publishing: Berlin/Heidelberg, Germany, 2015; Available online: https://link.springer.com/content/pdf/10.1007%2F978-3-319-13755-1.pdf (accessed on 24 January 2019).
- Brunner, J.L.; Schock, D.M.; Davidson, E.W.; Collins, J.P. Intraspecific Reservoirs: Complex Life History and the Persistence of a Lethal Ranavirus. Ecology 2004, 85, 560–566. [Google Scholar] [CrossRef]
- Allender, M.; Mitchell, M.; Torres, T.; Sekowska, J.; Driskell, E. Pathogenicity of Frog Virus 3-like Virus in Red-eared Slider Turtles (Trachemys scripta elegans) at Two Environmental Temperatures. J. Comp. Pathol. 2013, 149, 356–367. [Google Scholar] [CrossRef] [PubMed]
- Miller, D.L.; Pessier, A.P.; Hick, P.; Whittington, R.J. Comparative Pathology of Ranaviruses and Diagnostic Techniques. In Ranaviruses; Springer International Publishing: Cham, Switzerland, 2015; pp. 171–208. [Google Scholar] [CrossRef]
- Garden Wildlife Health. GWH Disease Incident Reports Search. Available online: https://app.bto.org/gwh/results/new/search-main.jsp;jsessionid=68fec2a4eb35d2da48a9dc5ac659 (accessed on 14 October 2022).
- OIE. Manual of Diagnostic Tests for Aquatic Animals: Chapter 2.1.2.—Infection with Ranavirus. Available online: https://www.woah.org/fileadmin/Home/eng/Health_standards/aahm/current/chapitre_ranavirus.pdf (accessed on 23 June 2022).
- Clark, H.F.; Gray, C.; Fabian, F.; Zeigel, R.; Karzon, D.T. Comparative Studies of Amphibian Cytoplasmic Virus Strains Isolated from the Leopard Frog, Bullfrog, and Newt. In Biology of Amphibian Tumors; Springer: Berlin/Heidelberg, Germany, 1969; pp. 310–326. [Google Scholar] [CrossRef]
- Ariel, E.; Holopainen, R.; Olesen, N.J.; Tapiovaara, H. Comparative study of ranavirus isolates from cod (Gadus morhua) and turbot (Psetta maxima) with reference to other ranaviruses. Arch. Virol. 2010, 155, 1261–1271. [Google Scholar] [CrossRef]
- Brunner, J.L.; Olson, A.D.; Rice, J.G.; Meiners, S.E.; Le Sage, M.J.; Cundiff, J.A.; Goldberg, C.S.; Pessier, A.P. Ranavirus infection dynamics and shedding in American bullfrogs: Consequences for spread and detection in trade. Dis. Aquat. Org. 2019, 135, 135–150. [Google Scholar] [CrossRef]
- Blaustein, A.R.; Gervasi, S.S.; Johnson, P.T.J.; Hoverman, J.; Belden, L.; Bradley, P.W.; Xie, G.Y. Ecophysiology meets conservation: Understanding the role of disease in amphibian population declines. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 1688–1707. [Google Scholar] [CrossRef] [PubMed]
- St-Amour, V.; Lesbarrères, D. Genetic evidence of Ranavirus in toe clips: An alternative to lethal sampling methods. Conserv. Genet. 2007, 8, 1247–1250. [Google Scholar] [CrossRef]
- Price, S.J. Emergence of a Virulent Wildlife Disease: Using Spatial Epidemiology and Phylogenetic Methods to Reconstruct the Spread of Amphibian Viruses. Ph.D. Thesis, Queen Mary University of London, London, UK, 2013. [Google Scholar]
- Saucedo, B.; Garner, T.W.J.; Kruithof, N.; Allain, S.J.R.; Goodman, M.J.; Cranfield, R.J.; Sergeant, C.; Vergara, D.A.; Kik, M.J.L.; Forzán, M.J.; et al. Common midwife toad ranaviruses replicate first in the oral cavity of smooth newts (Lissotriton vulgaris) and show distinct strain-associated pathogenicity. Sci. Rep. 2019, 9, 4453. [Google Scholar] [CrossRef]
- Goodman, R.M.; Miller, D.; Ararso, Y.T. Prevalence of Ranavirus in Virginia Turtles as Detected by Tail-Clip Sampling Versus Oral-Cloacal Swabbing. Northeast. Nat. 2013, 20, 325–332. Available online: file:///Users/charlotteford/Downloads/045.020.0208.pdf (accessed on 29 March 2022). [CrossRef]
- Brunner, J.L.; Olson, D.H.; Gray, M.J.; Miller, D.L.; Duffus, A.L. Global patterns of ranavirus detections. FACETS 2021, 6, 912–924. [Google Scholar] [CrossRef]
- Bucko, M.; Gieger, S. Ranaviruses: OIE Technical Disease Card. Available online: https://www.oie.int/app/uploads/2021/03/ranavirusesinfection-with.pdf (accessed on 23 March 2022).
- Lesbarrères, D.; Balseiro, A.; Brunner, J.; Chinchar, V.G.; Duffus, A.; Kerby, J.; Miller, D.; Robert, J.; Schock, D.M.; Waltzek, T.; et al. Ranavirus: Past, present and future. Biol. Lett. 2012, 8, 481–483. [Google Scholar] [CrossRef] [PubMed]
- Forzán, M.J.; Jones, K.M.; Ariel, E.; Whittington, R.J.; Wood, J.; Markham, R.J.F.; Daoust, P.-Y. Pathogenesis of Frog Virus 3 (Ranavirus, Iridoviridae) Infection in Wood Frogs (Rana sylvatica). SAGE 2017, 54, 531–548. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, A.A.; Langton, T.E.S.; Bennett, P.M.; Lewin, J.F.; Drury, S.E.N.; Gough, R.E.; Macgregor, S.K. Pathological and microbiological findings from incidents of unusual mortality of the common frog (Rana temporaria). Philos. Trans. R. Soc. B Biol. Sci. 1996, 351, 1539–1557. Available online: https://royalsocietypublishing.org/doi/pdf/10.1098/rstb.1996.0140 (accessed on 16 May 2019). [CrossRef]
- Cunningham, A.; Tems, C.; Russell, P. Immunohistochemical Demonstration of Ranavirus Antigen in the Tissues of Infected Frogs (Rana temporaria) with Systemic Haemorrhagic or Cutaneous Ulcerative Disease. J. Comp. Pathol. 2008, 138, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Rijks, J.M.; Saucedo, B.; Der Sluijs, A.S.-V.; Wilkie, G.S.; Van Asten, A.J.A.M.; Broek, J.V.D.; Boonyarittichaikij, R.; Stege, M.; Van Der Sterren, F.; Martel, A.; et al. Investigation of Amphibian Mortality Events in Wildlife Reveals an On-Going Ranavirus Epidemic in the North of the Netherlands. PLoS ONE 2016, 11, e157473. [Google Scholar] [CrossRef] [PubMed]
- Price, S.J.; Ariel, E.; Maclaine, A.; Rosa, G.M.; Gray, M.J.; Brunner, J.L.; Garner, T.W. From fish to frogs and beyond: Impact and host range of emergent ranaviruses. Virology 2017, 511, 272–279. [Google Scholar] [CrossRef]
- Rosa, G.M.; Pinto, J.S.; Laurentino, T.G.; Martel, A.; Pasmans, F.; Rebelo, R.; Griffiths, R.A.; Stöhr, A.C.; Marschang, R.E.; Price, S.; et al. Impact of asynchronous emergence of two lethal pathogens on amphibian assemblages. Sci. Rep. 2017, 7, srep43260. [Google Scholar] [CrossRef]
- Teacher, A.G.F.; Cunningham, A.A.; Garner, T.W.J. Assessing the long-term impact of Ranavirus infection in wild common frog populations. Anim. Conserv. 2010, 13, 514–522. [Google Scholar] [CrossRef]
- Price, S.J.; Wadia, A.; Wright, O.; Leung, W.T.M.; Cunningham, A.A.; Lawson, B. Screening of a long-term sample set reveals two Ranavirus lineages in British herpetofauna. PLoS ONE 2017, 12, e0184768. [Google Scholar] [CrossRef] [PubMed]
- Bayley, A.; Hill, B.; Feist, S. Susceptibility of the European common frog Rana temporaria to a panel of ranavirus isolates from fish and amphibian hosts. Dis. Aquat. Org. 2013, 103, 171–183. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, A.A.; Hyatt, A.D.; Russell, P.; Bennett, P.M. Emerging epidemic diseases of frogs in Britain are dependent on the source of ranavirus agent and the route of exposure. Epidemiol. Infect. 2007, 135, 1200–1212. [Google Scholar] [CrossRef] [PubMed]
- Pearman, P.B.; Garner, T.W.J.; Straub, M.; Greber, U.F. Response of the Italian Agile Frog (Rana Latastei) to a Ranavirus, Frog Virus 3: A Model for Viral Emergence in Naïve Populations. J. Wildl. Dis. 2004, 40, 660–669. Available online: https://www.jwildlifedis.org/doi/pdf/10.7589/0090-3558-40.4.660 (accessed on 7 June 2019). [CrossRef] [PubMed]
- Duffus, A.L.J.; Nichols, R.A.; Garner, T.W.J. Experimental evidence in support of single host maintenance of a multihost pathogen. Ecosphere 2014, 5, 1–11. [Google Scholar] [CrossRef]
- Gray, M.J.; Miller, D.; Hoverman, J. Reliability of non-lethal surveillance methods for detecting ranavirus infection. Dis. Aquat. Org. 2012, 99, 1–6. [Google Scholar] [CrossRef]
- Robert, J.; Abramowitz, L.; Gantress, J.; Morales, H.D. Xenopus Laevis: A Possible Vector of Ranavirus Infection? J. Wildl. Dis. 2007, 43, 645–652. [Google Scholar] [CrossRef] [PubMed]
- Leung, W.T.; Thomas-Walters, L.; Garner, T.W.; Balloux, F.; Durrant, C.; Price, S.J. A quantitative-PCR based method to estimate ranavirus viral load following normalisation by reference to an ultraconserved vertebrate target. J. Virol. Methods 2017, 249, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Wynne, F.; Puschendorf, R.; Knight, M.; Price, S. Choice of molecular assay determines ranavirus detection probability and inferences about prevalence and occurrence. Dis. Aquat. Org. 2020, 141, 139–147. [Google Scholar] [CrossRef]
- Morales, H.D.; Abramowitz, L.; Gertz, J.; Sowa, J.; Vogel, A.; Robert, J. Innate Immune Responses and Permissiveness to Ranavirus Infection of Peritoneal Leukocytes in the Frog Xenopus laevis. J. Virol. 2010, 84, 4912–4922. [Google Scholar] [CrossRef] [PubMed]
- Broquet, T.; Berset-Braendli, L.; Emaresi, G.; Fumagalli, L. Buccal swabs allow efficient and reliable microsatellite genotyping in amphibians. Conserv. Genet. 2007, 8, 509–511. [Google Scholar] [CrossRef]
- Robert, J.; George, E.; Andino, F.D.J.; Chen, G. Waterborne infectivity of the Ranavirus frog virus 3 in Xenopus laevis. Virology 2011, 417, 410–417. [Google Scholar] [CrossRef] [PubMed]
- Hoverman, J.; Gray, M.J.; Miller, D. Anuran susceptibilities to ranaviruses: Role of species identity, exposure route, and a novel virus isolate. Dis. Aquat. Org. 2010, 89, 97–107. [Google Scholar] [CrossRef]
- Duffus, A.L.; Garner, T.W.; Nichols, R.A.; Standridge, J.P.; Earl, J.E. Modelling Ranavirus Transmission in Populations of Common Frogs (Rana temporaria) in the United Kingdom. Viruses 2019, 11, 556. [Google Scholar] [CrossRef]
- Duffus, A.L.J.; Nichols, R.A.; Garner, T.W.J. Detection of a Frog Virus 3-like Ranavirus in Native and Introduced Amphibians in the United Kingdom in 2007 and 2008. Herptological Rev. 2014, 45, 608–610. Available online: http://jncc.defra.gov.uk/page-5166- (accessed on 20 May 2019).
- North, A.; Hodgson, D.J.; Price, S.J.; Griffiths, A.G.F. Anthropogenic and Ecological Drivers of Amphibian Disease (Ranavirosis). PLoS ONE 2015, 10, e0127037. [Google Scholar] [CrossRef]
- Price, S.J.; Garner, T.W.; Nichols, R.A.; Balloux, F.; Ayres, C.; de Alba, A.M.-C.; Bosch, J. Collapse of Amphibian Communities Due to an Introduced Ranavirus. Curr. Biol. 2014, 24, 2586–2591. [Google Scholar] [CrossRef] [PubMed]
- Brookes, L.B. Reduction and Refinement of Non-Model Amphibians Used in Amphibian Disease Testing. Ph.D. Thesis, The Royal Veterinary College, Herts, UK, 2022. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Individuals Sampled | Procedure | Storage Prior to Extraction | |||||
|---|---|---|---|---|---|---|---|---|
| Sample type | Lethal | Destructive | Invasive | Day 2 | Day 4 | Day 6 | ||
| Pre-death swab (control only) | x | 7 | 7 | 7 | Pre-euthanasia x 5 over stomach and each digit | 4 °C | ||
| Buccal swab | x | (H) 7 (M) 7 (L) 7 (C) 6 | (H) 7 (M) 7 (L) 7 (C) 6 | (H) 7 (M) 7 (L) 7 (C) 6 | Post-euthanasia x 5 inside the mouth and around the tongue | 4 °C | ||
| Digit swab | x | (H) 7 (M) 7 (L) 7 | (H) 7 (M) 7 (L) 7 | (H) 7 (M) 7 (L) 7 | Pre-euthanasia x 5 over each digit | 4 °C | ||
| Body swab | x | (H) 7 (M) 7 (L) 7 | (H) 7 (M) 7 (L) 7 | (H) 7 (M) 7 (L) 7 | Pre-euthanasia x 5 over the stomach | 4 °C | ||
| Environmental (tank) swab | (H) 7 (M) 7 (L) 7 | (H) 7 (M) 7 (L) 7 | (H) 7 (M) 7 (L) 7 | Pre-euthanasia x 5 over the walls and floor of the housing | 4 °C | |||
| Stool | (H) 7 (M) 7 (L) 7 | (H) 7 (M) 7 (L) 7 | (H) 7 (M) 7 (L) 7 | Pre-euthanasia Collected using forceps | −20 °C | |||
| Toe clip | x | x | (H) 7 (M) 7 (L) 7 | (H) 7 (M) 7 (L) 7 | (H) 7 (M) 7 (L) 7 | Post-euthanasia Remove fourth right toe of the right foot | 70% Ethanol | |
| Tissue (e.g., liver) | x | (H) 7 (M) 7 (L) 7 | (H) 7 (M) 7 (L) 7 | (H) 7 (M) 7 (L) 7 | Post-euthanasia Liver, Gastro-intestinal tract, Leg & (13) Tongue | 70% Ethanol | ||
| Category | Individuals Sampled | Procedure | Storage Prior to Extraction | |||||
|---|---|---|---|---|---|---|---|---|
| Sample type | Lethal | Destructive | Invasive | Day 4 | Day 6 | Day 8 | ||
| Buccal swab | x | (R) 7 (P) 7 (C) 3 | (R) 7 (P) 7 (C) 3 | (R) 7 (P) 7 (C) 3 | Post-euthanasia x 5 inside the mouth and around the tongue | 4 °C | ||
| Digit swab | x | (R) 7 (P) 7 (C) 3 | (R) 7 (P) 7 (C) 3 | (R) 7 (P) 7 (C) 3 | Pre-euthanasia x 5 over each digit | 4 °C | ||
| Tank swab | (R) 7 (P) 7 (C) 3 | ((R) 7 (P) 7 (C) 3 | (R) 7 (P) 7 (C) 3 | Pre-euthanasia x 5 over the walls and floor of the housing | 4 °C | |||
| Stool | (R) 7 (P) 7 (C) 3 | (R) 7 (P) 7 (C) 3 | (R) 7 (P) 7 (C) 3 | Pre-euthanasia Collected using forceps | −20 °C | |||
| Tissue (e.g., liver) | x | (R) 7 (P) 7 (C) 3 | (R) 7 (P) 7 (C) 3 | (R) 7 (P) 7 (C) 3 | Post-euthanasia Liver, Gastro-intestinal tract, Leg & (13) Tongue | 70% Ethanol | ||
| Assay (Ranked by Rate) | Acronym | Detection Rate % | Significant Differences | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| GIT | BS | TS | BdS | LI | DS | LV | S/P | LU | KD | LG | HT | ST | BR | TC | GB | |||
| Gastro-intestinal tract | GIT | 71 | NS | NS | NS | NS | NS | NS | * | ** | ** | *** | *** | * | *** | *** | *** | |
| Buccal swab | BS | 62 | NS | NS | NS | NS | NS | NS | NS | NS | NS | NS | NS | * | ** | *** | ||
| Tank swab | TS | 59 | NS | NS | NS | NS | NS | NS | NS | NS | NS | NS | NS | NS | ** | |||
| Body swab | BdS | 57 | NS | NS | NS | NS | NS | NS | NS | NS | NS | NS | NS | * | ||||
| Large intestine | LI | 57 | NS | NS | NS | NS | NS | NS | NS | NS | NS | NS | * | |||||
| Digit swab | DS | 56 | NS | NS | NS | NS | NS | NS | NS | NS | NS | * | ||||||
| Liver | TV | 56 | NS | NS | NS | NS | NS | NS | NS | NS | * | |||||||
| Spleen/pancreas | S/P | 54 | NS | NS | NS | NS | NS | NS | NS | NS | ||||||||
| Lungs | LU | 52 | NS | NS | NS | NS | NS | NS | NS | |||||||||
| Kidney | KD | 52 | NS | NS | NS | NS | NS | NS | ||||||||||
| Leg | LG | 44 | NS | NS | NS | NS | NS | |||||||||||
| Heart | HT | 44 | NS | NS | NS | NS | ||||||||||||
| Stool | ST | 44 | NS | NS | NS | |||||||||||||
| Brain | BR | 43 | NS | NS | ||||||||||||||
| Toe clip | TC | 41 | NS | |||||||||||||||
| Gall bladder | GB | 32 | ||||||||||||||||
| Assay (Ranked by Rate) | Detection Rate % | Significant Differences | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| RUK13 | PDE18 | Total | GIT | DS | LV | LG | TS | GIT | DS | LV | LG | TS | GIT | DS | LV | LG | TS | |
| Buccal swab (BS) | 86 | 57 | 71 | *** | *** | *** | *** | *** | ** | * | ** | ** | ** | NS | NS | NS | NS | NS |
| Gastro-intestinal tract (GIT) | 28 | 33 | 26 | NS | NS | NS | NS | NS | NS | NS | NS | NS | NS | NS | NS | |||
| Digit swab (DS) | 24 | 1 | 19 | NS | NS | NS | NS | NS | NS | NS | NS | NS | ||||||
| Liver (LV) | 1 | 19 | 14 | Total | NS | NS | RUK13 | NS | NS | PDE18 | NS | NS | ||||||
| Leg (LG) | 14 | 1 | 12 | NS | NS | NS | ||||||||||||
| Tank swab (TS) | 1 | 1 | 10 | |||||||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ford, C.E.; Brookes, L.M.; Skelly, E.; Sergeant, C.; Jordine, T.; Balloux, F.; Nichols, R.A.; Garner, T.W.J. Non-Lethal Detection of Frog Virus 3-Like (RUK13) and Common Midwife Toad Virus-Like (PDE18) Ranaviruses in Two UK-Native Amphibian Species. Viruses 2022, 14, 2635. https://doi.org/10.3390/v14122635
Ford CE, Brookes LM, Skelly E, Sergeant C, Jordine T, Balloux F, Nichols RA, Garner TWJ. Non-Lethal Detection of Frog Virus 3-Like (RUK13) and Common Midwife Toad Virus-Like (PDE18) Ranaviruses in Two UK-Native Amphibian Species. Viruses. 2022; 14(12):2635. https://doi.org/10.3390/v14122635
Chicago/Turabian StyleFord, Charlotte E., Lola M. Brookes, Emily Skelly, Chris Sergeant, Tresai Jordine, Francois Balloux, Richard A. Nichols, and Trenton W. J. Garner. 2022. "Non-Lethal Detection of Frog Virus 3-Like (RUK13) and Common Midwife Toad Virus-Like (PDE18) Ranaviruses in Two UK-Native Amphibian Species" Viruses 14, no. 12: 2635. https://doi.org/10.3390/v14122635
APA StyleFord, C. E., Brookes, L. M., Skelly, E., Sergeant, C., Jordine, T., Balloux, F., Nichols, R. A., & Garner, T. W. J. (2022). Non-Lethal Detection of Frog Virus 3-Like (RUK13) and Common Midwife Toad Virus-Like (PDE18) Ranaviruses in Two UK-Native Amphibian Species. Viruses, 14(12), 2635. https://doi.org/10.3390/v14122635