
Arboviruses in Mammals in the Neotropics: A Systematic Review to Strengthen Epidemiological Monitoring Strategies and Conservation Medicine

 ,
,  and
and 
Abstract
1. Introduction
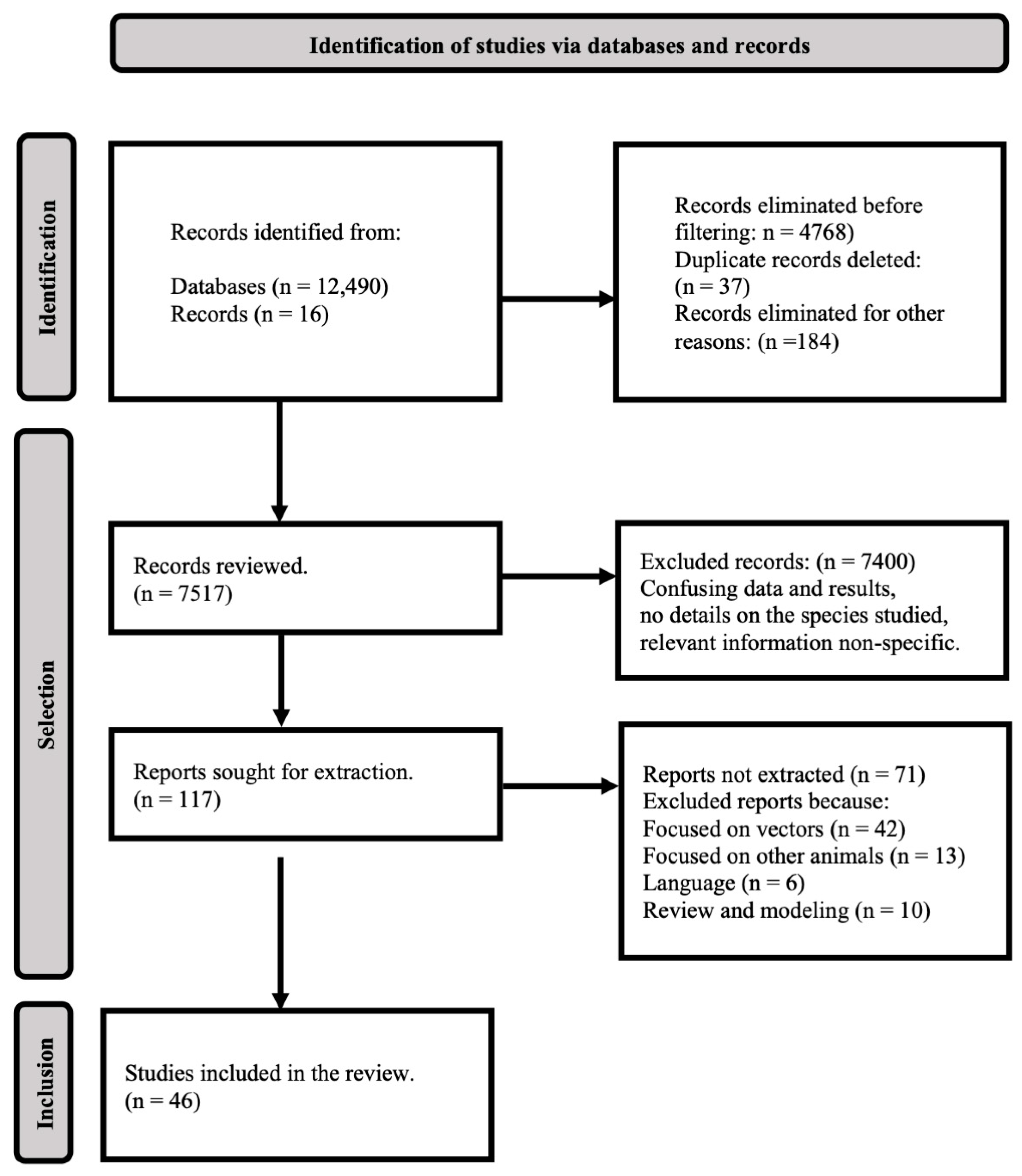
2. Methods
3. Results
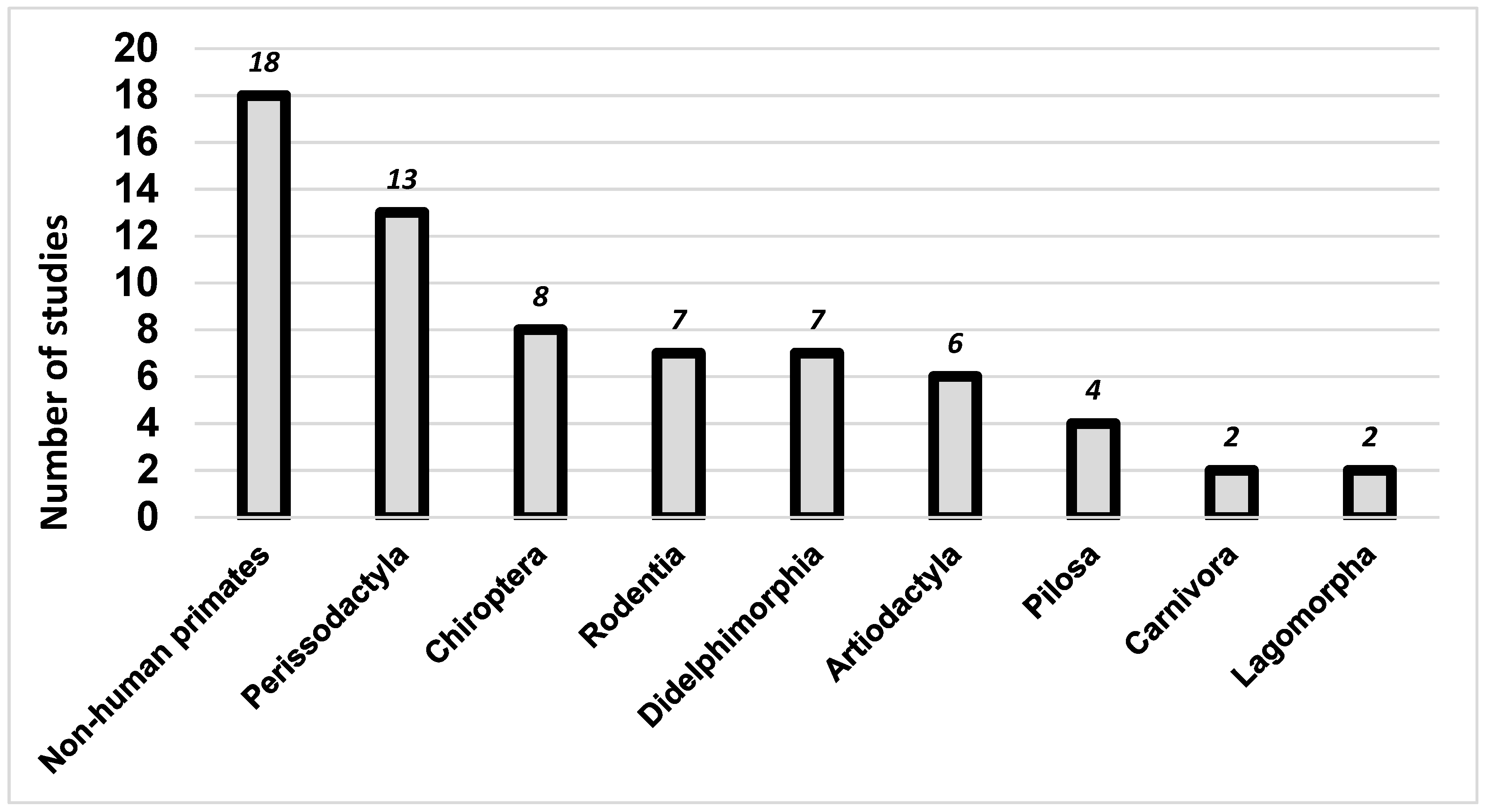
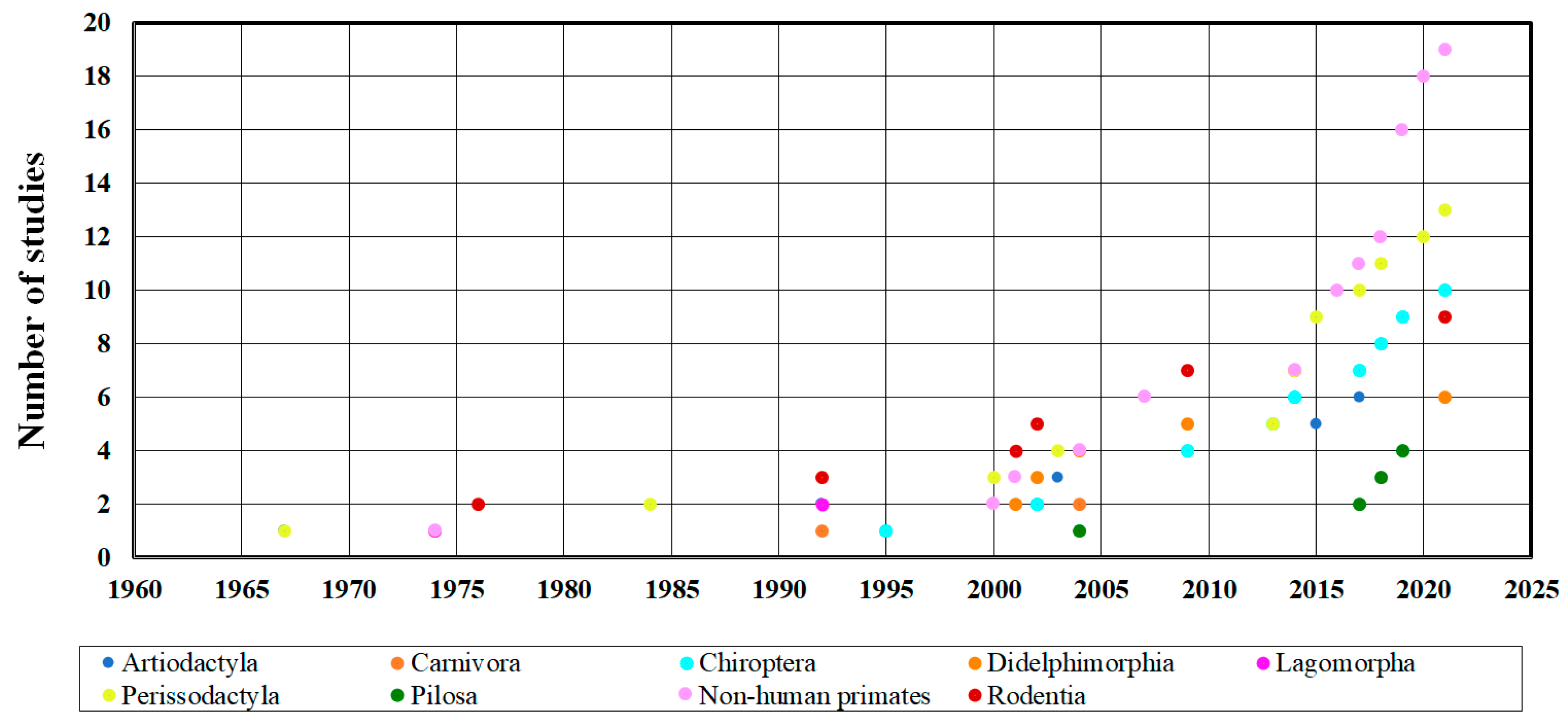
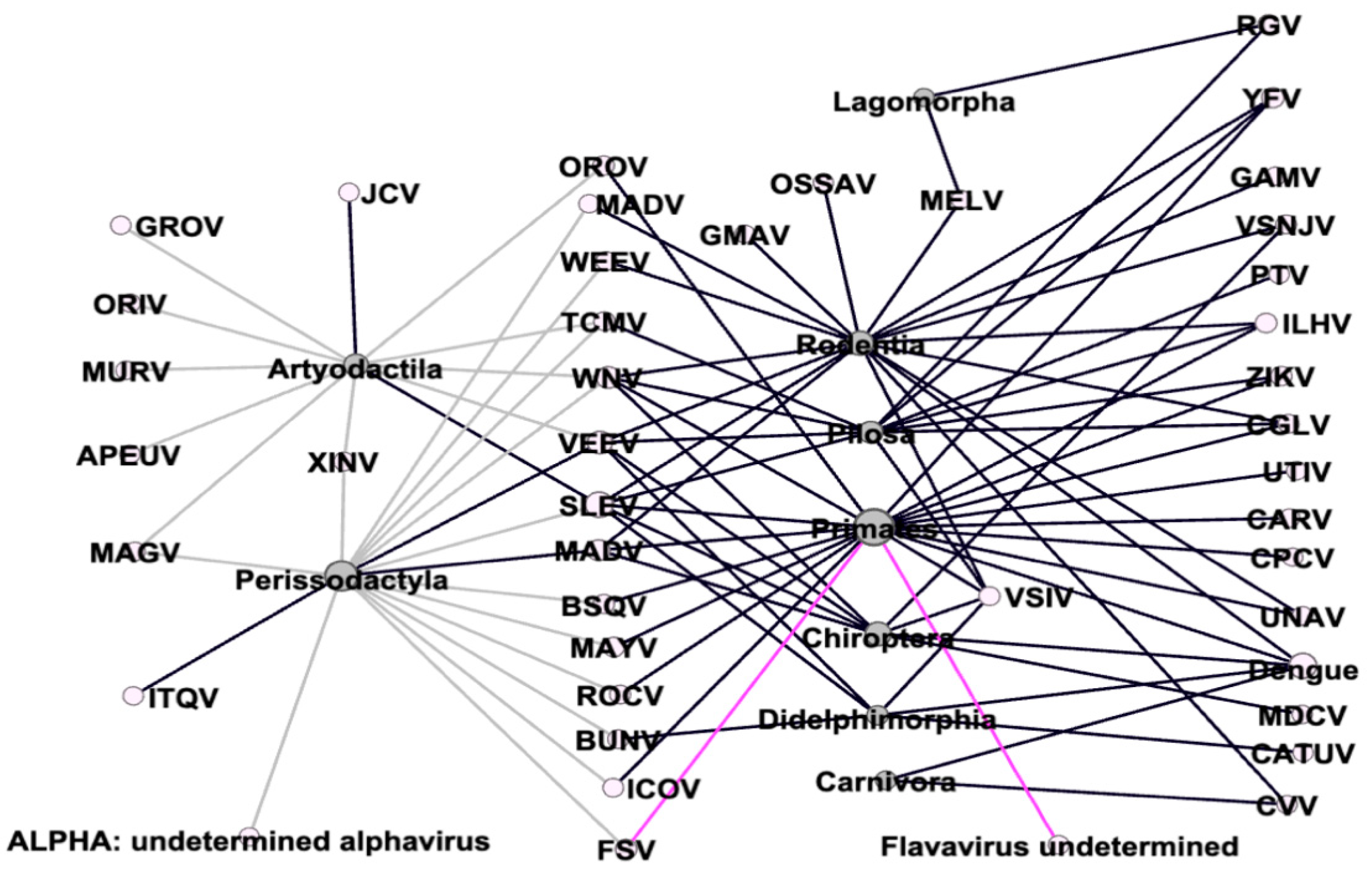
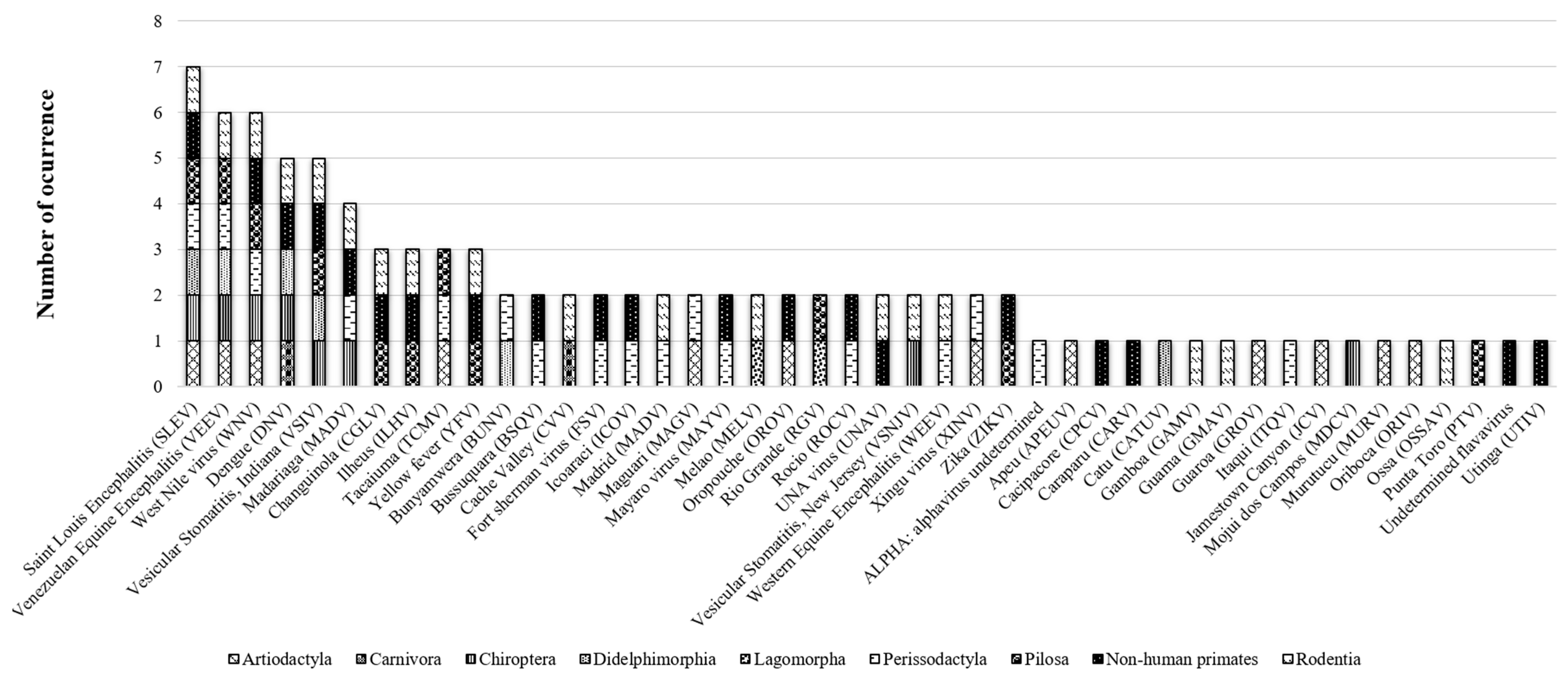
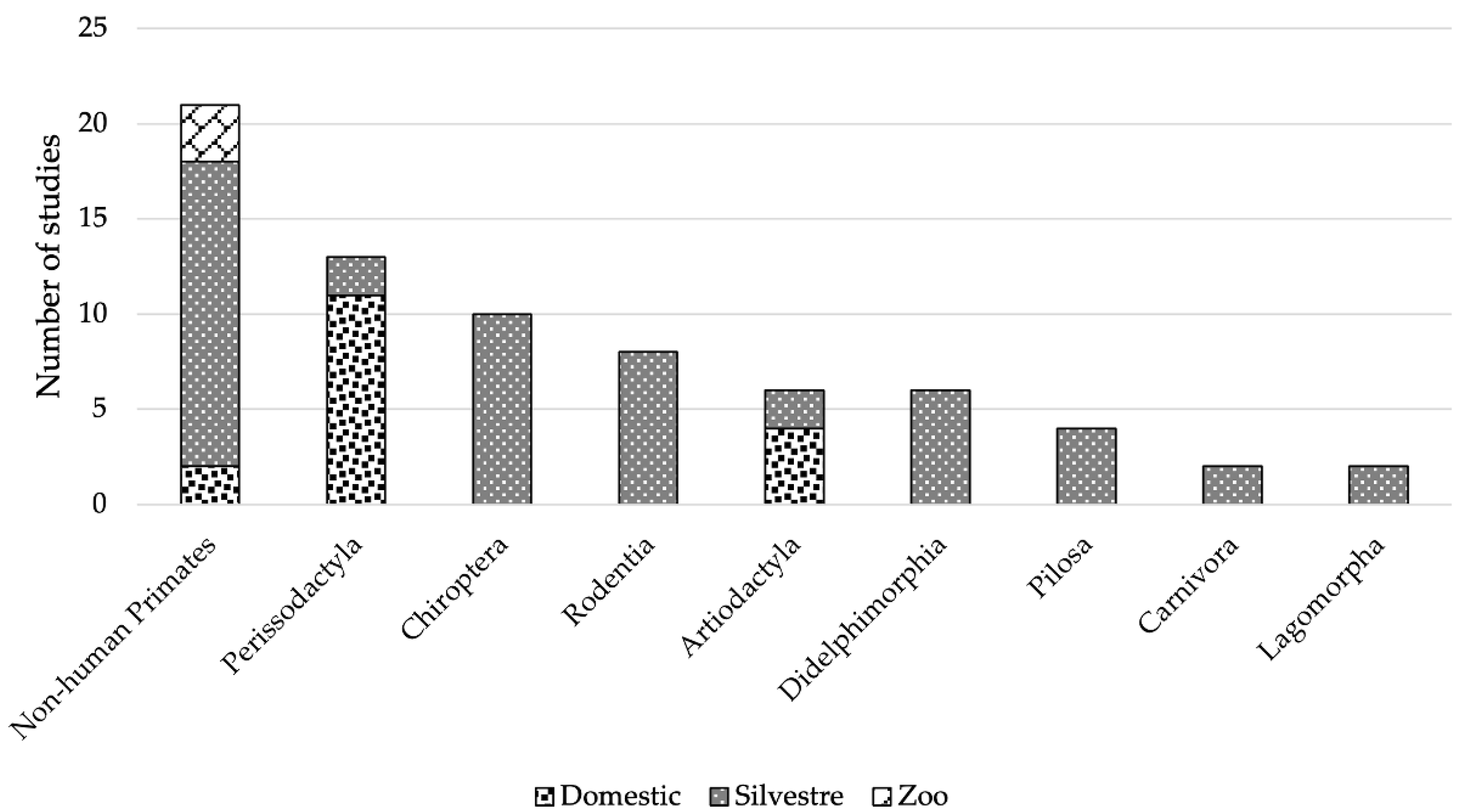
3.1. Arboviruses Richness
3.2. Prevalence and Detection Methodology
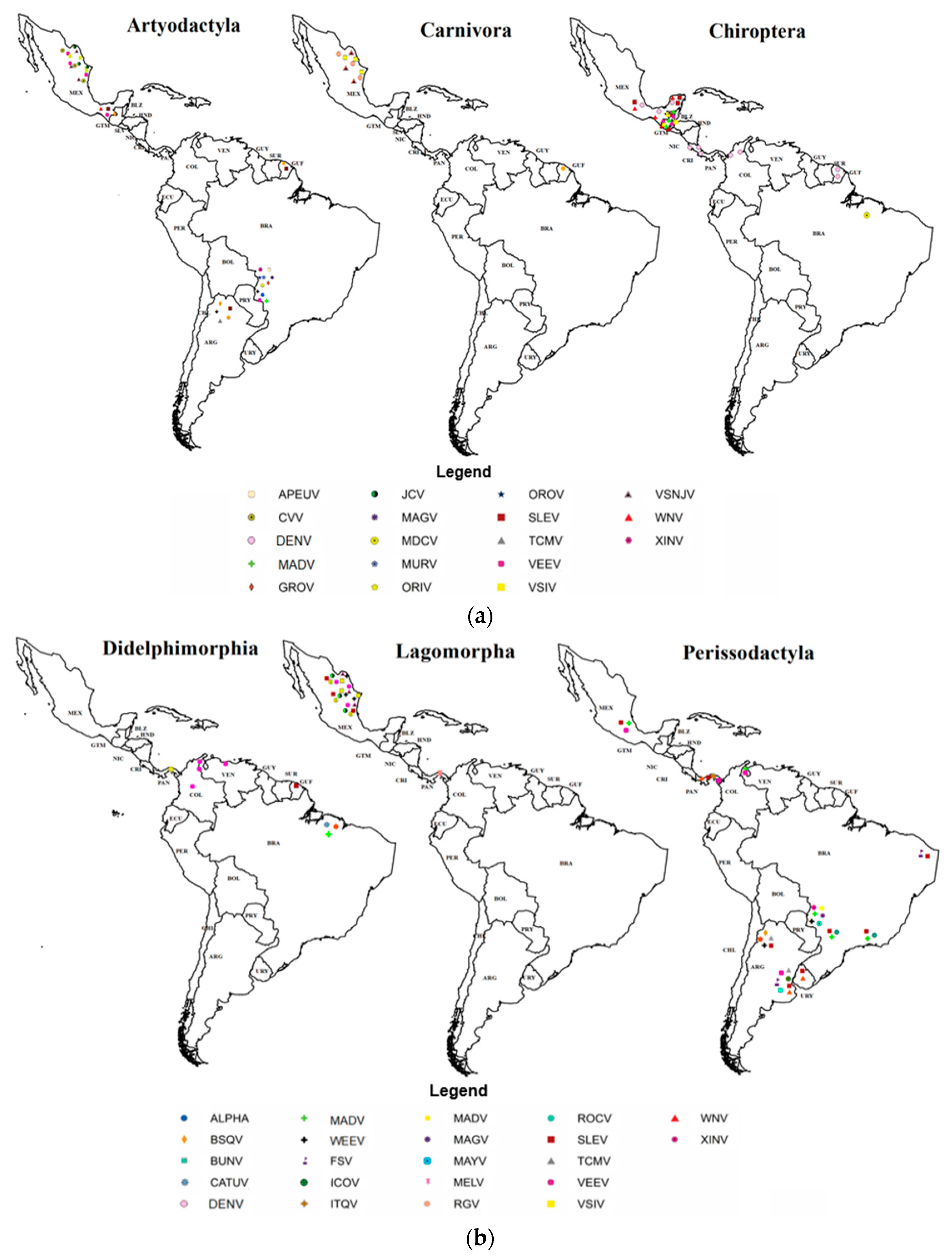
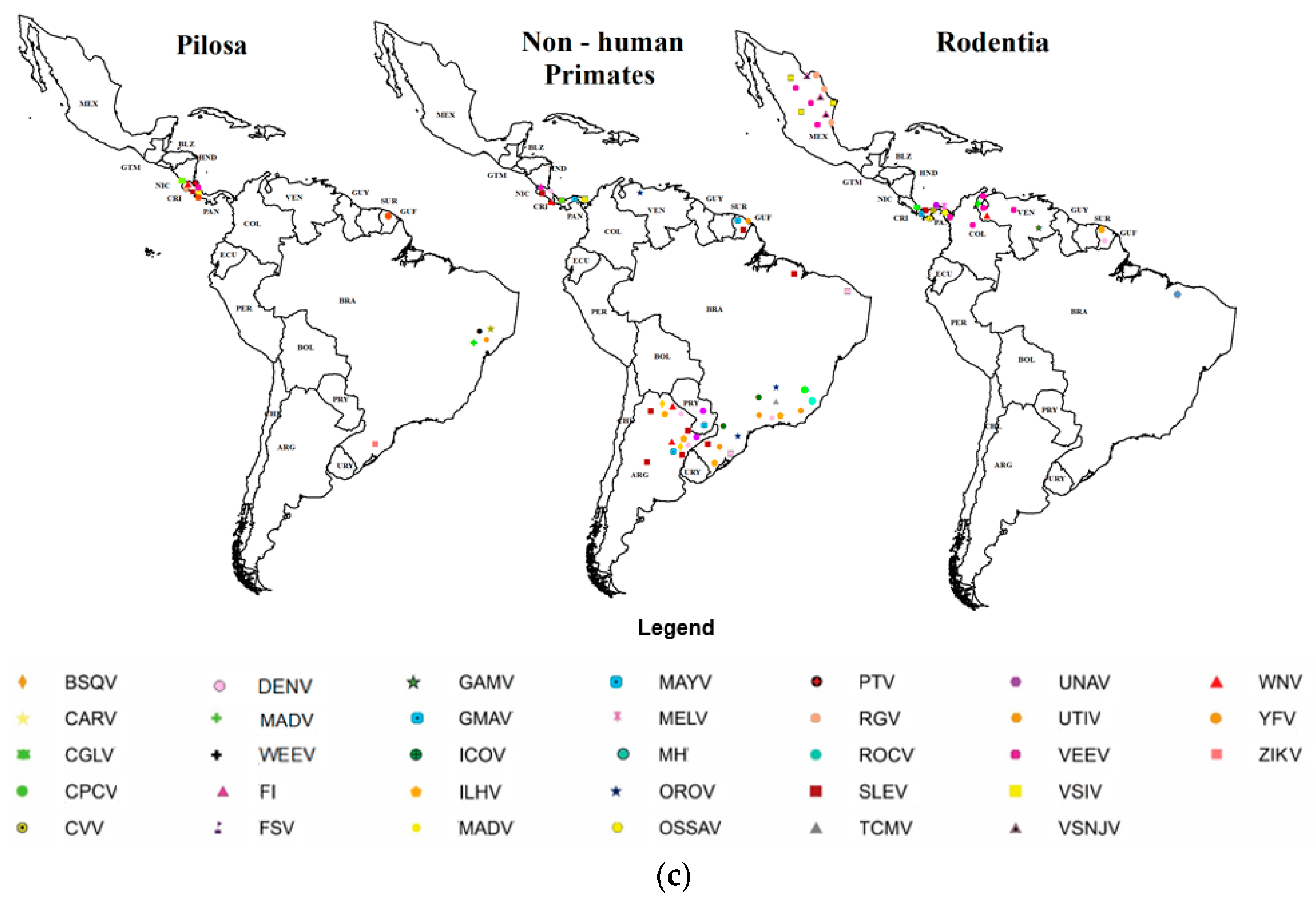
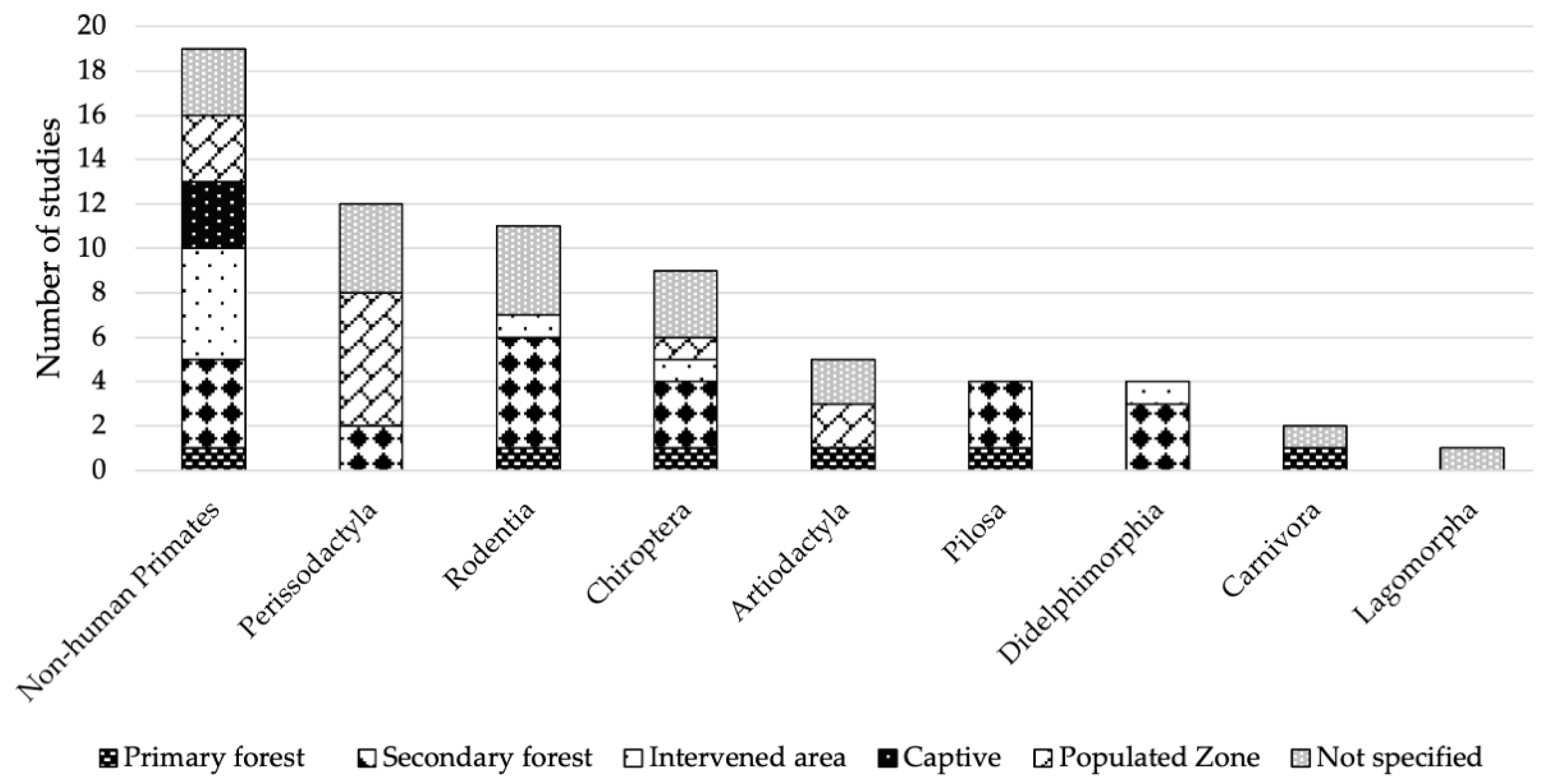
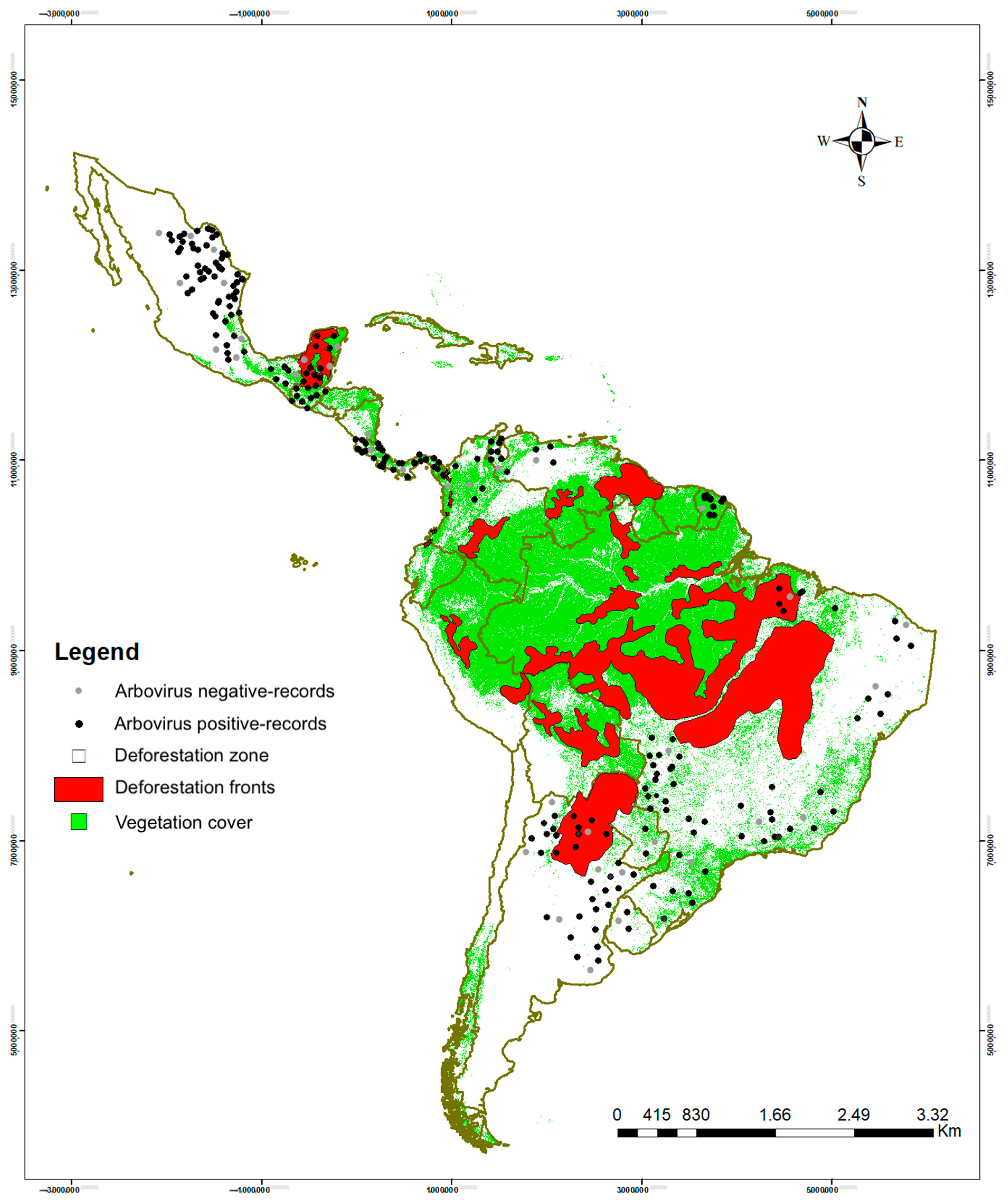
3.3. Habitat Types with Positive Orders and Sampling Conditions According to Order
3.4. Risk Factors
4. Discussion
4.1. Richness of Arboviruses
4.2. Prevalence and Detection Methodologies
4.3. Risk Factors
4.4. Socio-Ecological Aspects
4.5. Ecological Aspects
4.6. Health Policies
4.7. Actions to Take
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mammalian Host | Environment | Country | Arbovirus | Habitat | References |
|---|---|---|---|---|---|
| Artiodactyla | Domestic | Argentina Brazil Mexico | Apeu virus (APEUV) Guaroa virus (GROV) Venezuelan equine encephalitis virus (VEEV) Maguari virus (MAGV) Murutucu virus (MURV) Oriboca virus (ORIV) Oropouche virus (OROV) Tacaiuma virus (TCMV) West Nile virus (WNV) Xingu virus (XINV) | Primary forest Populated zone Not specified | [98,99,104,129] |
| Wild | French Guiana Mexico | Saint Louis encephalitis virus (SLEV) Jamestown Canyon virus (JCV) | Primary fores Not specified | [105,114] | |
| Carnívora | Wild | French Guiana Mexico | Dengue virus (DENV) Cache Valley virus (CVV) | Primary forest Not specified | [105,114] |
| Chiroptera | Wild | Brazil Colombia Costa Rica Guatemala French Guiana Mexico | Dengue virus (DENV) Eastern equine encephalitis virus (EEEV) Venezuelan Equine Encephalitis virus (VEEV) Saint Louis encephalitis virus (SLEV) Vesicular stomatitis, New Jersey virus (VSNJV) Vesicular stomatitis, Indiana (VSIV) Mojui dos Campos virus (MDCV) West Nile virus (WNV) | Primary forest Secondary forest Intervened area Not specified Populated zone | [108,109,113,116,117,118,120,133,138,281] |
| Didelphimorphia | Wild | Brazil Colombia French Guiana Panama Venezuela | Bunyamwera virus (BUNV) Catu virus (CATUV) Dengue virus (DENV) Venezuelan equine encephalitis virus (VEEV) Saint Louis encephalitis virus (SLEV) Vesicular stomatitis, Indiana virus (VSIV) | Primary forest Secondary forest Not specified Intervened area | [67,101,106,113,114,132] |
| Lagomorpha | Wild | Mexico Panama | Melao virus (MELV) Rio Grande virus (RGV) | Not specified | [105,106] |
| Perissodactyla | Domestic | Argentina Brazil Mexico Panama Uruguay Venezuela | ALPHA: alphavirus undetermined Bunyamwera virus (BUNV) Bussuquara virus (BSQV) Eastern equine encephalitis virus (EEEV) Weastern equine encephalitis virus (WEEV) Venezuelan equine encephalitis virus (VEEV) Saint Louis encephalitis virus (SLEV) Fort Sherman virus (FSV) Icoaraci virus (ICOV) Itaqui virus (ITQV) Madrid virus (MADV) Maguari virus (MAGV) Mayaro virus (MAYV) Rocio virus (ROCV) Tacaiuma virus (TCMV) West Nile virus (WNV) Xingu virus (XINV) | Secondary forest Not specified Populated zone | [98,99,100,104,119,122,123,125,126,129,130,131,135] |
| Pilosa | Wild | Brazil Costa Rica French Guiana | Changuinola virus (CGLV) Venezuelan equine encephalitis virus (VEEV) Saint Louis encephalitis virus (SLEV) Vesicular stomatitis, Indiana virus (VSIV) Yellow fever virus (YFV) Ilheus virus (ILHV) Punta Toro virus (PTV) Rio Grande virus (RGV) Tacaiuma virus (TCMV) West Nile virus (WNV) Zika virus (ZIKV) | Primary forest Secondary forest | [102,107,114,124] |
| Non-human primates | Domestic or Zoological settings | Brazil Costa Rica | Dengue virus (DENV) Yellow fever virus (YFV) Flavavirus indeterminado Fort Sherman virus (FSV) Oropouche virus (OROV) | Captive Populated zone | [110,111,134,136] |
| Wild | Argentina Brazil Costa Rica French Guiana Panama Paraguay Venezuela | Bussuquara virus (BSQV) Cacipacore virus (CPCV) Caraparu virus (CARV) Changuinola virus (CGLV) Dengue virus (DENV) Eastern equine encephalitis virus (EEEV) Saint Louis encephalitis virus (SLEV) Vesicular stomatitis, Indiana virus (VSIV) Yellow fever virus (YFV) Icoaraci virus (ICOV) Ilheus virus (ILHV) Mayaro virus (MAYV) Oropouche virus (OROV) Rocio virus (ROCV) UNA virus (UNAV) Utinga virus (UTIV) West Nile virus (WNV) Zika virus (ZIKV) | Primary forest Secondary forest Intervened area Populated zone | [102,103,106,110,111,112,114,115,124,127,128,131,137,139,140,141] | |
| Rodentia | Wild | Brazil Colombia French Guiana Mexico Panama Venezuela | Cache Valley virus (CVV) Changuinola virus (CGLV) Dengue virus (DENV) Eastern equine encephalitis virus (EEEV) Weastern equine encephalitis virus (WEEV)Venezuelan equine encephalitis virus (VEEV) Saint Louis encephalitis virus SLEV) Vesicular stomatitis, New Jersey virus (VSNJV) Vesicular stomatitis, Indiana virus (VSIV) Yellow fever virus (YFV) Gamboa virus (GAMV) Guama virus (GMAV) Ilheus virus (ILHV) Madrid virus (MADV) Melao virus (MELV) Ossa virus (OSSAV) UNA virus (UNAV) West Nile virus (WNV) | Primary forest Secondary forest Captive Not specified Intervened area | [67,101,105,106,113,114,121,132,196] |
Appendix B
| # | Title | Author | Prevalence | Positive | Negative | Type of Area |
|---|---|---|---|---|---|---|
| 1 | Arbovirus serosurvey Orthobunyavirus, Flavivirus, and Alphavirus) in a draft horse population from Santa Fe, Argentina 2013–2016) | Albrieu-Llinás 2021 [123] | TOTAL 222 = INFECTED prevalence FSV = 76.8%, SLEV = 59.6%, WNV = 27.5%, RNV = 7.4% | FSV: 170 SLEV: 132 WNV: 61 RNV: 16 | FSV: 52 SLEV: 90 WNV: 161 RNV: 206 | Deforested fronts |
| 2 | Prevalence of Flavivirus antibodies in Alouatta caraya primate autochthonous of Argentina | Contigiani et al., 2000 [127] | TOTAL 105 = INFECTED prevalence HI 35.23%), NT 32.38%) | 37 34 | 68 71 | Deforested fronts |
| 3 | Serological survey on arbovirus detected in animals in the province of Tucuman, Argentina | De Ruiz Holgado et al., 1967 [129] | Total 75 = infected 58 | 58 | 17 | Deforested fronts |
| 4 | Infection by UNA virus Alphavirus; Togaviridae) and risk factor analysis in black howler monkeys Alouatta caraya) from Paraguay and Argentina | Díaz et al., 2007 [137] | TOTAL 90 = INFECTED 67 46 UNAV y 21 UNAV y el MAYV) | 67 | 23 | Deforested fronts |
| 5 | First isolation of Bunyamwera virus Bunyaviridae family) from horses with neurological disease and an abortion in Argentina | Tauro et al., 2015 [100] | 2 HORSES = 2 INFECTED | 2 | 0 | Deforested fronts |
| 6 | Neutralizing antibodies for orthobunyaviruses in Pantanal, Brazil | Pauvolid-Corrêa et al., 2017 [99] | TOTAL 607 = 373 equidos + 126 bovinos INFECTED TOTAL 499 | 499 | 108 | Deforested fronts |
| 7 | Identification of animal hosts of Fort Sherman virus, a New World zoonotic orthobunyavirus | de Oliveira Filho et al., 2020 [135] | TOTAL 192 = 2 INFECTED | 2 | 190 | Deforested fronts |
| 8 | Yellow fever surveillance challenge: Investigation of a marmoset non-autochthonous case | Fernandes et al., 2020 [134] | TOTAL 1 = 1 INFECTED | 1 | 0 | Vegetation cover |
| 9 | Identification of the encephalitis equine virus, Parana, Brazil | Fernández et al., 2000 [119] | TOTAL 22 = INFECTED 12 | 12 | 10 | Deforested fronts |
| 10 | Detection of antibodies to Oropouche virus in non-human primates in Goiânia City, Goiás | Gibrail et al., 2016 [136] | TOTAL 50 = 2 INFECTED | 2 | 48 | Vegetation cover |
| 11 | Neutralising antibodies for Mayaro virus in Pantanal, Brazil | Pauvolid-Corrêa et al., 2015 [98] | TOTAL 985 = 239 INFECTED | 239 | 746 | Vegetation cover |
| 12 | A Saint Louis encephalitis and Rocio virus serosurvey in Brazilian horses | Silva et al., 2014 [130] | TOTAL 753 = INFECTED 415 | 415 | 338 | Vegetation cover |
| 13 | Serological evidence for Saint Louis encephalitis virus in free-ranging New World monkeys and horses within the upper Paraná River basin region, Southern Brazil | Svoboda et al., 2014 [131] | TOTAL 133 = 30 INFECTED | 30 | 103 | Vegetation cover |
| 14 | Ultrastructural, antigenic and physicochemical characterization of the Mojuí dos Campos Bunyavirus) isolated from bat in the Brazilian Amazon region | Wanzeller et al., 2002 [138] | TOTAL 1 = INFECTED 1 | 1 | 0 | Deforested fronts |
| 15 | A survey to assess potential human disease hazards along proposed sea level canal routes in Panamá and Colombia. V. Arbovirus infection in non-human vertebrates | Srihonges et al., 1974 [106] | TOTAL 2214 = INFECTED 28 | 28 | 2186 | Vegetation cover |
| 16 | Serosurvey of selected arboviral pathogens in free-ranging, two-toed sloths (Choloepus hoffmanni) and three-toed sloths (Bradypus variegatus) in Costa Rica, 2005-07 | Medlin et al., 2016 [107] | TOTAL 109 = 87 INFECTED | 87 | 22 | Vegetation cover |
| 17 | Serologic survey of neotropical bats in Guatemala for virus antibodies | Ubico et al., 1995 [120] | TOTAL 332 = INFECTED 87 | 87 | 245 | Vegetation cover |
| 18 | Dengue infection in neotropical forest mammals | de Thoisy et al., 2009 [113] | TOTAL 616 = INFECTED 92 | 92 | 524 | Vegetation cover |
| 19 | Health evaluation of translocated free-ranging primates in French Guiana | de Thoisy et al., 2001 [112] | TOTAL 141 = INFECTED | Dengue virus II 20/141, Yellow fever virus 47/141, Saint Louis virus 16/141, Mayaro virus 76/141 Average 40 | Dengue virus II 121/141, Yellow fever virus 94/141, Saint Louis virus 125/141, Mayaro virus 65/141 Average 101 | Vegetation cover |
| 20 | Serologic survey for selected arboviruses and other potential pathogens in wildlife from Mexico | Aguirre et al., 1992 [105] | TOTAL 80 = 1 INFECTED | 1 | 79 | Deforested fronts |
| 21 | Serologic survey of domestic animals for zoonotic arbovirus infections in the Lacandón Forest region of Chiapas, Mexico | Ulloa et al., 2003 [104] | TOTA 72 = INFECTED 54 | 54 | 18 | Vegetation cover |
| 22 | Human and Equine Infection with Alphaviruses and Flaviviruses in Panamá during 2010: A Cross-Sectional Study of Household Contacts during an Encephalitis Outbreak | Carrera et al., 2018 [126] | TOTAL 194 = INFECTED 126 | 126 | 68 | Vegetation cover |
| 23 | Seroprevalence of St. Louis encephalitis virus and West Nile virus Flavivirus, Flaviviridae) in horses, Uruguay | Burgueño et al., 2013 [125] | TOTAL 425 = INFCTED 205 | 205 | 220 | Deforested fronts |
| 24 | Ecological studies of enzootic Venezuelan equine encephalitis in north-central Venezuela, 1997–1998 | Salas et al., 2001 [101] | TOTAL 543 = INFECTED 30 | 30 | 513 | Deforested fronts |
| 25 | Studies of arboviruses in Southwestern Venezuela: I. Isolations of Venezuelan and Eastern Equine Encephalitis viruses from sentinel hamsters in the Catatumbo region | Walder et al., 1976 [121] | TOTAL 95 = INFECTED 30 | 30 | 65 | Deforested fronts |
| 26 | Arbovirus studies in the Guajira region of Venezuela: activities of eastern equine encephalitis and Venezuelan equine encephalitis viruses during an interepizootic period | Walder et al., 1984 [122] | TOTAL 64 = INFECTED 60 | 60 | 4 | Deforested fronts |
| 27 | Detection of dengue virus in bat flies Diptera: Streblidae) of common vampire bats, Desmodus rotundus, in Progreso, Hidalgo, Mexico | Abundes-Gallegos et al., 2018 [108] | TOTAL 16 = INFECTED 8 | 8 | 8 | Vegetation cover |
| 28 | Detection of antibodies against Icoaraci, Ilhéus, and Saint Louis Encephalitis arboviruses during yellow fever monitoring surveillance in non-human primates Alouatta caraya) in southern Brazil | Almeida et al., 2019 [124] | TOTAL 26 = INFECTED 5 | 5 | 21 | Deforested fronts |
| 29 | Contrasting sylvatic foci of Venezuelan equine encephalitis virus in northern South America | Barrera et al., 2002 [132] | TOTAL 546 = INFECTED 20 | 20 | 526 | Deforested fronts |
| 30 | Study of Arboviruses in Philander opossum, Didelphis marsupialis and Nectomys rattus captured from forest fragments in the municipality of Belém, Pará, Brazil | Bernal et al., 2021 [67] | TOTAL 14 = INFECTED 8 | 8 | 6 | Deforested fronts |
| 31 | Dengue virus infection in neotropical forest mammals: incidental hosts or potential reservoirs? | Lavergne et al., 2009 [281] | TOTAL 464 = INFECTED 92 | 92 | 372 | Vegetation cover |
| 32 | Two Cases of Natural Infection of Dengue-2 Virus in Bats in the Colombian Caribbean | Calderón et al., 2021 [109] | TOTAL 286 = INFECTED 2 | 2 | 284 | Deforested fronts |
| 33 | Surveillance of arboviruses in primates and sloths in the Atlantic Forest, Bahia, Brazil | Catenacci et al., 2018 [102] | TOTAL 139 = INFECTED 25 | 25 | 114 | Vegetation cover |
| 34 | Flaviviruses infections in neotropical primates suggest long-term circulation of Saint Louis Encephalitis and Dengue virus spillback in socioeconomic regions with high numbers of Dengue human cases in Costa Rica | Chaves et al., 2020 [110] | TOTAL 86 = INFECTED 35 | 35 | 51 | Vegetation cover |
| 35 | Serosurvey of Nonhuman Primates in Costa Rica at the Human–Wildlife Interface Reveals High Exposure to Flaviviruses | Chaves et al., 2021 [111] | TOTAL 86 = INFECTED 38 | 38 | 48 | Vegetation cover |
| 36 | Immunity to yellow fever, Oropouche and Saint Louis viruses in a wild howler monkey | de Almeida et al., 2016 [128] | TOTAL 1 = INFECTED 1 | 1 | 0 | Deforested fronts |
| 37 | Detection of a novel African-lineage-like Zika virus naturally infecting free-living neotropical primates in Southern Brazil | de Almeida et al., 2019 [140] | TOAL 50 = INFECTED 9 | 9 | 41 | Vegetation cover |
| 38 | Wild terrestrial rainforest mammals as potential reservoirs for flaviviruses yellow fever, dengue 2 and St Louis encephalitis viruses) in French Guiana | De Thoisy, B Dussart, Philippe Kazanji, M. 2004 [114] | TOTAL 574= INFECTED 50 | 50 | 524 | Vegetation cover |
| 39 | Zika Virus in peridomestic neotropical primates, Northeast Brazil | Favoretto, Silvana et al., 2019 [141] | TOTAL 132 = INFECTED 9 | 9 | 123 | Deforested fronts |
| 40 | Detection of antibodies against flavivirus over time in wild non-human primates from the lowlands of Costa Rica | Dolz, Gaby et al., 2019 [115] | TOTAL 209 = INFECTED 53 | 53 | 156 | Deforested fronts |
| 41 | Serologic evidence of flavivirus infection in bats in the Yucatan Peninsula of Mexico | Machain-Williams, Carlos et al., 2013 [116] | TOTAL 140 = infected 26 | 26 | 114 | Deforested fronts |
| 42 | Neotropical bats that co-habit with humans function as dead-end hosts for dengue virus | Vicente-Santos, Amanda et al., 2017 [118] | TOTAL 318 = INFECTED 28 | 28 | 290 | Deforested fronts |
| 43 | Dengue virus in bats from southeastern Mexico | Sotomayor-Bonilla, Jesús et al., 2014 [117] | TOTAL 146 = INFECTED 79 | 79 | 67 | Vegetation cover |
| 44 | Isolation of Madre de Dios Virus Orthobunyavirus; Bunyaviridae), an Oropouche virus species reassortant, from a monkey in Venezuela | Navarro et al., 2016 [139] | TOTAL 2 = INFECTED 1 | 1 | 1 | Deforested fronts |
| 45 | Eco-epidemiology of the Venezuelan equine encephalitis virus in bats of Cordoba and Sucre, Colombia | Guzmán, Camilo et al., 2019 [133] | TOTAL 286 = INFECTED 2 | 2 | 284 | Deforested fronts |
| 46 | Detection of the mosquito-borne flaviviruses, West Nile, dengue, Saint Louis encephalitis, Ilheus, Bussuquara, and yellow fever in free-ranging black howlers Alouatta caraya) of northeastern Argentina | Morales et al., 2017 [103] | TOTAL 108 = INFECTED 70 | 70 | 38 | Deforested fronts |
References
- Beech, E.; Rivers, M.; Oldfield, S.; Smith, P. GlobalTreeSearch: The first complete global database of tree species and country distributions. J. Sustain. For. 2017, 36, 454–489. [Google Scholar] [CrossRef]
- Dick, C.W.; Pennington, R.T. History and geography of Neotropical tree diversity. Annu. Rev. Ecol. Evol. Syst. 2019, 50, 279–301. [Google Scholar] [CrossRef]
- Burgin, C.J.; Colella, J.P.; Kahn, P.L.; Upham, N.S. How many species of mammals are there? J. Mammal. 2018, 99, 1–14. [Google Scholar] [CrossRef]
- Patterson, B.D. Patterns and trends in the discovery of new Neotropical mammals. Divers. Distrib. 2000, 6, 145–151. [Google Scholar] [CrossRef]
- Ripple, W.J.; Newsome, T.M.; Wolf, C.; Dirzo, R.; Everatt, K.T.; Galetti, M.; Hayward, M.W.; Kerley, G.I.; Levi, T.; Lindsey, P.A. Collapse of the world’s largest herbivores. Sci. Adv. 2015, 1, e1400103. [Google Scholar] [CrossRef] [PubMed]
- Davidson, A.D.; Detling, J.K.; Brown, J.H. Ecological roles and conservation challenges of social, burrowing, herbivorous mammals in the world’s grasslands. Front. Ecol. Environ. 2012, 10, 477–486. [Google Scholar] [CrossRef]
- Johnson, S.D.; Pauw, A.; Midgley, J. Rodent pollination in the African lily Massonia depressa (Hyacinthaceae). Am. J. Bot. 2001, 88, 1768–1773. [Google Scholar] [CrossRef]
- Fuzessy, L.F.; Janson, C.H.; Silveira, F.A. How far do Neotropical primates disperse seeds? Am. J. Primatol. 2017, 79, e22659. [Google Scholar] [CrossRef]
- Ripple, W.J.; Wolf, C.; Newsome, T.M.; Hoffmann, M.; Wirsing, A.J.; McCauley, D.J. Extinction risk is most acute for the world’s largest and smallest vertebrates. Proc. Natl. Acad. Sci. USA 2017, 114, 10678–10683. [Google Scholar] [CrossRef]
- Lacher, T.E., Jr.; Davidson, A.D.; Fleming, T.H.; Gómez-Ruiz, E.P.; McCracken, G.F.; Owen-Smith, N.; Peres, C.A.; Vander Wall, S.B. The functional roles of mammals in ecosystems. J. Mammal. 2019, 100, 942–964. [Google Scholar] [CrossRef]
- Patterson, B. Fathoming tropical biodiversity: The continuing discovery of Neotropical mammals. Divers. Distrib. 2001, 7, 191–196. [Google Scholar] [CrossRef]
- Morrone, J. Neotropical Biogeography: Regionalization and Evolution; CRC Press & Francis Group: Boca Raton, FL, USA, 2017; p. 282. [Google Scholar]
- Gómez-Ortiz, Y.; Moreno, C. Pilares del ecosistema: Jenga de mamíferos neotropicales. Ecofronteras 2017, 21, 26–30. [Google Scholar]
- Groombridge, B.; Jenkins, M.D.; Jenkins, M. World Atlas of Biodiversity: Earth’s Living Resources in the 21st Century, 1st ed.; UNEP-WCMC by the University of California Press: Berkeley, CA, USA, 2002. [Google Scholar]
- Noguera-Urbano, E.A.; Escalante, T. Areas of Endemism of the Neotropical Mammals (Mammalia). Acta Biol. Colomb. 2015, 20, 47–65. [Google Scholar] [CrossRef]
- Mendoza-Ponce, A.; Corona-Núnez, R.O.; Galicia, L.; Kraxner, F. Identifying hotspots of land use cover change under socioeconomic and climate change scenarios in Mexico. Ambio 2019, 48, 336–349. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Mittermeier, R.A.; Turner, W.R.; Larsen, F.W.; Brooks, T.M.; Gascon, C. Global biodiversity conservation: The critical role of hotspots. In Biodiversity Hotspots; Springer: Berlin/Heidelberg, Germany, 2011; pp. 3–22. [Google Scholar]
- Ceballos, G.; Ehrlich, P.R.; Dirzo, R. Biological annihilation via the ongoing sixth mass extinction signaled by vertebrate population losses and declines. Proc. Natl. Acad. Sci. USA 2017, 114, E6089–E6096. [Google Scholar] [CrossRef]
- Bishop, P.; Angulo, A.; Lewis, J.; Moore, R.; Rabb, G.; Moreno, J.G. The Amphibian Extinction Crisis-what will it take to put the action into the Amphibian Conservation Action Plan? SAPIENS Surv. Perspect. Integr. Environ. Soc. 2012, 5, 97–111. [Google Scholar]
- Reisen, W.K. Landscape epidemiology of vector-borne diseases. Annu. Rev. Entomol. 2010, 55, 461–483. [Google Scholar] [CrossRef]
- Walls, S.C.; Barichivich, W.J.; Brown, M.E. Drought, deluge and declines: The impact of precipitation extremes on amphibians in a changing climate. Biology 2013, 2, 399–418. [Google Scholar] [CrossRef]
- Lawler, J.J.; Shafer, S.L.; White, D.; Kareiva, P.; Maurer, E.P.; Blaustein, A.R.; Bartlein, P.J. Projected climate-induced faunal change in the Western Hemisphere. Ecology 2009, 90, 588–597. [Google Scholar] [CrossRef]
- Mccain, C.M.; King, S.R. Body size and activity times mediate mammalian responses to climate change. Glob. Change Biol. 2014, 20, 1760–1769. [Google Scholar] [CrossRef] [PubMed]
- Lovegrove, B.G.; Canale, C.; Levesque, D.; Fluch, G.; Řeháková-Petrů, M.; Ruf, T. Are tropical small mammals physiologically vulnerable to Arrhenius effects and climate change? Physiol. Biochem. Zool. 2014, 87, 30–45. [Google Scholar] [CrossRef] [PubMed]
- Arredondo-García, J.; Méndez-Herrera, A.; Medina-Cortina, H. Arbovirus en Latinoamérica. Acta Pediátr. Méx. 2016, 37, 111–131. [Google Scholar] [CrossRef]
- Young, P.; Ng, L.; Hall, R.; Smith, D.; Johansen, C. Arbovirus infection. In Manson’s Tropical Diseases; Farrar, J., Hotez, P.J., Junghanss, T., Kang, G., Lalloo, D., White, N.J., Eds.; Elsevier: Amsterdam, The Netherlands, 2013; pp. 129–161. [Google Scholar]
- Goodman, A.G.; Rasmussen, A.L. Host-pathogen interactions during arboviral infections. Front. Cell. Infect. Microbiol. 2019, 9, 77. [Google Scholar] [CrossRef] [PubMed]
- Hanley, K.A.; Weaver, S.C. Arbovirus evolution. In Origin and Evolution of Viruses, 2nd ed.; Domingo, E., Parrish, C.R., Holland, J.J., Eds.; Elsevier: New York, NY, USA, 2008; pp. 351–391. [Google Scholar]
- Stanley, N. Reoviridae: Orbivirus and reovirus infections of mammals and birds. In Comparative Diagnosis of Viral Diseases; Elsevier: Amsterdam, The Netherlands, 2012; Volume 4, pp. 67–104. [Google Scholar]
- Bamford, D.; Zuckerman, M. Encyclopedia of Virology; Academic Press: Amsterdam, The Netherlands, 2021. [Google Scholar]
- Brès, P. Impact of arboviruses on human and animal health. In The Arboviruses: Epidemiology and Ecology; CRC Press: Boca Raton, FL, USA, 2020; pp. 1–18. [Google Scholar]
- Artsob, H.; Lindsay, R.; Drebot, M. Arboviruses. In International Encyclopedia of Public Health, 2nd ed.; Quah, S.R., Ed.; Academic Press: Oxford, UK, 2017; pp. 154–160. [Google Scholar]
- Kenney, J.L.; Brault, A.C. The role of environmental, virological and vector interactions in dictating biological transmission of arthropod-borne viruses by mosquitoes. Adv. Virus Res. 2014, 89, 39–83. [Google Scholar] [CrossRef]
- Olive, M.-M.; Goodman, S.M.; Reynes, J.-M. The role of wild mammals in the maintenance of Rift Valley fever virus. J. Wildl. Dis. 2012, 48, 241–266. [Google Scholar] [CrossRef]
- Quinnell, R.J.; Courtenay, O. Transmission, reservoir hosts and control of zoonotic visceral leishmaniasis. Parasitology 2009, 136, 1915–1934. [Google Scholar] [CrossRef]
- Kuno, G.; Chang, G.-J.J. Biological transmission of arboviruses: Reexamination of and new insights into components, mechanisms, and unique traits as well as their evolutionary trends. Clin. Microbiol. Rev. 2005, 18, 608–637. [Google Scholar] [CrossRef]
- Akpan, G.E.; Adepoju, K.A.; Oladosu, O.R. Potential distribution of dominant malaria vector species in tropical region under climate change scenarios. PLoS ONE 2019, 14, e0218523. [Google Scholar] [CrossRef]
- Hubálek, Z.; Rudolf, I.; Nowotny, N. Arboviruses pathogenic for domestic and wild animals. Adv. Virus Res. 2014, 89, 201–275. [Google Scholar] [CrossRef]
- Schneider, B.S.; Higgs, S. The enhancement of arbovirus transmission and disease by mosquito saliva is associated with modulation of the host immune response. Trans. R. Soc. Trop. Med. Hyg. 2008, 102, 400–408. [Google Scholar] [CrossRef] [PubMed]
- Liang, G.; Gao, X.; Gould, E. Factors responsible for the emergence of arboviruses; strategies, challenges and limitations for their control. Emerg. Microbes Infect. 2015, 4, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Sotomayor-Bonilla, J.; Tolsá-García, M.J.; García-Peña, G.E.; Santiago-Alarcon, D.; Mendoza, H.; Alvarez-Mendizabal, P.; Rico-Chávez, O.; Sarmiento-Silva, R.E.; Suzán, G. Insights into the host specificity of mosquito-borne flaviviruses infecting wild mammals. EcoHealth 2019, 16, 726–733. [Google Scholar] [CrossRef] [PubMed]
- Nene, V.; Wortman, J.R.; Lawson, D.; Haas, B.; Kodira, C.; Tu, Z.J.; Loftus, B.; Xi, Z.; Megy, K.; Grabherr, M. Genome sequence of Aedes aegypti, a major arbovirus vector. Science 2007, 316, 1718–1723. [Google Scholar] [CrossRef]
- San Miguel, L.G.; Sierra, M.J.; Vazquez, A.; Fernandez-Martínez, B.; Molina, R.; Sanchez-Seco, M.P.; Lucientes, J.; Figuerola, J.; de Ory, F.; Monge, S. Enfermedades asociadas a flebovirus trasmitidos por flebótomos:¿ Qué riesgo tenemos en Espana? Enferm. Infecc. Microbiol. Clín. 2020, 39, 345–351. [Google Scholar] [CrossRef]
- Acevedo, M.d.l.Á.; Arrivillaga, J. Eco-epidemiología de flebovirus (Bunyaviridae, Phlebovirus) transmitidos por flebótomos (Psychodidae, Phlebotominae). Bol. Malariol. Salud Ambient. 2008, 48, 3–16. [Google Scholar]
- Higgs, S.; Vanlandingham, D.L. Influences of arthropod vectors on encephalitic arboviruses. In Neurotropic Viral Infections; Springer: Berlin/Heidelberg, Germany, 2016; Volume 2, pp. 371–401. [Google Scholar]
- Gubler, D. Human arbovirus infections worldwide. Ann. N. Y. Acad. Sci. 2001, 951, 13–24. [Google Scholar] [CrossRef]
- Labuda, M.; Nuttall, P. Viruses transmitted by ticks. In Ticks: Biology, Disease and Control; CABI: Cambridge, UK, 2008; pp. 253–280. [Google Scholar]
- Woolhouse, M.E.; Gowtage-Sequeria, S. Host range and emerging and reemerging pathogens. Emerg. Infect. Dis. 2005, 11, 1842. [Google Scholar] [CrossRef]
- Weaver, S.C. Urbanization and geographic expansion of zoonotic arboviral diseases: Mechanisms and potential strategies for prevention. Trends Microbiol. 2013, 21, 360–363. [Google Scholar] [CrossRef]
- Wimberly, M.C.; Davis, J.K.; Evans, M.V.; Hess, A.; Newberry, P.M.; Solano-Asamoah, N.; Murdock, C.C. Land cover affects microclimate and temperature suitability for arbovirus transmission in an urban landscape. PLoS Negl. Trop. Dis. 2020, 14, e0008614. [Google Scholar] [CrossRef]
- Gould, E.; Pettersson, J.; Higgs, S.; Charrel, R.; De Lamballerie, X. Emerging arboviruses: Why today? One Health 2017, 4, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Keesing, F.; Belden, L.K.; Daszak, P.; Dobson, A.; Harvell, C.D.; Holt, R.D.; Hudson, P.; Jolles, A.; Jones, K.E.; Mitchell, C.E. Impacts of biodiversity on the emergence and transmission of infectious diseases. Nature 2010, 468, 647–652. [Google Scholar] [CrossRef] [PubMed]
- Suzán, G.; Marcé, E.; Giermakowski, J.T.; Mills, J.N.; Ceballos, G.; Ostfeld, R.S.; Armién, B.; Pascale, J.M.; Yates, T.L. Experimental evidence for reduced rodent diversity causing increased hantavirus prevalence. PLoS ONE 2009, 4, e5461. [Google Scholar] [CrossRef]
- Gibb, R.; Redding, D.W.; Chin, K.Q.; Donnelly, C.A.; Blackburn, T.M.; Newbold, T.; Jones, K.E. Zoonotic host diversity increases in human-dominated ecosystems. Nature 2020, 584, 398–402. [Google Scholar] [CrossRef] [PubMed]
- Gottdenker, N.L.; Streicker, D.G.; Faust, C.L.; Carroll, C. Anthropogenic land use change and infectious diseases: A review of the evidence. EcoHealth 2014, 11, 619–632. [Google Scholar] [CrossRef] [PubMed]
- Lowe, R.; Lee, S.; Lana, R.M.; Codeço, C.T.; Castro, M.C.; Pascual, M. Emerging arboviruses in the urbanized Amazon rainforest. Br. Med. J. 2020, 371, m4385. [Google Scholar] [CrossRef]
- Sang, R.; Lutomiah, J.; Said, M.; Makio, A.; Koka, H.; Koskei, E.; Nyunja, A.; Owaka, S.; Matoke-Muhia, D.; Bukachi, S. Effects of irrigation and rainfall on the population dynamics of Rift Valley fever and other arbovirus mosquito vectors in the epidemic-prone Tana River County, Kenya. J. Med. Entomol. 2017, 54, 460–470. [Google Scholar] [CrossRef]
- Huang, Y.-J.S.; Higgs, S.; Vanlandingham, D.L. Arbovirus-mosquito vector-host interactions and the impact on transmission and disease pathogenesis of arboviruses. Front. Microbiol. 2019, 10, 22. [Google Scholar] [CrossRef]
- Gould, E.A.; Higgs, S. Impact of climate change and other factors on emerging arbovirus diseases. Trans. R. Soc. Trop. Med. Hyg. 2009, 103, 109–121. [Google Scholar] [CrossRef]
- Yuill, T. The ecology of tropical arthropod-borne viruses. Annu. Rev. Ecol. Syst. 1986, 17, 189–219. [Google Scholar] [CrossRef]
- Reisen, F.; Duran, S.M.; Flannigan, M.; Elliott, C.; Rideout, K. Wildfire smoke and public health risk. Int. J. Wildland Fire 2015, 24, 1029–1044. [Google Scholar] [CrossRef]
- Guth, S.; Hanley, K.A.; Althouse, B.M.; Boots, M. Ecological processes underlying the emergence of novel enzootic cycles: Arboviruses in the neotropics as a case study. PLoS Negl. Trop. Dis. 2020, 14, e0008338. [Google Scholar] [CrossRef] [PubMed]
- Young, P.R. Arboviruses: A Family on the Move. Adv. Exp. Med. Biol. 2018, 1062, 1–10. [Google Scholar] [CrossRef]
- Luis, A.D.; Hayman, D.T.; O’Shea, T.J.; Cryan, P.M.; Gilbert, A.T.; Pulliam, J.R.; Mills, J.N.; Timonin, M.E.; Willis, C.K.; Cunningham, A.A. A comparison of bats and rodents as reservoirs of zoonotic viruses: Are bats special? Proc. R. Soc. B Biol. Sci. 2013, 280, 20122753. [Google Scholar] [CrossRef]
- Feldhamer, G.A.; Drickamer, L.C.; Vessey, S.H.; Merritt, J.F.; Krajewski, C. Mammalogy: Adaptation, Diversity, Ecology, 4th ed.; Press, J.H.U., Ed.; The Johns Hopkins University Press: Baltimore, MD, USA, 2015; p. 643. [Google Scholar]
- Bernal, M.K.M.; Chiang, J.O.; Mendes, F.F.; Andrade, S.L.d.S.; Silva, S.K.S.M.d.; Pereira, W.L.A. Study of Arboviruses in Philander opossum, Didelphis marsupialis and Nectomys rattus captured from forest fragments in the municipality of Belém, Pará, Brazil. Ciência Rural 2021, 51. [Google Scholar] [CrossRef]
- Morand, S.; Krasnov, B.R.; Poulin, R. (Eds.) Micromammals and Macroparasites: From Evolutionary Ecology to Management; Springer: Tokyo, Japan, 2007; p. 647. [Google Scholar]
- Faust, C.L.; McCallum, H.I.; Bloomfield, L.S.; Gottdenker, N.L.; Gillespie, T.R.; Torney, C.J.; Dobson, A.P.; Plowright, R.K. Pathogen spillover during land conversion. Ecol. Lett. 2018, 21, 471–483. [Google Scholar] [CrossRef]
- Plowright, R.K.; Parrish, C.R.; McCallum, H.; Hudson, P.J.; Ko, A.I.; Graham, A.L.; Lloyd-Smith, J.O. Pathways to zoonotic spillover. Nat. Rev. Microbiol. 2017, 15, 502–510. [Google Scholar] [CrossRef] [PubMed]
- Ellwanger, J.H.; Chies, J.A.B. Zoonotic spillover and emerging viral diseases–time to intensify zoonoses surveillance in Brazil. Braz. J. Infect. Dis. 2018, 22, 76–78. [Google Scholar] [CrossRef]
- Johnson, C.K.; Hitchens, P.L.; Evans, T.S.; Goldstein, T.; Thomas, K.; Clements, A.; Joly, D.O.; Wolfe, N.D.; Daszak, P.; Karesh, W.B. Spillover and pandemic properties of zoonotic viruses with high host plasticity. Sci. Rep. 2015, 5, 14830. [Google Scholar] [CrossRef]
- Khalil, H.; Ecke, F.; Evander, M.; Magnusson, M.; Hörnfeldt, B. Declining ecosystem health and the dilution effect. Sci. Rep. 2016, 6, 31314. [Google Scholar] [CrossRef]
- Wasay, M.; Khatri, I.A.; Abd-Allah, F. Arbovirus infections of the nervous system: Current trends and future threats. Neurology 2015, 84, 421–423. [Google Scholar] [CrossRef] [PubMed]
- Weaver, S.; Reisen, W. Present and future arboviral threats. Antivir. Res. 2010, 85, 328–345. [Google Scholar] [CrossRef] [PubMed]
- Arrivillaga, J.; Caraballo, V. Medicina de la Conservación. Rev. Bioméd. 2009, 20, 55–67. [Google Scholar]
- Daszak, P.; Tabor, G.M.; Kilpatrick, A.M.; Epstein, J.; Plowright, R. Conservation medicine and a new agenda for emerging diseases. Ann. N. Y. Acad. Sci. 2004, 1026, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Frank, D. One world, one health, one medicine. Can. Vet. J. 2008, 49, 1063. [Google Scholar] [PubMed]
- Benelli, G.; Senthil-Nathan, S. Together in the Fight against Arthropod-Borne Diseases: A One Health Perspective. Int. J. Environ. Res. Public Health 2019, 16, 4876. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Moher, D. Updating guidance for reporting systematic reviews: Development of the PRISMA 2020 statement. J. Clin. Epidemiol. 2021, 134, 103–112. [Google Scholar] [CrossRef]
- Yepes-Nuñez, J.J.; Urrútia, G.; Romero-García, M.; Alonso-Fernández, S. Declaración PRISMA 2020: Una guía actualizada para la publicación de revisiones sistemáticas. Rev. Esp. Cardiol. 2021, 74, 790–799. [Google Scholar] [CrossRef]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; Group, T.P. Preferred Reporting Items for Systematic Reviews and Meta-Analyses: The PRISMA Statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef]
- Liberati, A.; Altman, D.G.; Tetzlaff, J.; Mulrow, C.; Gøtzsche, P.C.; Ioannidis, J.P.; Clarke, M.; Devereaux, P.J.; Kleijnen, J.; Moher, D. The PRISMA statement for reporting systematic reviews and meta-analyses of studies that evaluate health care interventions: Explanation and elaboration. J. Clin. Epidemiol. 2009, 62, e1–e34. [Google Scholar] [CrossRef]
- IBM. IBM SPSS Statistics for Windows; Version 20.0; IBM Corp.: Armonk, NY, USA, 2021. [Google Scholar]
- Torres-Fonseca, A.; López-Hernández, D. Criterios para publicar artículos de revisión sistemática. Rev. Espec. Méd.-Qúir. 2014, 19, 393–399. [Google Scholar]
- Kazmi, R.; Jawawi, D.N.; Mohamad, R.; Ghani, I. Effective regression test case selection: A systematic literature review. ACM Comput. Surv. 2017, 50, 1–32. [Google Scholar] [CrossRef]
- Breslow, N. A generalized Kruskal-Wallis test for comparing K samples subject to unequal patterns of censorship. Biometrika 1970, 57, 579–594. [Google Scholar] [CrossRef]
- Vargha, A.; Delaney, H.D. The Kruskal-Wallis test and stochastic homogeneity. J. Educ. Behav. Stat. 1998, 23, 170–192. [Google Scholar] [CrossRef]
- ESRI. ArcGIS Desktop: Release 10; Environmental Systems Research Institute: Redlands, CA, USA, 2011. [Google Scholar]
- Bastian, M.; Heymann, S.; Jacomy, M. Gephi: An open source software for exploring and manipulating networks. In Proceedings of the Third International AAAI Conference on Weblogs and Social Media, San Jose, CA, USA, 17–20 May 2009. [Google Scholar]
- Chao, A.; Chazdon, R.L.; Colwell, R.K.; Shen, T.J. A new statistical approach for assessing similarity of species composition with incidence and abundance data. Ecol. Lett. 2005, 8, 148–159. [Google Scholar] [CrossRef]
- Jules, P.R.; Olivier, O.G.; Ndongo, D. Utilisation des macrophytes comme descripteurs de la qualité des eaux de la rivière Kondi dans la ville de Douala (Cameroun-Afrique Centrale). J. Appl. Biosci. 2012, 53, 3797–3811. [Google Scholar]
- Magurran, A.E. Measuring Biological Diversity; John Wiley & Sons: Hoboken, NJ, USA, 2013. [Google Scholar]
- Arévalo-Cepeda, Y.; Bogotá-Sierra, M.; Cortés-Pirazán, L.; Sierra-Vargas, A.; Moscoso, J.; Méndeza, W. Virus mayaro: Un arbovirus que amenaza a Colombia. Números 2020, 5, 93–103. [Google Scholar]
- WWF. Global Observation and Biodiversity Information Portal. Available online: https://globil.panda.org/apps/panda::deforestation-fronts-2020-web-map-app/about (accessed on 6 October 2021).
- Pacheco, P.; Mo, K.; Dudley, N.; Shapiro, A.; Aguilar-Amuchastegui, N.; Ling, P.Y.; Anderson, C.; Marx, A. Deforestation Fronts: Drivers and Responses in a Changing World; WWF: Gland, Switzerland, 2021. [Google Scholar]
- Ortíz Agui, M.; Villar Carbajal, E.I.; Llanos de Tarazona, M.I. Estrategia comunicativa orientada a la reducción de la exposición a factores de riesgo de arbovirosis. MediSur 2020, 18, 161–170. [Google Scholar]
- Pauvolid-Corrêa, A.; Juliano, R.S.; Campos, Z.; Velez, J.; Nogueira, R.M.; Komar, N. Neutralising antibodies for Mayaro virus in Pantanal, Brazil. Memórias Inst. Oswaldo Cruz 2015, 110, 125–133. [Google Scholar] [CrossRef]
- Pauvolid-Corrêa, A.; Campos, Z.; Soares, R.; Nogueira, R.M.R.; Komar, N. Neutralizing antibodies for orthobunyaviruses in Pantanal, Brazil. PLoS Negl. Trop. Dis. 2017, 11, e0006014. [Google Scholar] [CrossRef]
- Tauro, L.B.; Rivarola, M.E.; Lucca, E.; Mariño, B.; Mazzini, R.; Cardoso, J.F.; Barrandeguy, M.E.; Teixeira Nunes, M.R.; Contigiani, M.S. First isolation of Bunyamwera virus (Bunyaviridae family) from horses with neurological disease and an abortion in Argentina. Vet. J. 2015, 206, 111–114. [Google Scholar] [CrossRef] [PubMed]
- Salas, R.A.; Garcia, C.Z.; Liria, J.; Barrera, R.; Navarro, J.C.; Medina, G.; Vasquez, C.; Fernandez, Z.; Weaver, S.C. Ecological studies of enzootic Venezuelan equine encephalitis in north-central Venezuela, 1997-1998. Am. J. Trop. Med. Hyg. 2001, 64, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Catenacci, L.S.; Ferreira, M.; Martins, L.; De Vleeschouwer, K.; Cassano, C.; Oliveira, L.; Canale, G.; Deem, S.; Tello, J.; Parker, P. Surveillance of arboviruses in primates and sloths in the Atlantic Forest, Bahia, Brazil. EcoHealth 2018, 15, 777–791. [Google Scholar] [CrossRef] [PubMed]
- Morales, M.A.; Fabbri, C.M.; Zunino, G.E.; Kowalewski, M.M.; Luppo, V.C.; Enría, D.A.; Levis, S.C.; Calderón, G.E. Detection of the mosquito-borne flaviviruses, West Nile, dengue, Saint Louis encephalitis, Ilheus, Bussuquara, and yellow fever in free-ranging black howlers (Alouatta caraya) of northeastern Argentina. PLoS Negl. Trop. Dis. 2017, 11, e0005351. [Google Scholar] [CrossRef]
- Ulloa, A.; Langevin, S.A.; Mendez-Sanchez, J.D.; Arredondo-Jimenez, J.I.; Raetz, J.L.; Powers, A.M.; Villarreal-Treviño, C.; Gubler, D.J.; Komar, N. Serologic survey of domestic animals for zoonotic arbovirus infections in the Lacandón Forest region of Chiapas, Mexico. Vector Borne Zoonotic Dis. 2003, 3, 3–9. [Google Scholar] [CrossRef]
- Aguirre, A.A.; McLean, R.G.; Cook, R.S.; Quan, T.J. Serologic survey for selected arboviruses and other potential pathogens in wildlife from Mexico. J. Wildl. Dis. 1992, 28, 435–442. [Google Scholar] [CrossRef]
- Srihonges, S.; Galindo, P.; Eldridge, B.F.; Young, D.G.; Gerhardt, R.R. A survey to assess potential human disease hazards along proposed sea level canal routes in Panamá and Colombia. V. Arbovirus infection in non-human vertebrates. Mil. Med. 1974, 141, 449–453. [Google Scholar] [CrossRef]
- Medlin, S.; Deardorff, E.R.; Hanley, C.S.; Vergneau-Grosset, C.; Siudak-Campfield, A.; Dallwig, R.; da Rosa, A.T.; Tesh, R.B.; Martin, M.P.; Weaver, S.C.; et al. Serosurvey of selected arboviral pathogens in free-ranging, two-toed sloths (Choloepus hoffmanni) and three-toed sloths (Bradypus variegatus) in Costa Rica, 2005–2007. J. Wildl. Dis. 2016, 52, 883–892. [Google Scholar] [CrossRef]
- Abundes-Gallegos, J.; Salas-Rojas, M.; Galvez-Romero, G.; Perea-Martínez, L.; Obregón-Morales, C.Y.; Morales-Malacara, J.B.; Chomel, B.B.; Stuckey, M.J.; Moreno-Sandoval, H.; García-Baltazar, A. Detection of dengue virus in bat flies (Diptera: Streblidae) of common vampire bats, Desmodus rotundus, in Progreso, Hidalgo, Mexico. Vector-Borne Zoonotic Dis. 2018, 18, 70–73. [Google Scholar] [CrossRef]
- Calderón, A.; Guzmán, C.; Oviedo-Socarras, T.; Mattar, S.; Rodríguez, V.; Castañeda, V.; Moraes Figueiredo, L.T. Two Cases of Natural Infection of Dengue-2 Virus in Bats in the Colombian Caribbean. Trop. Med. Infect. Dis. 2021, 6, 35. [Google Scholar] [CrossRef]
- Chaves, A.; Piche-Ovares, M.; Corrales, E.; Suzán, G.; Moreira-Soto, A.; Gutiérrez-Espeleta, G.A. Flaviviruses infections in neotropical primates suggest long-term circulation of Saint Louis Encephalitis and Dengue virus spillback in socioeconomic regions with high numbers of Dengue human cases in Costa Rica. Anat. Morphol. 2020. preprints. [Google Scholar]
- Chaves, A.; Piche-Ovares, M.; Ibarra-Cerdeña, C.N.; Corrales-Aguilar, E.; Suzán, G.; Moreira-Soto, A.; Gutiérrez-Espeleta, G.A. Serosurvey of Nonhuman Primates in Costa Rica at the Human–Wildlife Interface Reveals High Exposure to Flaviviruses. Insects 2021, 12, 554. [Google Scholar] [CrossRef] [PubMed]
- De Thoisy, B.; Vogel, I.; Reynes, J.M.; Pouliquen, J.F.; Carme, B.; Kazanji, M.; Vié, J.C. Health evaluation of translocated free-ranging primates in French Guiana. Am. J. Primatol. 2001, 54, 1–16. [Google Scholar] [CrossRef]
- De Thoisy, B.; Lacoste, V.; Germain, A.; Muñoz-Jordán, J.; Colón, C.; Mauffrey, J.F.; Delaval, M.; Catzeflis, F.; Kazanji, M.; Matheus, S.; et al. Dengue infection in neotropical forest mammals. Vector Borne Zoonotic Dis. 2009, 9, 157–170. [Google Scholar] [CrossRef] [PubMed]
- De Thoisy, B.; Dussart, P.; Kazanji, M. Wild terrestrial rainforest mammals as potential reservoirs for flaviviruses (yellow fever, dengue 2 and St Louis encephalitis viruses) in French Guiana. Trans. R. Soc. Trop. Med. Hyg. 2004, 98, 409–412. [Google Scholar] [CrossRef]
- Dolz, G.; Chaves, A.; Gutiérrez-Espeleta, G.A.; Ortiz-Malavasi, E.; Bernal-Valle, S.; Herrero, M.V. Detection of antibodies against flavivirus over time in wild non-human primates from the lowlands of Costa Rica. PLoS ONE 2019, 14, e0219271. [Google Scholar] [CrossRef]
- Machain-Williams, C.; López-Uribe, M.; Talavera-Aguilar, L.; Carrillo-Navarrete, J.; Vera-Escalante, L.; Puerto-Manzano, F.; Ulloa, A.; Farfán-Ale, J.A.; Garcia-Rejon, J.; Blitvich, B.J. Serologic evidence of flavivirus infection in bats in the Yucatan Peninsula of Mexico. J. Wildl. Dis. 2013, 49, 684–689. [Google Scholar] [CrossRef]
- Sotomayor-Bonilla, J.; Chaves, A.; Rico-Chávez, O.; Rostal, M.K.; Ojeda-Flores, R.; Salas-Rojas, M.; Aguilar-Setien, Á.; Ibáñez-Bernal, S.; Barbachano-Guerrero, A.; Gutiérrez-Espeleta, G. Dengue virus in bats from southeastern Mexico. Am. J. Trop. Med. Hyg. 2014, 91, 129. [Google Scholar] [CrossRef]
- Vicente-Santos, A.; Moreira-Soto, A.; Soto-Garita, C.; Chaverri, L.G.; Chaves, A.; Drexler, J.F.; Morales, J.A.; Alfaro-Alarcón, A.; Rodríguez-Herrera, B.; Corrales-Aguilar, E. Neotropical bats that co-habit with humans function as dead-end hosts for dengue virus. PLoS Negl. Trop. Dis. 2017, 11, e0005537. [Google Scholar] [CrossRef]
- Fernández, Z.; Richartz, R.; Travassos da Rosa, A.; Soccol, V.T. Identification of the encephalitis equine virus, Parana, Brazil. Rev. Saude Publica 2000, 34, 232–235. [Google Scholar] [CrossRef]
- Ubico, S.R.; McLean, R.G. Serologic survey of neotropical bats in Guatemala for virus antibodies. J. Wildl. Dis. 1995, 31, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Walder, R.; Suárez, O.M. Studies of arboviruses in Southwestern Venezuela: I. Isolations of Venezuelan and Eastern Equine Encephalitis viruses from sentinel hamsters in the Catatumbo region. Int. J. Epidemiol. 1976, 5, 375–378. [Google Scholar] [CrossRef]
- Walder, R.; Suarez, O.M.; Calisher, C.H. Arbovirus studies in the Guajira region of Venezuela: Activities of eastern equine encephalitis and Venezuelan equine encephalitis viruses during an interepizootic period. Am. J. Trop. Med. Hyg. 1984, 33, 699–707. [Google Scholar] [CrossRef] [PubMed]
- Albrieu-Llinás, G.; Gallardo, R.; Konigheim, B.S.; Quaglia, A.I.; Mariño, B.; Curiotti, J.; Mazzini, R.; Contigiani, M.S. Arbovirus serosurvey (Orthobunyavirus, Flavivirus, and Alphavirus) in a draft horse population from Santa Fe, Argentina (2013–2016). Arch. Virol. 2021, 166, 881–884. [Google Scholar] [CrossRef] [PubMed]
- Almeida, M.A.B.d.; Santos, E.d.; Cardoso, J.d.C.; Noll, C.A.; Lima, M.d.M.; Silva, F.d.A.e.; Ferreira, M.S.; Martins, L.C.; Vasconcelos, P.F.d.C.; Bicca-Marques, J.C. Detection of antibodies against Icoaraci, Ilhéus, and Saint Louis Encephalitis arboviruses during yellow fever monitoring surveillance in non-human primates (Alouatta caraya) in southern Brazil. J. Med. Primatol. 2019, 48, 211–217. [Google Scholar] [CrossRef]
- Burgueño, A.; Spinsanti, L.; Díaz, L.A.; Rivarola, M.E.; Arbiza, J.; Contigiani, M.; Delfraro, A. Seroprevalence of St. Louis encephalitis virus and West Nile virus (Flavivirus, Flaviviridae) in horses, Uruguay. BioMed Res. Int. 2013, 2013, 582957. [Google Scholar] [CrossRef]
- Carrera, J.P.; Bagamian, K.H.; Travassos da Rosa, A.P.; Wang, E.; Beltran, D.; Gundaker, N.D.; Armien, B.; Arroyo, G.; Sosa, N.; Pascale, J.M.; et al. Human and Equine Infection with Alphaviruses and Flaviviruses in Panamá during 2010: A Cross-Sectional Study of Household Contacts during an Encephalitis Outbreak. Am. J. Trop. Med. Hyg. 2018, 98, 1798–1804. [Google Scholar] [CrossRef]
- Contigiani, M.S.; Fernández, C.; Spinsanti, L.I.; Díaz, G.E. Prevalence of Flavivirus antibodies in Alouatta caraya primate autochthonous of Argentina. Medicina 2000, 60, 348–350. [Google Scholar]
- De Almeida, M.; Cardoso, J.C.; Santos, E.; Romano, A.; Chiang, J.O.; Martins, L.C.; da Costa Vasconcelos, P.F.; Marques, J.C.B. Immunity to yellow fever, Oropouche and Saint Louis viruses in a wild howler monkey. Neotrop. Primates 2016, 23, 19–21. [Google Scholar]
- De Ruiz Holgado, A.P.; Raya, J.M.; Sabattini, M.S.; De Nader, O.R.; De Castagnaro, N.R. Serological survey on arbovirus detected in animals in the province of Tucuman, Argentina. Bol. Oficina Sanit. Panam. Pan Am. Sanit. Bur. 1967, 63, 323–329. [Google Scholar]
- Silva, J.R.; Romeiro, M.F.; Souza, W.M.; Munhoz, T.D.; Borges, G.P.; Soares, O.A.; Campos, C.H.; Machado, R.Z.; Silva, M.L.; Faria, J.L.; et al. A Saint Louis encephalitis and Rocio virus serosurvey in Brazilian horses. Rev. Soc. Bras. Med. Trop. 2014, 47, 414–417. [Google Scholar] [CrossRef]
- Svoboda, W.K.; Martins, L.C.; Malanski Lde, S.; Shiozawa, M.M.; Spohr, K.A.; Hilst, C.L.; Aguiar, L.M.; Ludwig, G.; Passos Fde, C.; Silva, L.R.; et al. Serological evidence for Saint Louis encephalitis virus in free-ranging New World monkeys and horses within the upper Paraná River basin region, Southern Brazil. Rev. Soc. Bras. Med. Trop. 2014, 47, 280–286. [Google Scholar] [CrossRef] [PubMed]
- Barrera, R.; Ferro, C.; Navarro, J.-C.; Freier, J.; Liria, J.; Salas, R.; Ahumada, M.; Vasquez, C.; Gonzalez, M.; Kang, W. Contrasting sylvatic foci of Venezuelan equine encephalitis virus in northern South America. Am. J. Trop. Med. Hyg. 2002, 67, 324–334. [Google Scholar] [CrossRef] [PubMed]
- Guzmán, C.; Calderón, A.; Martinez, C.; Oviedo, M.; Mattar, S. Eco-epidemiology of the Venezuelan equine encephalitis virus in bats of Cordoba and Sucre, Colombia. Acta Trop. 2019, 191, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, N.; Guerra, J.M.; Cunha, M.S.; Beraldo, K.R.F.; Ressio, R.A.; Cirqueira, C.D.S.; Agostini, T.L.; de Camargo, J.P.; Landi, N.; Saad, L.D.C.; et al. Yellow fever surveillance challenge: Investigation of a marmoset non-autochthonous case. Acta Trop. 2020, 212, 105702. [Google Scholar] [CrossRef]
- De Oliveira Filho, E.F.; Carneiro, I.O.; Ribas, J.R.L.; Fischer, C.; Marklewitz, M.; Junglen, S.; Netto, E.M.; Franke, C.R.; Drexler, J.F. Identification of animal hosts of Fort Sherman virus, a New World zoonotic orthobunyavirus. Transbound. Emerg. Dis. 2020, 67, 1433–1441. [Google Scholar] [CrossRef]
- Gibrail, M.M.; Fiaccadori, F.S.; Souza, M.; Almeida, T.N.; Chiang, J.O.; Martins, L.C.; Ferreira, M.S.; Cardoso, D. Detection of antibodies to Oropouche virus in non-human primates in Goiânia City, Goiás. Rev. Soc. Bras. Med. Trop. 2016, 49, 357–360. [Google Scholar] [CrossRef]
- Díaz, L.A.; Díaz Mdel, P.; Almirón, W.R.; Contigiani, M.S. Infection by UNA virus (Alphavirus; Togaviridae) and risk factor analysis in black howler monkeys (Alouatta caraya) from Paraguay and Argentina. Trans. R. Soc. Trop. Med. Hyg. 2007, 101, 1039–1041. [Google Scholar] [CrossRef]
- Wanzeller, A.L.; Diniz, J.A.; Gomes, M.L.; Cruz, A.C.; Soares, M.C.; de Souza, W.; Travassos da Rosa, A.P.; Vasconcelos, P.F. Ultrastructural, antigenic and physicochemical characterization of the Mojuí dos Campos (Bunyavirus) isolated from bat in the Brazilian Amazon region. Mem. Inst. Oswaldo Cruz 2002, 97, 307–311. [Google Scholar] [CrossRef]
- Navarro, J.-C.; Giambalvo, D.; Hernandez, R.; Auguste, A.J.; Tesh, R.B.; Weaver, S.C.; Montañez, H.; Liria, J.; Lima, A.; Da Rosa, J.F.S.T. Isolation of Madre de Dios Virus (Orthobunyavirus; Bunyaviridae), an Oropouche virus species reassortant, from a monkey in Venezuela. Am. J. Trop. Med. Hyg. 2016, 95, 328. [Google Scholar] [CrossRef]
- De Almeida, P.R.; Ehlers, L.P.; Demoliner, M.; Eisen, A.K.A.; Girardi, V.; De Lorenzo, C.; Bianchi, M.V.; Mello, L.; Pavarini, S.P.; Driemeier, D. Detection of a novel African-lineage-like Zika virus naturally infecting free-living neotropical primates in Southern Brazil. bioRxiv 2019, 828871. [Google Scholar] [CrossRef]
- Favoretto, S.R.; Araujo, D.B.; Duarte, N.F.; Oliveira, D.B.; Nathalia, G.; Mesquita, F.; Leal, F.; Machado, R.R.; Gaio, F.; Oliveira, W.F. Zika Virus in peridomestic neotropical primates, Northeast Brazil. EcoHealth 2019, 16, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Robert, M.A.; Tinunin, D.T.; Benitez, E.M.; Ludueña-Almeida, F.; Romero, M.; Stewart-Ibarra, A.M.; Estallo, E.L. Arbovirus emergence in temperate climates: The case of Córdoba, Argentina, 2009–2018. bioRxiv 2019, 602003. [Google Scholar] [CrossRef]
- Queiroz, J.T.M.d.; Silva, P.N.; Heller, L. New premises for sanitation in arbovirus infections control in Brazil. Cad. Saude Publica 2020, 36, e00223719. [Google Scholar] [CrossRef]
- Tesh, R.B.; da Rosa, A.P.T.; Guzman, H.; Araujo, T.P.; Xiao, S.-Y. Immunization with heterologous flaviviruses protective against fatal West Nile encephalitis. Emerg. Infect. Dis. 2002, 8, 245. [Google Scholar] [CrossRef]
- Pfeffer, M.; Dobler, G. Emergence of zoonotic arboviruses by animal trade and migration. Parasites Vectors 2010, 3, 35. [Google Scholar] [CrossRef]
- Lounibos, L.P. Invasions by insect vectors of human disease. Annu. Rev. Entomol. 2002, 47, 233–266. [Google Scholar] [CrossRef]
- Kilpatrick, A.M.; Randolph, S.E. Drivers, dynamics, and control of emerging vector-borne zoonotic diseases. Lancet 2012, 380, 1946–1955. [Google Scholar] [CrossRef]
- Mouchet, J. Airport malaria: A rare disease still poorly understood. Eurosurveillance 2000, 5, 75–76. [Google Scholar] [CrossRef]
- Sinti-Hesse, C.; Díaz-Soria, F.; Casanova-Rojas, W.; Carey-Ángeles, C.; Tello-Espinoza, R.; Espinoza, J.; Zevallos, K. Embarcaciones fluviales como medio de dispersión de Aedes aegypti hacia zonas fronterizas de la Amazonia peruana. Rev. Peru. Med. Exp. Salud Pública 2019, 36, 392–399. [Google Scholar] [CrossRef]
- Findlater, A.; Bogoch, I.I. Human mobility and the global spread of infectious diseases: A focus on air travel. Trends Parasitol. 2018, 34, 772–783. [Google Scholar] [CrossRef] [PubMed]
- OMS. Manual de Vigilancia y Control en Puertos, Aeropuertos y Pasos Fronterizos Terrestres; World Helath Organization: Geneva, Switzerlanc, 2016; Volume 96.
- Whiteman, A.; Desjardins, M.R.; Eskildsen, G.A.; Loaiza, J.R. Detecting space-time clusters of dengue fever in Panama after adjusting for vector surveillance data. PLoS Negl. Trop. Dis. 2019, 13, e0007266. [Google Scholar] [CrossRef] [PubMed]
- Piche-Ovares, M. Arbovirus en Costa Rica:¿ A qué nos enfrentamos y cuáles son los retos futuros? Arbovirus in Costa Rica: What we face and what are the future challenges? Rev. Colegio Microb. Quim. Clin. Costa Rica. 2018, 24, 127–144. [Google Scholar]
- Patterson, J.; Sammon, M.; Garg, M. Dengue, Zika and chikungunya: Emerging arboviruses in the New World. West. J. Emerg. Med. 2016, 17, 671. [Google Scholar] [CrossRef] [PubMed]
- Alarcón-Elbal, P. Deforestation and mosquito-borne diseases: Another’wake-up call’to Latin America. InterAm. J. Med. Health 2018, 1, e201801003. [Google Scholar] [CrossRef]
- Mourão, M.P.G.; Bastos, M.d.S.; Figueiredo, R.M.P.d.; Gimaque, J.B.d.L.; Alves, V.d.C.R.; Saraiva, M.d.G.G.; Figueiredo, M.L.G.; Ramasawmy, R.; Nogueira, M.L.; Figueiredo, L.T.M. Arboviral diseases in the Western Brazilian Amazon: A perspective and analysis from a tertiary health & research center in Manaus, State of Amazonas. Rev. Da Soc. Bras. Med. Trop. 2015, 48, 20–26. [Google Scholar] [CrossRef]
- Apex-Brasil. Investigación Y Desarrollo. Available online: http://www.apexbrasil.com.br/es/investigacion-y-desarrollo (accessed on 20 September 2021).
- Rezende, R.B. Análise epidemiológica das arboviroses emergentes e reemergentes no Brasil entre os anos de 2019–2020. Res. Soc. Dev. 2021, 10, e33010212611. [Google Scholar] [CrossRef]
- Souza-Lopes, O.; Sacchetta, L. Isolation of Mucambo virus, a member of the Venezuelan equine encephalitis virus complex in the State of Sâo Paulo, Brasil. Rev. Inst. Med. Trop. Sao Paulo 1978, 20, 82–86. [Google Scholar]
- Martinet, J.-P.; Ferté, H.; Failloux, A.-B.; Schaffner, F.; Depaquit, J. Mosquitoes of north-western Europe as potential vectors of arboviruses: A review. Viruses 2019, 11, 1059. [Google Scholar] [CrossRef]
- Tauil, P.L. Aspectos críticos do controle do dengue no Brasil. Cad. Saude Publica 2002, 18, 867–871. [Google Scholar] [CrossRef]
- Wilder-Smith, A.; Ooi, E.-E.; Horstick, O.; Wills, B. Dengue. Lancet 2019, 393, 350–363. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, L.T.M. Emergent arboviruses in Brazil. Rev. Soc. Bras. Med. Trop. 2007, 40, 224–229. [Google Scholar] [CrossRef] [PubMed]
- Weaver, S.C.; Ferro, C.; Barrera, R.; Boshell, J.; Navarro, J.-C. Venezuelan equine encephalitis. Annu. Rev. Entomol. 2004, 49, 141–174. [Google Scholar] [CrossRef] [PubMed]
- Torres, R.; Samudio, R.; Carrera, J.-P.; Young, J.; Márquez, R.; Hurtado, L.; Weaver, S.; Chaves, L.F.; Tesh, R.; Cáceres, L. Enzootic mosquito vector species at equine encephalitis transmission foci in the República de Panamá. PLoS ONE 2017, 12, e0185491. [Google Scholar] [CrossRef] [PubMed]
- Cupp, E.; Scherer, W.; Ordonez, J. Transmission of Venezuelan encephalitis virus by naturally infected Culex (Melanoconion) opisthopus. Am. J. Trop. Med. Hyg. 1979, 28, 1060–1063. [Google Scholar] [CrossRef]
- Galindo, P.; Grayson, M.A. Culex (Melanoconion) aikenii: Natural vector in Panama of endemic Venezuelan encephalitis. Science 1971, 172, 594–595. [Google Scholar] [CrossRef]
- Pisano, M.B.; Ré, V.E.; Diaz, L.A.; Stein, M.; Dantur Juri, M.J.; Farías, A.A.; Sanchez Seco, M.P.; Tenorio, A.; Almiron, W.R.; Contigiani de Minio, M.S. Actividad de virus del complejo Encefalitis Equina Venezolana (VEEV) en Argentina. Rev. Fac. Cienc. Méd. 2009, 66, 48–51. [Google Scholar]
- Navarro, J.-C.; Medina, G.; Vasquez, C.; Coffey, L.L.; Wang, E.; Suárez, A.; Biord, H.; Salas, M.; Weaver, S.C. Postepizootic persistence of Venezuelan equine encephalitis virus, Venezuela. Emerg. Infect. Dis. 2005, 11, 1907. [Google Scholar] [CrossRef]
- Vargas, D.S.; Jaime, J.; Vera, V.J. Aspectos generales del virus de la encefalitis equina venezolana (VEEV). Orinoquia 2009, 13, 59–67. [Google Scholar]
- Weaver, S. Host range, amplification and arboviral disease emergence. In Infectious Diseases from Nature: Mechanisms of Viral Emergence and Persistence; Springer: Berlin/Heidelberg, Germany, 2005; pp. 33–44. [Google Scholar] [CrossRef]
- Vieira, C.J.d.S.P.; Thies, S.F.; da Silva, D.J.F.; Kubiszeski, J.R.; Barreto, E.S.; de Oliveira Monteiro, H.A.; Mondini, A.; São Bernardo, C.S.; de Morais Bronzoni, R.V. Ecological aspects of potential arbovirus vectors (Diptera: Culicidae) in an urban landscape of Southern Amazon, Brazil. Acta Trop. 2020, 202, 105276. [Google Scholar] [CrossRef]
- Barrera, R.; Torres, N.; Freier, J.E.; Navarro, J.C.; García, C.Z.; Salas, R.; Vasquez, C.; Weaver, S.C. Characterization of enzootic foci of Venezuelan equine encephalitis virus in western Venezuela. Vector Borne Zoonotic Dis. 2001, 1, 219–230. [Google Scholar] [CrossRef] [PubMed]
- Reisen, W.; Monath, T. Western Equine Encephalomelitis. In The Arboviruses: Epidemiology and Ecology; Monath, T., Ed.; CRC Press: Boca Raton, FL, USA, 2019; Volume 2, pp. 1–241. [Google Scholar]
- Kock, R. Vertebrate reservoirs and secondary epidemiological cycles of vector-borne diseases. Rev. Sci. Tech. 2015, 34, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Coffey, L.L.; Forrester, N.; Tsetsarkin, K.; Vasilakis, N.; Weaver, S.C. Factors shaping the adaptive landscape for arboviruses: Implications for the emergence of disease. Future Microbiol. 2013, 8, 155–176. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Martínez, Y.; Vega-Useche, L.; Villamil-Gómez, W.E.; Rodriguez-Morales, A.J. Saint Louis Encephalitis Virus, another re-emerging arbovirus: A literature review of worldwide research. Infez. Med. 2017, 25, 77–79. [Google Scholar]
- Curren, E.J.; Lindsey, N.P.; Fischer, M.; Hills, S.L. St. Louis encephalitis virus disease in the United States, 2003–2017. Am. J. Trop. Med. Hyg. 2018, 99, 1074. [Google Scholar] [CrossRef] [PubMed]
- Diaz, A.; Coffey, L.L.; Burkett-Cadena, N.; Day, J.F. Reemergence of St. Louis encephalitis virus in the Americas. Emerg. Infect. Dis. 2018, 24, 2150. [Google Scholar] [CrossRef]
- Root, J.J. West Nile virus associations in wild mammals: A synthesis. Arch. Virol. 2013, 158, 735–752. [Google Scholar] [CrossRef]
- Root, J.J.; Bosco-Lauth, A.M. West Nile virus associations in wild mammals: An update. Viruses 2019, 11, 459. [Google Scholar] [CrossRef]
- Ozkul, A.; Yildirim, Y.; Pinar, D.; Akcali, A.; Yilmaz, V.; COLAK, D. Serological evidence of West Nile Virus (WNV) in mammalian species in Turkey. Epidemiol. Infect. 2006, 134, 826–829. [Google Scholar] [CrossRef]
- Docherty, D.E.; Samuel, M.D.; Nolden, C.A.; Egstad, K.F.; Griffin, K.M. West Nile virus antibody prevalence in wild mammals, Southern Wisconsin. Emerg. Infect. Dis. 2006, 12, 1982. [Google Scholar] [CrossRef]
- Ciota, A.T.; Kramer, L.D. Vector-virus interactions and transmission dynamics of West Nile virus. Viruses 2013, 5, 3021–3047. [Google Scholar] [CrossRef] [PubMed]
- McLean, R.; Ubico, S.; Bourne, D.; Komar, N. West Nile virus in livestock and wildlife. In Japanese Encephalitis and West Nile Viruses; Springer: Berlin/Heidelberg, Germany, 2002; pp. 271–308. [Google Scholar] [CrossRef]
- Gómez, A.; Kilpatrick, A.M.; Kramer, L.D.; Dupuis, A.P. Land use and West Nile virus seroprevalence in wild mammals. Emerg. Infect. Dis. 2008, 14, 962. [Google Scholar] [CrossRef] [PubMed]
- Blitvich, B.J.; Juarez, L.I.; Tucker, B.J.; Rowley, W.A.; Platt, K.B. Antibodies to West Nile virus in raccoons and other wild peridomestic mammals in Iowa. J. Wildl. Dis. 2009, 45, 1163–1168. [Google Scholar] [CrossRef] [PubMed]
- Almeida, A.; Galão, R.; Sousa, C.; Novo, M.; Parreira, R.; Pinto, J.; Piedade, J.; Esteves, A. Potential mosquito vectors of arboviruses in Portugal: Species, distribution, abundance and West Nile infection. Trans. R. Soc. Trop. Med. Hyg. 2008, 102, 823–832. [Google Scholar] [CrossRef]
- Napp, S.; Petrić, D.; Busquets, N. West Nile virus and other mosquito-borne viruses present in Eastern Europe. Pathog. Glob. Health 2018, 112, 233–248. [Google Scholar] [CrossRef]
- Charrel, R.N.; de Lamballerie, X. West Nile virus, an emerging arbovirus. Presse Med. 2004, 33, 1521–1526. [Google Scholar] [CrossRef]
- Taylor, R.; Work, T.; Hurlbut, H.; Rizk, F. A study of the ecology of West Nile Virus in Egypt1. Am. J. Trop. Med. Hyg. 1956, 5, 579–620. [Google Scholar] [CrossRef]
- Tendero, D.T. Capitulo 19: Enfermedades víricas transmitidas por artrópodos (enfermedades por arbovirus). In Medicina Humanitaria; Estebanes, P., Ed.; Ediciones Díaz de Santos: Madrid, Spain, 2005; p. 858. [Google Scholar]
- Gurukumar, K.; Priyadarshini, D.; Patil, J.; Bhagat, A.; Singh, A.; Shah, P.; Cecilia, D. Development of real time PCR for detection and quantitation of dengue viruses. Virol. J. 2009, 6, 10. [Google Scholar] [CrossRef]
- Pinheiro, F.d.P.; Bensabath, G. Vigilancia e investigacion de las enfermedades infecciosas a lo largo de la carretera transamazonica del Brasil. Boletín Oficina Sanit. Panam. 1974, 77, 187–198. [Google Scholar]
- Suwanmanee, S.; Surasombatpattana, P.; Soonthornworasiri, N.; Hamel, R.; Maneekan, P.; Missé, D.; Luplertlop, N. Monitoring arbovirus in Thailand: Surveillance of dengue, chikungunya and zika virus, with a focus on coinfections. Acta Trop. 2018, 188, 244–250. [Google Scholar] [CrossRef]
- De Thoisy, B.; Matheus, S.; Catzeflis, F.; Clément, L.; Barrioz, S.; Guidez, A.; Donato, D.; Cornu, J.F.; Brunaux, O.; Guitet, S.; et al. Maripa hantavirus in French Guiana: Phylogenetic position and predicted spatial distribution of rodent hosts. Am. J. Trop. Med. Hyg. 2014, 90, 988–992. [Google Scholar] [CrossRef] [PubMed]
- Andersen, K.G.; Rambaut, A.; Lipkin, W.I.; Holmes, E.C.; Garry, R.F. The proximal origin of SARS-CoV-2. Nat. Med. 2020, 26, 450–452. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Wu, C.; Li, X.; Song, Y.; Yao, X.; Wu, X.; Duan, Y.; Zhang, H.; Wang, Y.; Qian, Z. On the origin and continuing evolution of SARS-CoV-2. Natl. Sci. Rev. 2020, 7, 1012–1023. [Google Scholar] [CrossRef] [PubMed]
- Espinal, M.A.; Andrus, J.K.; Jauregui, B.; Hull Waterman, S.; Morens, D.M.; Santos, J.I.; Horstick, O.; Francis, L.A.; Olson, D. Arbovirosis emergentes y reemergentes transmitidas por Aedes en la Región de las Américas: Implicaciones en materia de políticas de salud. Rev. Panam. Salud Publica 2019, 43, 1–8. [Google Scholar] [CrossRef]
- Mendenhall, I.H.; Manuel, M.; Moorthy, M.; Lee, T.T.; Low, D.H.; Missé, D.; Gubler, D.J.; Ellis, B.R.; Ooi, E.E.; Pompon, J. Peridomestic Aedes malayensis and Aedes albopictus are capable vectors of arboviruses in cities. PLoS Negl. Trop. Dis. 2017, 11, e0005667. [Google Scholar] [CrossRef]
- Fraiture, M.-A.; Coucke, W.; Pol, M.; Rousset, D.; Gourinat, A.-C.; Biron, A.; Broeders, S.; Vandermassen, E.; Dupont-Rouzeyrol, M.; Roosens, N.H. Non-Invasive versus Invasive Samples for Zika Virus Surveillance: A Comparative Study in New Caledonia and French Guiana in 2015–2016. Microorganisms 2021, 9, 1312. [Google Scholar] [CrossRef]
- dos Santos Fonseca, L.M.; Carvalho, R.H.; Bandeira, A.C.; Sardi, S.I.; Campos, G.S. Oropouche Virus Detection in Febrile Patients’ Saliva and Urine Samples in Salvador, Bahia, Brazil. Jpn. J. Infect. Dis. 2020, 73, 164–165. [Google Scholar] [CrossRef]
- Musso, D.; Teissier, A.; Rouault, E.; Teururai, S.; de Pina, J.-J.; Nhan, T.-X. Detection of chikungunya virus in saliva and urine. Virol. J. 2016, 13, 102. [Google Scholar] [CrossRef]
- Milich, K.M.; Koestler, B.J.; Simmons, J.H.; Nehete, P.N.; Di Fiore, A.; Williams, L.E.; Dudley, J.P.; Vanchiere, J.; Payne, S.M. Methods for detecting Zika virus in feces: A case study in captive squirrel monkeys (Saimiri boliviensis boliviensis). PLoS ONE 2018, 13, e0209391. [Google Scholar] [CrossRef]
- Taberlet, P.; Luikart, G. Non-invasive genetic sampling and individual identification. Biol. J. Linn. Soc. 1999, 68, 41–55. [Google Scholar] [CrossRef]
- Morin, P.; Woodruff, D. Noninvasive genotyping for vertebrate. In Molecular Genetic Approaches in Conservation; Oxford University Press: New York, NY, USA, 1996; pp. 298–313. [Google Scholar]
- Carrillo-Bilbao, G.; Martin-Solano, S.; Saegerman, C. Zoonotic Blood-Borne Pathogens in Non-Human Primates in the Neotropical Region: A Systematic Review. Pathogens 2021, 10, 1009. [Google Scholar] [CrossRef] [PubMed]
- West, K.A.; Heymann, E.W.; Mueller, B.; Gillespie, T.R. Patterns of Infection with Cryptosporidium sp. and Giardia sp. in Three Species of Free-Ranging Primates in the Peruvian Amazon. Int. J. Primatol. 2013, 34, 939–945. [Google Scholar] [CrossRef]
- Mlinarić, A.; Horvat, M.; Šupak Smolčić, V. Dealing with the positive publication bias: Why you should really publish your negative results. Biochem. Med. 2017, 27, 030201. [Google Scholar] [CrossRef] [PubMed]
- Matosin, N.; Frank, E.; Engel, M.; Lum, J.S.; Newell, K.A. Negativity towards negative results: A discussion of the disconnect between scientific worth and scientific culture. Dis. Model Mech. 2014, 7, 171–173. [Google Scholar] [CrossRef]
- Carrillo-Bilbao, G.; Navarro, J.-C.; Martin-Solano, S.; Chávez-Larrea, M.-A.; Cholota-Iza, C.; Saegerman, C. First Molecular Identification of Trypanosomes and Absence of Babesia sp. DNA in Faeces of Non-Human Primates in the Ecuadorian Amazon. Pathogens 2022, 11, 1490. [Google Scholar] [CrossRef]
- Albanna, B.; Handl, J.; Heeks, R. Publication outperformance among global South researchers: An analysis of individual-level and publication-level predictors of positive deviance. Scientometrics 2021, 126, 8375–8431. [Google Scholar] [CrossRef]
- Shuman, E.K. Global climate change and infectious diseases. N. Engl. J. Med. 2010, 362, 1061–1063. [Google Scholar] [CrossRef]
- Chen, M.-J.; Lin, C.-Y.; Wu, Y.-T.; Wu, P.-C.; Lung, S.-C.; Su, H.-J. Effects of extreme precipitation to the distribution of infectious diseases in Taiwan, 1994–2008. PLoS ONE 2012, 7, e34651. [Google Scholar] [CrossRef]
- Morin, C.W.; Comrie, A.C. Regional and seasonal response of a West Nile virus vector to climate change. Proc. Natl. Acad. Sci. USA 2013, 110, 15620–15625. [Google Scholar] [CrossRef]
- Liang, L.; Gong, P. Climate change and human infectious diseases: A synthesis of research findings from global and spatio-temporal perspectives. Environ. Int. 2017, 103, 99–108. [Google Scholar] [CrossRef]
- Jones, L.; Davies, C.; Steele, G.; Nuttall, P. A novel mode of arbovirus transmission involving a nonviremic host. Science 1987, 237, 775–777. [Google Scholar] [CrossRef] [PubMed]
- Forshey, B.M.; Guevara, C.; Laguna-Torres, V.A.; Cespedes, M.; Vargas, J.; Gianella, A.; Vallejo, E.; Madrid, C.; Aguayo, N.; Gotuzzo, E. Arboviral etiologies of acute febrile illnesses in Western South America, 2000–2007. PLoS Negl. Trop. Dis. 2010, 4, e787. [Google Scholar] [CrossRef] [PubMed]
- Lwande, O.W.; Obanda, V.; Lindström, A.; Ahlm, C.; Evander, M.; Näslund, J.; Bucht, G. Globe-trotting Aedes aegypti and Aedes albopictus: Risk factors for arbovirus pandemics. Vector-Borne Zoonotic Dis. 2020, 20, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Couto-Lima, D.; Andreazzi, C.S.; Leite, P.J.; Bersot, M.I.L.; Alencar, J.; Lourenço-de-Oliveira, R. Seasonal population dynamics of the primary yellow fever vector Haemagogus leucocelaenus (Dyar & Shannon)(Diptera: Culicidae) is mainly influenced by temperature in the Atlantic Forest, southeast Brazil. Mem. Inst. Oswaldo Cruz 2020, 115. [Google Scholar] [CrossRef]
- González-Salazar, C.; Stephens, C.R.; Sánchez-Cordero, V. Predicting the potential role of non-human hosts in Zika virus maintenance. EcoHealth 2017, 14, 171–177. [Google Scholar] [CrossRef]
- Reeves, W.C. Mosquito vector and vertebrate host interaction: The key to maintenance of certain arboviruses. In Ecology and Physiology of Parasites; University of Toronto Press: Toronto, ON, Canada, 2019; pp. 223–231. [Google Scholar]
- Coffey, L.L.; Vasilakis, N.; Brault, A.C.; Powers, A.M.; Tripet, F.; Weaver, S.C. Arbovirus evolution in vivo is constrained by host alternation. Proc. Natl. Acad. Sci. USA 2008, 105, 6970–6975. [Google Scholar] [CrossRef]
- Peterson, A.T. Ecological niche modelling and understanding the geography of disease transmission. Vet. Ital. 2007, 43, 393–400. [Google Scholar]
- Barnosky, A.D.; Hadly, E.A.; Bell, C.J. Mammalian Response to Global Warming on Varied Temporal Scales. J. Mammal. 2003, 84, 354–368. [Google Scholar] [CrossRef]
- Jones, R.; Kulkarni, M.A.; Davidson, T.M.; Team, R.-L.R.; Talbot, B. Arbovirus vectors of epidemiological concern in the Americas: A scoping review of entomological studies on Zika, dengue and chikungunya virus vectors. PLoS ONE 2020, 15, e0220753. [Google Scholar] [CrossRef]
- Bittar, C.; Machado, R.R.; Comelis, M.T.; Bueno, L.M.; Morielle-Versute, E.; Beguelini, M.R.; De Souza, R.P.; Nogueira, M.L.; Rahal, P. Lack of serological and molecular evidence of arbovirus infections in bats from Brazil. PLoS ONE 2018, 13, e0207010. [Google Scholar] [CrossRef]
- Navarro, J.-C.; Carrera, J.-P.; Liria, J.; Auguste, A.J.; Weaver, S.C. Alphaviruses in Latin America and the Introduction of Chikungunya Virus. In Human Virology in Latin America: From Biology to Control; Ludert, J.E., Pujol, F.H., Arbiza, J., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 169–192. [Google Scholar]
- Tongthainan, D.; Mongkol, N.; Jiamsomboon, K.; Suthisawat, S.; Sanyathitiseree, P.; Sukmak, M.; Wajjwalku, W.; Poovorawan, Y.; Ieamsaard, G.; Sangkharak, B. Seroprevalence of Dengue, Zika, and Chikungunya Viruses in Wild Monkeys in Thailand. Am. J. Trop. Med. Hyg. 2020, 103, 1228. [Google Scholar] [CrossRef] [PubMed]
- Vourc’h, G.; Halos, L.; Desvars, A.; Boué, F.; Pascal, M.; Lecollinet, S.; Zientara, S.; Duval, T.; Nzonza, A.; Brémont, M. Chikungunya antibodies detected in non-human primates and rats in three Indian Ocean islands after the 2006 ChikV outbreak. Vet. Res. 2014, 45, 52. [Google Scholar] [CrossRef] [PubMed]
- Halstead, S.B.; Udomsakdi, S. Vertebrate hosts of chikungunya virus. Bull. World Health Organ. 1966, 35, 89. [Google Scholar] [PubMed]
- Mascarenhas, M.; Garasia, S.; Berthiaume, P.; Corrin, T.; Greig, J.; Ng, V.; Young, I.; Waddell, L. A scoping review of published literature on chikungunya virus. PLoS ONE 2018, 13, e0207554. [Google Scholar] [CrossRef]
- Gardner, J.; Rudd, P.A.; Prow, N.A.; Belarbi, E.; Roques, P.; Larcher, T.; Gresh, L.; Balmaseda, A.; Harris, E.; Schroder, W.A. Infectious chikungunya virus in the saliva of mice, monkeys and humans. PLoS ONE 2015, 10, e0139481. [Google Scholar] [CrossRef] [PubMed]
- Opayele Adewale, V.; Adeniji Johnson, A.; Ibrahim Kolade, T.; Olaleye Olufemi, D. Influence of meteorological variables on diversity and abundance of mosquito vectors in two livestock farms in Ibadan, Nigeria: Public health implications. J. Mosq. Res. 2017, 7, 70. [Google Scholar] [CrossRef]
- Chaves, B.A.; Orfano, A.; Nogueira, P.; Rodrigues, N.; Campolina, T.; Nacif-Pimenta, R.; Pires, A.; Júnior, A.; Paz, A.; Vaz, E.; et al. Coinfection with Zika Virus (ZIKV) and Dengue Virus Results in Preferential ZIKV Transmission by Vector Bite to Vertebrate Host. J. Infect. Dis. 2018, 218, 563–571. [Google Scholar] [CrossRef]
- Salazar-Maya, S.; Tascon-Terranova, V.; Palacio-Holguín, S.; Vélez-Quintero, D.; Ocampo-Betancur, M.; Ulloa-Zuluaga, E.; Bonilla-Aldana, D.K.; Rodríguez-Morales, A.J. Principales enfermedades infecciosas y zoonóticas en el Equus caballus y su estado actual en el trópico colombiano. Rev. Panam. Enferm. Infecc. 2018, 1, 98–101. [Google Scholar]
- Brown, J.E.; Evans, B.R.; Zheng, W.; Obas, V.; Barrera-Martinez, L.; Egizi, A.; Zhao, H.; Caccone, A.; Powell, J.R. Human impacts have shaped historical and recent evolution in Aedes aegypti, the dengue and yellow fever mosquito. Evolution 2014, 68, 514–525. [Google Scholar] [CrossRef]
- Caminade, C.; McIntyre, K.M.; Jones, A.E. Impact of recent and future climate change on vector-borne diseases. Ann. N. Y. Acad. Sci. 2019, 1436, 157. [Google Scholar] [CrossRef]
- Rogers, D.; Randolph, S. Climate change and vector-borne diseases. Adv. Parasitol. 2006, 62, 345–381. [Google Scholar] [CrossRef] [PubMed]
- Gottwalt, A. Impacts of deforestation on vector-borne disease incidence. Columbia Univ. J. Glob. Health 2013, 3, 16–19. [Google Scholar] [CrossRef]
- Lines, J.; Harpham, T.; Leake, C.; Schofield, C. Trends, priorities and policy directions in the control of vector-borne diseases in urban environments. Health Policy Plan. 1994, 9, 113–129. [Google Scholar] [CrossRef] [PubMed]
- Hunter, P.R. Climate change and waterborne and vector-borne disease. J. Appl. Microbiol. 2003, 94, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Donalisio, M.R.; Freitas, A.R.R.; Zuben, A.P.B.V. Arboviruses emerging in Brazil: Challenges for clinic and implications for public health. Rev. Saude Publica 2017, 51, 30. [Google Scholar] [CrossRef]
- Vasconcelos, P.F.; Travassos da Rosa, A.; Rodrigues, S.G.; Travassos da Rosa, E.S.; Dégallier, N.; Travassos da Rosa, J.F. Inadequate management of natural ecosystem in the Brazilian Amazon region results in the emergence and reemergence of arboviruses. Cad. Saude Publica 2001, 17, S155–S164. [Google Scholar] [CrossRef]
- Foley, J.A.; Asner, G.P.; Costa, M.H.; Coe, M.T.; DeFries, R.; Gibbs, H.K.; Howard, E.A.; Olson, S.; Patz, J.; Ramankutty, N. Amazonia revealed: Forest degradation and loss of ecosystem goods and services in the Amazon Basin. Front. Ecol. Environ. 2007, 5, 25–32. [Google Scholar] [CrossRef]
- Davies, T.J.; Purvis, A.; Gittleman, J.L. Quaternary climate change and the geographic ranges of mammals. Am. Nat. 2009, 174, 297–307. [Google Scholar] [CrossRef]
- Berberian, G.; Rosanova, M.T. Impacto del cambio climático en las enfermedades infecciosas. Arch. Argent. Pediatr. 2012, 110, 39–45. [Google Scholar] [CrossRef]
- Walsh, J.; Molyneux, D.; Birley, M. Deforestation: Effects on vector-borne disease. Parasitology 1993, 106, S55–S75. [Google Scholar] [CrossRef]
- Burkett-Cadena, N.D.; Vittor, A.Y. Deforestation and vector-borne disease: Forest conversion favors important mosquito vectors of human pathogens. Basic Appl. Ecol. 2018, 26, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Musso, D.; Rodriguez-Morales, A.J.; Levi, J.E.; Cao-Lormeau, V.-M.; Gubler, D.J. Unexpected outbreaks of arbovirus infections: Lessons learned from the Pacific and tropical America. Lancet Infect. Dis. 2018, 18, e355–e361. [Google Scholar] [CrossRef]
- Altizer, S.; Ostfeld, R.S.; Johnson, P.T.; Kutz, S.; Harvell, C.D. Climate change and infectious diseases: From evidence to a predictive framework. Science 2013, 341, 514–519. [Google Scholar] [CrossRef]
- Cardinale, B.J.; Duffy, J.E.; Gonzalez, A.; Hooper, D.U.; Perrings, C.; Venail, P.; Narwani, A.; Mace, G.M.; Tilman, D.; Wardle, D.A. Biodiversity loss and its impact on humanity. Nature 2012, 486, 59–67. [Google Scholar] [CrossRef]
- Walther, G.-R.; Post, E.; Convey, P.; Menzel, A.; Parmesan, C.; Beebee, T.J.; Fromentin, J.-M.; Hoegh-Guldberg, O.; Bairlein, F. Ecological responses to recent climate change. Nature 2002, 416, 389–395. [Google Scholar] [CrossRef]
- Coluchi, N. Neuroinvasividad, neurotropismo y neurovirulencia en las infecciones por Arbovirus. Un desafío presente. Discov. Med. 2019, 3, 9–12. [Google Scholar]
- Watts, N.; Adger, W.N.; Agnolucci, P.; Blackstock, J.; Byass, P.; Cai, W.; Chaytor, S.; Colbourn, T.; Collins, M.; Cooper, A. Health and climate change: Policy responses to protect public health. Lancet 2015, 386, 1861–1914. [Google Scholar] [CrossRef] [PubMed]
- Amela Heras, C.; Sierra Moros, M.J. Enfermedades transmitidas por vectores: Un nuevo reto para los sistemas de vigilancia y la salud pública. Gac. Sanit. 2016, 30, 167–169. [Google Scholar] [CrossRef] [PubMed]
- Achee, N.L.; Grieco, J.P.; Vatandoost, H.; Seixas, G.; Pinto, J.; Ching-Ng, L.; Martins, A.J.; Juntarajumnong, W.; Corbel, V.; Gouagna, C. Alternative strategies for mosquito-borne arbovirus control. PLoS Negl. Trop. Dis. 2019, 13, e0006822. [Google Scholar] [CrossRef]
- Stephen, C.; Sleeman, J.; Nguyen, N.; Zimmer, P.; Duff, J.; Gavier-Widén, D.; Grillo, T.; Lee, H.; Rijks, J.; Ryser-Degiorgis, M.-P. Proposed attributes of national wildlife health programmes. Rev. Sci. Tech.-Off. Int. Des Epizoot. 2018, 37. [Google Scholar] [CrossRef]
- Spiegel, J.; Bennett, S.; Hattersley, L.; Hayden, M.H.; Kittayapong, P.; Nalim, S.; Wang, D.N.C.; Zielinski-Gutiérrez, E.; Gubler, D. Barriers and bridges to prevention and control of dengue: The need for a social–ecological approach. EcoHealth 2005, 2, 273–290. [Google Scholar] [CrossRef]
- Girard, M.; Nelson, C.B.; Picot, V.; Gubler, D.J. Arboviruses: A global public health threat. Vaccine 2020, 38, 3989–3994. [Google Scholar] [CrossRef] [PubMed]
- Esser, H.J.; Mögling, R.; Cleton, N.B.; Van Der Jeugd, H.; Sprong, H.; Stroo, A.; Koopmans, M.P.; De Boer, W.F.; Reusken, C.B. Risk factors associated with sustained circulation of six zoonotic arboviruses: A systematic review for selection of surveillance sites in non-endemic areas. Parasites Vectors 2019, 12, 265. [Google Scholar] [CrossRef]
- Wu, X.; Lu, Y.; Zhou, S.; Chen, L.; Xu, B. Impact of climate change on human infectious diseases: Empirical evidence and human adaptation. Environ. Int. 2016, 86, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Paixão, E.S.; Teixeira, M.G.; Rodrigues, L.C. Zika, chikungunya and dengue: The causes and threats of new and re-emerging arboviral diseases. BMJ Glob. Health 2018, 3, e000530. [Google Scholar] [CrossRef]
- Tami, A.; Grillet, M.E.; Grobusch, M.P. Applying geographical information systems (GIS) to arboviral disease surveillance and control: A powerful tool. Travel Med. Infect. Dis. 2016, 14, 9–10. [Google Scholar] [CrossRef] [PubMed]
- Wilke, A.B.B.; Gomes, A.d.C.; Natal, D.; Marrelli, M.T. Controle de vectores utilizando mosquitos geneticamente modificados. Rev. Saude Publica 2009, 43, 869–874. [Google Scholar] [CrossRef]
- Dusfour, I.; Vontas, J.; David, J.-P.; Weetman, D.; Fonseca, D.M.; Corbel, V.; Raghavendra, K.; Coulibaly, M.B.; Martins, A.J.; Kasai, S. Management of insecticide resistance in the major Aedes vectors of arboviruses: Advances and challenges. PLoS Negl. Trop. Dis. 2019, 13, e0007615. [Google Scholar] [CrossRef]
- Jansen, C.C.; Beebe, N.W. The dengue vector Aedes aegypti: What comes next. Microbes Infect. 2010, 12, 272–279. [Google Scholar] [CrossRef]
- Soper, F.L. El Aedes Aegypti y la fiebre amarilla. Boletín Oficina Sanit. Panam. 1968, 64, 187–196. [Google Scholar]
- Montero, A. Fiebre chikungunya-Una nueva amenaza global. Med. Clín. 2015, 145, 118–123. [Google Scholar] [CrossRef] [PubMed]
- Cucunubá, Z.M. De la epidemia de Zika en Latinoamérica y la toma de decisiones bajo incertidumbre. Rev. Univ. Ind. Santander. Salud 2016, 48, 158–160. [Google Scholar]
- Araújo, P.A.; Freitas, M.O.; Chiang, J.O.; Silva, F.A.; Chagas, L.L.; Casseb, S.M.; Silva, S.P.; Nunes-Neto, J.P.; Rosa-Júnior, J.W.; Nascimento, B.S. Investigation about the occurrence of transmission cycles of arbovirus in the tropical forest, amazon region. Viruses 2019, 11, 774. [Google Scholar] [CrossRef] [PubMed]
- Romero-Vázquez, A.; Martínez-Hernández, C.M.; Cano-González, S.B.; Jiménez-Sastre, A. Impacto de las acciones de promoción y control en la prevención del dengue. Salud Tabasco 2005, 11, 380–386. [Google Scholar]
- Llanos, M.; Carbajal, E.I.V.; Agui, M.O. Communicative strategy aimed at reducing exposure to arbovirus risk factors. MediSur 2020, 18, 161–170. [Google Scholar]
- Russell, R. Mosquito-borne arboviruses in Australia: The current scene and implications of climate change for human health. Int. J. Parasitol. 1998, 28, 955–969. [Google Scholar] [CrossRef]
- Rodríguez-Morales, A.J.; Paniz-Mondolfi, A.E.; Villamil-Gómez, W.E.; Navarro, J.C. Mayaro, Oropouche and Venezuelan equine encephalitis viruses: Following in the footsteps of Zika? Travel Med. Infect. Dis. 2017, 15, 72–73. [Google Scholar] [CrossRef]
- Muñoz, M.; Navarro, J.C. Virus Mayaro: Un arbovirus reemergente en Venezuela y Latinoamérica. Biomédica 2012, 32, 286–302. [Google Scholar] [CrossRef]
- Maia, L.; Bezerra, M.; Costa, M.; Souza, E.; Oliveira, M.; Ribeiro, A.; Miyazaki, R.; Slhessarenko, R. Natural vertical infection by dengue virus serotype 4, Zika virus and Mayaro virus in Aedes (Stegomyia) aegypti and Aedes (Stegomyia) albopictus. Med. Vet. Entomol. 2019, 33, 437–442. [Google Scholar] [CrossRef]
- Wiggins, K.; Eastmond, B.; Alto, B.W. Transmission potential of Mayaro virus in Florida Aedes aegypti and Aedes albopictus mosquitoes. Med. Vet. Entomol. 2018, 32, 436–442. [Google Scholar] [CrossRef]
- Pereira, T.N.; Carvalho, F.D.; De Mendonça, S.F.; Rocha, M.N.; Moreira, L.A. Vector competence of Aedes aegypti, Aedes albopictus, and Culex quinquefasciatus mosquitoes for Mayaro virus. PLoS Negl. Trop. Dis. 2020, 14, e0007518. [Google Scholar] [CrossRef] [PubMed]
- Antonelli, A.; Zizka, A.; Carvalho, F.A.; Scharn, R.; Bacon, C.D.; Silvestro, D.; Condamine, F.L. Amazonia is the primary source of Neotropical biodiversity. Proc. Natl. Acad. Sci. USA 2018, 115, 6034–6039. [Google Scholar] [CrossRef] [PubMed]
- Lavergne, A.; Lacoste, V.; Germain, A.; Matheus, S.; Dussart, P.; Deparis, X.; de Thoisy, B. Dengue virus infection in neotropical forest mammals: Incidental hosts or potential reservoirs? Med. Trop. Rev. Corps Sante Colon. 2009, 69, 345–350. [Google Scholar]










| Arbovirus | Mammalian Host | Country | Number | Prevalence (%) | Methods | Type of Sample | Reference |
|---|---|---|---|---|---|---|---|
| ALPHA: alphavirus undetermined | Perissodactyla | Brazil | 985 | 24.26 | PRNT | Blood | [98] |
| Apeu virus (APEUV) | Artyodactyla | Brazil | 607 | 82.2 | PRNT | Serum | [99] |
| Bunyamwera virus (BUNV) | Perissodactyla Didelphimorphia | Argentina Venezuela | 2 | 100 | PRC qPCR | Tissue | [100,101] |
| Bussuquara virus (BSQV) | Perissodactyla Non-human primates | Argentina Brazil Mexico | 108 72 | 77.33 64.81 | PRNT | Serum | [102,103,104] |
| Cache Valley virus (CVV) | Carnívora Rodentia | Mexico Panama | 2214 80 | 1.25 1.26 | HI | Blood Serum | [105,106] |
| Cacipacore virus (CPCV) | Non-human primates | Brazil | 139 | 17.98 | HI PRNT | Serum | [102] |
| Caraparu virus (CARV) | Non-human primates | Brazil | 139 | 17.98 | HI PRNT | Serum | [102] |
| Catu virus (CATUV) | Didelphimorphia | Brazil | 14 | 57.14 | HI | Serum | [67] |
| Changuinola virus (CGLV) | Pilosa Non-human primates Rodentia | Colombia Costa Rica Panama | 2214 4109 | 1.26 79.81 | HI | Blood Serum | [106,107] |
| Dengue virus (DENV) | Carnivora Chiroptera Didelphimorphia Non-human primates Rodentia | Argentina Brazil Colombia Costa Rica French Guiana Mexico | 616 16 | 64.81 0.69 | ELISA HI PCR PRNT qPCR | Blood Serum Tissue | [102,103,108,109,110,111,112,113,114,115,116,117,118] |
| Madariaga virus (MADV) | Chiroptera Perissodactyla Non-human primates Rodentia | Brazil Guatemala Mexico Venezuela | 2214 14 | 93.75 1.26 | HI PRNT | Blood Serum Tissue | [67,98,102,104,119,120,121,122] |
| Western equine encephalitis virus (WEEV) | Perissodactyla Rodentia | Brazil Mexico | 985 75 | 77.33 1.25 | HI PRNT | Blood Serum | [98,105] |
| Saint Louis encephalitis virus (SLEV) | Artiodactyla Chiroptera Didelphimorphia Perissodactyla Pilosa Non-human primates Rodentia | Argentina Brazil Colombia Costa Rica Guatemala French Guiana Mexico Panama Uruguay | 2214 1 | 100 1.25 | ELISA HI PRNT qPCR | Blood Serum | [102,103,104,105,106,107,110,111,112,114,116,120,123,124,125,126,127,128,129,130,131] |
| Venezuelan equine encephalitis virus (VEEV) | Artiodactyla Chiroptera Didelphimorphia Perissodactyla Pilosa Rodentia | Argentina Brazil Colombia Costa Rica Guatemala Mexico Panama Venezuela | 985 64 | 93.75 0.69 | HI PRNT qPCR | Blood Serum Tissue | [98,101,104,105,106,107,120,121,122,123,126,129,132,133] |
| Vesicular stomatitis, Indiana virus (VSIV) | Chiroptera Didelphimorphia Pilosa Non-human primates Rodentia | Colombia Costa Rica Guatemala Mexico Panama | 2214 80 | 79.81 1.25 | HI PRNT | Blood Serum | [105,106,107,120] |
| Vesicular stomatitis, New Jersey virus (VSNJV) | Chiroptera Rodentia | Guatemala Mexico | 332 80 | 26.2 1.25 | HI PRNT | Blood Serum | [105,120] |
| Yellow fever virus (YFV) | Pilosa Non-human primates Rodentia | Brazil French Guiana | 574 1 | 100 8.71 | HI PRNT qPCR | Blood Serum Tissue | [102,112,114,128,134] |
| Undetermined flavavirus | Non-human primates | Costa Rica | 86 | 40.69 44.18 | PRNT | Serum | [110,111] |
| Fort Sherman virus (FSV) | Perissodactyla Non-human primates | Argentina Brazil | 222 50 | 76.8 1.04 | HI PRNT qPCR | Blood Serum | [123,135,136] |
| Gamboa virus (GAMV) | Rodentia | Venezuela | 543 | 5.52 | qPCR HI | Blood Tissue | [101] |
| Guama virus (GMAV) | Rodentia | Colombia Panama | 2214 | 1.26 | HI | Serum | [106] |
| Guaroa virus (GROV) | Artiodactyla | Brazil | 607 | 82.2 | PRNT | Serum | [99] |
| Icoaraci virus (ICOV) | Perissodactyla Non-human primates | Argentina Brazil | 222 26 | 76.8 17.98 | HI PRNT qPCR | Blood Serum | [123,124] |
| Ilheus virus (ILHV) | Pilosa Non-human primates | Argentina Brazil Costa Rica | 139 14 | 79.81 17.98 | HI PRNT qPCR | Blood Serum Tissue | [67,102,103,107,124] |
| Itaqui virus (ITQV) | Perissodactyla | Venezuela | 64 | 93.75 | HI | Serum Tissue | [122] |
| Jamestown Canyon virus (JCV) | Artiodactyla | Mexico | 80 | 1.25 | HI | Blood | [105] |
| Madrid virus (MADV) | Perissodactyla Rodentia | Colombia Panama | 2214 194 | 64.94 1.26 | HI | Serum | [106,126] |
| Maguari virus (MAGV) | Artiodactyla Perissodactyla | Brazil | 607 | 82.2 | PRNT | Serum | [99] |
| Mayaro virus (MAYV) | Perissodactyla Non-human primates | Argentina Brazil French Guiana Panama Paraguay | 2214 90 | 74.44 1.26 | HI PRNT | Blood Serum | [98,106,112,137] |
| Melao virus (MELV) | Lagomorpha Rodentia | Colombia Panama | 2214 | 1.26 | HI | Serum | [106] |
| Mojui dos Campos virus (MDCV) | Chiroptera | Brazil | 1 | 100 | HISTOL | Blood | [138] |
| Murutucu virus (MURV) | Artiodactyla | Brazil | 607 | 82.2 | PRNT | Serum | [99] |
| Oriboca virus (ORIV) | Artiodactyla | Brazil | 607 | 82.2 | PRNT | Serum | [99] |
| Oropouche virus (OROV) | Artiodactyla Non-human primates | Brazil Venezuela | 607 1 | 100 4 | HI PCR PRNT | Blood Serum Tissue | [99,128,136,139] |
| Ossa virus (OSSAV) | Rodentia | Colombia Panama | 2214 | 1.26 | HI | Serum | [106] |
| Punta Toro virus (PTV) | Pilosa | Costa Rica | 109 | 79.81 | HI | Blood | [107] |
| Rio Grande virus (RGV) | Lagomorpha Pilosa | Costa Rica Mexico | 109 80 | 79.81 1.25 | HI | Blood | [105,107] |
| Rocio virus (ROCV) | Perissodactyla Non-human primates | Brazil | 753 139 | 55.11 17.98 | ELISA HI PRNT | Serum Tissue | [102,130] |
| Tacaiuma virus (TCMV) | Artiodactyla Perissodactyla Pilosa | Argentina Brazil | 222 75 | 77.33 17.98 | HI PRNT | Blood Serum | [102,123,129] |
| UNA virus (UNAV) | Non-human primates Rodentia | Argentina Colombia Panama Paraguay | 2214 90 | 74.44 1.26 | HI PRNT | Blood Serum | [106,137] |
| Utinga virus (UTIV) | Non-human primates | Brazil | 139 | 17.98 | PRNT | Serum | [102] |
| West Nile virus (WNV) | Artiodactyla Chiroptera Perissodactyla Pilosa Non-human primates Rodentia | Argentina Costa Rica Mexico Panama Uruguay Venezuela | 425 72 | 79.81 18.57 | ELISA HI PRNT | Blood Serum Tissue | [103,104,107,111,115,116,121,123,125,126] |
| Xingu virus virus (XINV) | Artiodactyla Perissodactyla | Brazil | 607 | 82.2 | PRNT | Serum | [99] |
| Zika virus (ZIKV) | Pilosa Non-human primates | Brazil | 132 10 | 100 6.81 | PRNT qPCR | Blood Tissue | [140,141] |
| Argentina | Brazil | Colombia | Costa Rica | Guatemala | French Guiana | Mexico | Panama | Paraguay | Uruguay | Venezuela | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Argentina | |||||||||||
| Brazil | 45.00 | ||||||||||
| Colombia | 36.36 | 16.67 | |||||||||
| Costa Rica | 47.62 | 21.05 | 36.36 | ||||||||
| Guatemala | 25.00 | 24.24 | 37.50 | 40.00 | |||||||
| French Guiana | 37.50 | 24.24 | 25.00 | 26.67 | 20.00 | ||||||
| Mexico | 41.67 | 30.00 | 36.36 | 54.55 | 58.82 | 23.53 | |||||
| Panama | 50.00 | 26.32 | 80.00 | 43.48 | 50.00 | 25.00 | 56.00 | ||||
| Paraguay | 30.77 | 6.67 | 15.38 | 0.00 | 0.00 | 28.57 | 0.00 | 30.77 | |||
| Uruguay | 30.77 | 6.67 | 14.29 | 33.33 | 28.57 | 28.57 | 28.57 | 28.57 | 0.00 | ||
| Venezuela | 35.29 | 17.65 | 11.76 | 25.00 | 36.36 | 0.00 | 33.33 | 33.33 | 0.00 | 25.00 |
| Country | Argentina | Brazil | Colombia | Costa Rica | Guatemala | French Guiana | Mexico | Panama | Paraguay | Uruguay | Venezuela |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Argentina | |||||||||||
| Brazil | 29.03 | ||||||||||
| Colombia | 22.22 | 9.09 | |||||||||
| Costa Rica | 31.25 | 11.76 | 22.22 | ||||||||
| Guatemala | 14.29 | 13.79 | 23.08 | 25.00 | |||||||
| French Guiana | 23.08 | 13.79 | 14.29 | 15.38 | 11.11 | ||||||
| Mexico | 26.32 | 17.65 | 22.22 | 37.50 | 41.67 | 13.33 | |||||
| Panama | 33.33 | 15.15 | 66.67 | 27.78 | 33.33 | 14.29 | 38.89 | ||||
| Paraguay | 18.18 | 3.45 | 8.33 | 0.00 | 0.00 | 16.67 | 0.00 | 18.18 | |||
| Uruguay | 18.18 | 3.45 | 7.69 | 20.00 | 16.67 | 16.67 | 16.67 | 16.67 | 0.00 | ||
| Venezuela | 21.43 | 9.68 | 6.25 | 14.29 | 22.22 | 0.00 | 20.00 | 20.00 | 0.00 | 14.29 |
| Orders | Artiodactyla | Carnívora | Chiroptera | Didelphimorphia | Lagomorpha | Perissodactyla | Pilosa | Non-Human Primates | Rodentia |
|---|---|---|---|---|---|---|---|---|---|
| Artiodactyla | |||||||||
| Carnívora | 0.00 | ||||||||
| Chiroptera | 30.00 | 20.00 | |||||||
| Didelphimorphia | 22.22 | 25.00 | 61.54 | ||||||
| Lagomorpha | 0.00 | 0.00 | 0.00 | 0.00 | |||||
| Perissodactyla | 34.48 | 0.00 | 32.00 | 26.09 | 0.00 | ||||
| Pilosa | 34.78 | 0.00 | 42.11 | 35.29 | 15.38 | 28.57 | |||
| Non-human primates | 18.75 | 9.09 | 35.71 | 23.08 | 0.00 | 43.24 | 45.16 | ||
| Rodentia | 19.35 | 19.05 | 51.85 | 32.00 | 9.52 | 33.33 | 46.67 | 46.15 |
| Orders | Artiodactyla | Carnívora | Chiroptera | Didelphimorphia | Lagomorpha | Perissodactyla | Pilosa | Non-Human Primates | Rodentia |
|---|---|---|---|---|---|---|---|---|---|
| Artiodactyla | |||||||||
| Carnívora | 0.00 | ||||||||
| Chiroptera | 17.65 | 11.11 | |||||||
| Didelphimorphia | 22.22 | 14.29 | 44.44 | ||||||
| Lagomorpha | 0.00 | 0.00 | 0.00 | 0.00 | |||||
| Perissodactyla | 20.83 | 0.00 | 19.05 | 15.00 | 0.00 | ||||
| Pilosa | 21.05 | 0.00 | 26.67 | 21.43 | 8.33 | 16.67 | |||
| Non-human primates | 10.34 | 4.76 | 21.74 | 13.04 | 0.00 | 27.59 | 29.17 | ||
| Rodentia | 10.71 | 10.53 | 35.00 | 19.05 | 5.00 | 20.00 | 30.43 | 30.00 |
| Positives Mammals to Arbovirus | Negatives Mammals to Arbovirus | Total | |
|---|---|---|---|
| Mammals in deforested fronts | 1312 | 3268 | 4580 |
| Mammals in vegetation cover | 1549 | 5634 | 7183 |
| Total | 2861 | 8902 | 11,763 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Romero, C.; Carrillo Bilbao, G.A.; Navarro, J.-C.; Martin-Solano, S.; Saegerman, C. Arboviruses in Mammals in the Neotropics: A Systematic Review to Strengthen Epidemiological Monitoring Strategies and Conservation Medicine. Viruses 2023, 15, 417. https://doi.org/10.3390/v15020417
García-Romero C, Carrillo Bilbao GA, Navarro J-C, Martin-Solano S, Saegerman C. Arboviruses in Mammals in the Neotropics: A Systematic Review to Strengthen Epidemiological Monitoring Strategies and Conservation Medicine. Viruses. 2023; 15(2):417. https://doi.org/10.3390/v15020417
Chicago/Turabian StyleGarcía-Romero, Cinthya, Gabriel Alberto Carrillo Bilbao, Juan-Carlos Navarro, Sarah Martin-Solano, and Claude Saegerman. 2023. "Arboviruses in Mammals in the Neotropics: A Systematic Review to Strengthen Epidemiological Monitoring Strategies and Conservation Medicine" Viruses 15, no. 2: 417. https://doi.org/10.3390/v15020417
APA StyleGarcía-Romero, C., Carrillo Bilbao, G. A., Navarro, J.-C., Martin-Solano, S., & Saegerman, C. (2023). Arboviruses in Mammals in the Neotropics: A Systematic Review to Strengthen Epidemiological Monitoring Strategies and Conservation Medicine. Viruses, 15(2), 417. https://doi.org/10.3390/v15020417