

Virally Suppressed People Living with HIV Who Use Opioids Have Diminished Latency Reversal

Abstract
1. Introduction
2. Materials and Methods
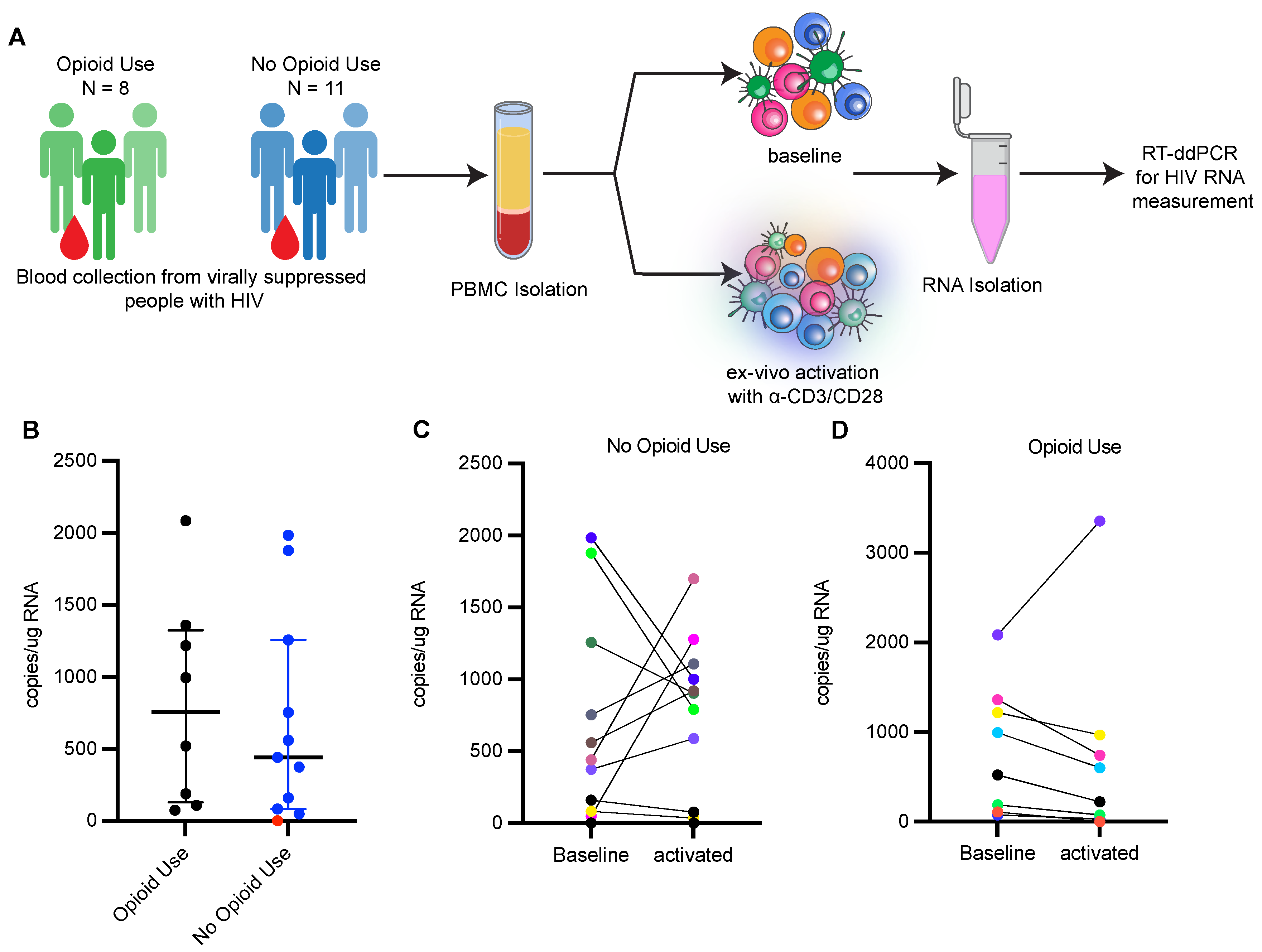
2.1. Study Design
2.2. PBMC and CD4+ T Cell Isolation for In Vitro Experiments
2.3. Packaging of Viruses for In Vitro Infections
2.4. In Vitro HIV Infections
2.5. Quantitative Real-Time Alu-PCR
2.6. RT-qPCR for Measuring RNA
2.7. HIV p24 ELISA
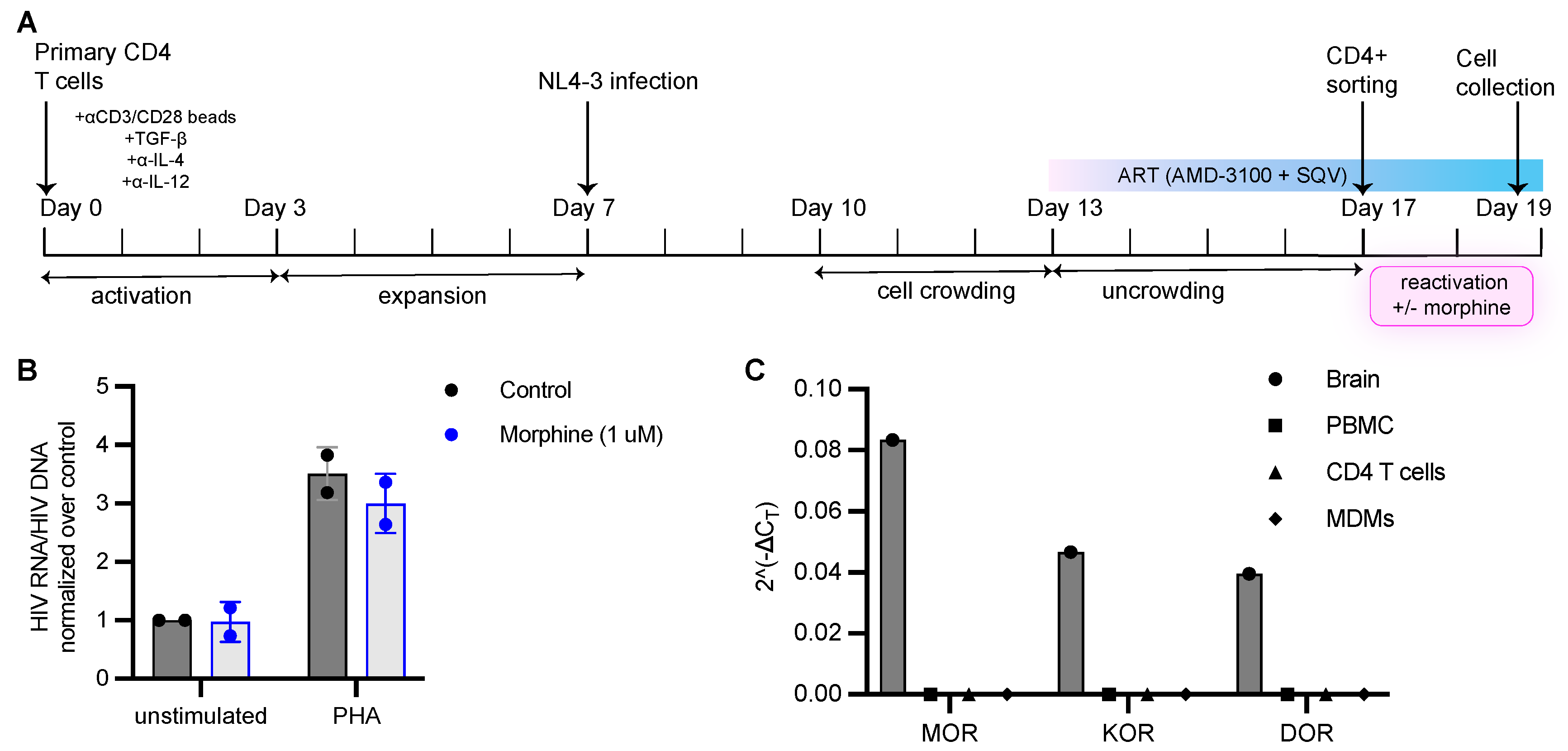
2.8. Generation of Latently Infected Cells and Reactivation
2.9. Flow Cytometry
2.10. Intact Proviral DNA Assay
2.11. Ex Vivo Activation of HIV Transcription from Clinical Samples
2.12. Measuring HIV Transcription with RT-ddPCR Assay
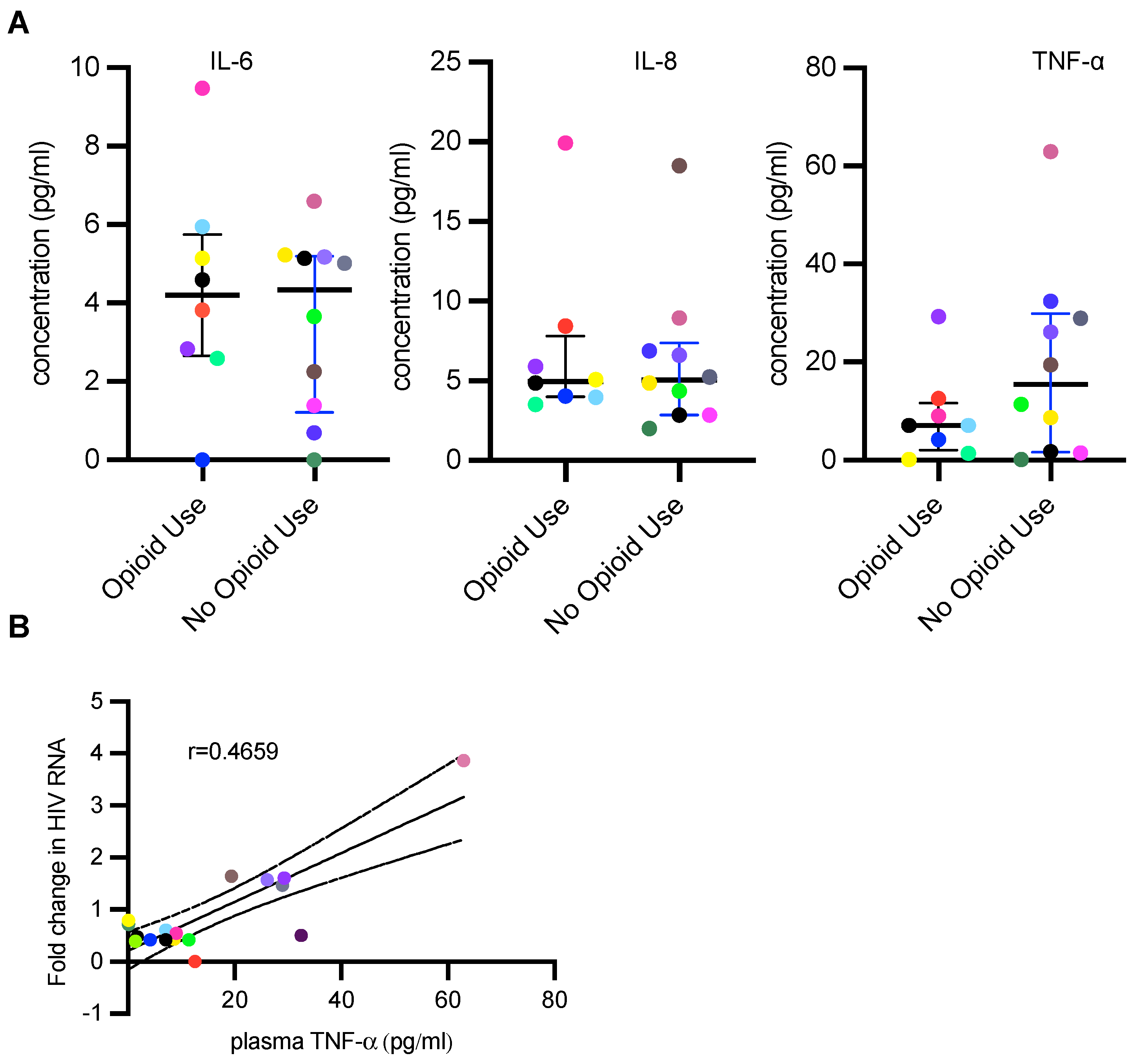
2.13. Plasma Cytokine Measurement
2.14. Statistical Analysis
2.15. Study Approval
3. Results
3.1. HIV-1 Proviruses Are Comparable between PWH Currently Using Opioids and Not Using Opioids
3.2. Reactivation of HIV-1 from Ex Vivo Stimulated PBMCs Is Limited in PWH Using Opioids
3.3. PWH Using Opioids Have Lower Plasma TNF-α Concentration
3.4. Morphine Has No Impact on HIV Infection or Establishment of Latent Infection In Vitro
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ballantyne, J.C.; LaForge, S.K. Opioid Dependence and Addiction during Opioid Treatment of Chronic Pain. PAIN 2007, 129, 235–255. [Google Scholar] [CrossRef]
- Kosten, T.R.; George, T.P. The Neurobiology of Opioid Dependence: Implications for Treatment. Sci. Pract. Perspect. 2002, 1, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Substance Abuse and Mental Health Services Administration. Key Substance Use and Mental Health Indicators in the United States: Results from the 2020 National Survey on Drug Use and Health. 2020, HHS Publication No. PEP21-07-01-003, NSDUH Series H-56. Available online: https://www.samhsa.gov/data/ (accessed on 24 July 2022).
- Products—Vital Statistics Rapid Release—Provisional Drug Overdose Data. Available online: https://www.cdc.gov/nchs/nvss/vsrr/drug-overdose-data.htm (accessed on 24 July 2022).
- Quaglio, G.; Talamini, G.; Lechi, A.; Mezzelani, P.; Lugoboni, F. Prevalence of Tuberculosis Infection and Comparison of Multiple-Puncture Liquid Tuberculin Test and Mantoux Test Among Drug Users. Scand. J. Infect. Dis. 2002, 34, 574–576. [Google Scholar] [CrossRef] [PubMed]
- MacFarlane, A.S.; Peng, X.; Meissler, J.J.; Rogers, T.J.; Geller, E.B.; Adler, M.W.; Eisenstein, T.K. Morphine Increases Susceptibility to Oral Salmonella Typhimurium Infection. J. Infect. Dis. 2000, 181, 1350–1358. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Ninkovic, J.; Banerjee, S.; Charboneau, R.G.; Das, S.; Dutta, R.; Kirchner, V.A.; Koodie, L.; Ma, J.; Meng, J.; et al. Opioid Drug Abuse and Modulation of Immune Function: Consequences in the Susceptibility to Opportunistic Infections. J. Neuroimmune Pharmacol. 2011, 6, 442. [Google Scholar] [CrossRef] [PubMed]
- Ronald, P.J.M.; Robertson, J.R.; Elton, R.A. Continued Drug Use and Other Cofactors for Progression to AIDS among Injecting Drug Users. AIDS 1994, 8, 339–344. [Google Scholar] [CrossRef]
- HIV and Injection Drug Use|HIV Transmission|HIV Basics|HIV/AIDS|CDC. Available online: https://www.cdc.gov/hiv/basics/hiv-transmission/injection-drug-use.html (accessed on 24 July 2022).
- Plein, L.M.; Rittner, H.L. Opioids and the Immune System—Friend or Foe. Br. J. Pharmacol. 2018, 175, 2717–2725. [Google Scholar] [CrossRef]
- Lucas, G.M.; Griswold, M.; Gebo, K.A.; Keruly, J.; Chaisson, R.E.; Moore, R.D. Illicit Drug Use and HIV-1 Disease Progression: A Longitudinal Study in the Era of Highly Active Antiretroviral Therapy. Am. J. Epidemiol. 2006, 163, 412–420. [Google Scholar] [CrossRef]
- Flores, J.; Liang, Y.; Ketchum, N.S.; Turner, B.J.; Bullock, D.; Villarreal, R.; Potter, J.S.; Taylor, B.S. Prescription Opioid Use Is Associated with Virologic Failure in People Living with HIV. AIDS Behav. 2018, 22, 1323–1328. [Google Scholar] [CrossRef]
- Lucas, G.M.; Cheever, L.W.; Chaisson, R.E.; Moore, R.D. Detrimental Effects of Continued Illicit Drug Use on the Treatment of HIV-1 Infection. JAIDS J. Acquir. Immune Defic. Syndr. 2001, 27, 251–259. [Google Scholar] [CrossRef]
- Lert, F.; Kazatchkine, M.D. Antiretroviral HIV Treatment and Care for Injecting Drug Users: An Evidence-Based Overview. Int. J. Drug Policy 2007, 18, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Azar, P.; Wood, E.; Nguyen, P.; Luma, M.; Montaner, J.; Kerr, T.; Milloy, M.-J. Drug Use Patterns Associated with Risk of Non-Adherence to Antiretroviral Therapy among HIV-Positive Illicit Drug Users in a Canadian Setting: A Longitudinal Analysis. BMC Infect. Dis. 2015, 15, 193. [Google Scholar] [CrossRef] [PubMed]
- Peterson, P.; Sharp, B.; Gekker, G.; Portoghese, P.; Sannerud, K.; Balfour, H. Morphine Promotes the Growth of HIV-1 in Human Peripheral Blood Mononuclear Cell Cocultures. Aids 1990, 4, 869–874. [Google Scholar] [CrossRef] [PubMed]
- Gornalusse, G.G.; Vojtech, L.N.; Levy, C.N.; Hughes, S.M.; Kim, Y.; Valdez, R.; Pandey, U.; Ochsenbauer, C.; Astronomo, R.; McElrath, J.; et al. Buprenorphine Increases HIV-1 Infection In Vitro but Does Not Reactivate HIV-1 from Latency. Viruses 2021, 13, 1472. [Google Scholar] [CrossRef] [PubMed]
- Steele, A.D.; Henderson, E.E.; Rogers, T.J. μ-Opioid Modulation of HIV-1 Coreceptor Expressionand HIV-1 Replication. Virology 2003, 309, 99–107. [Google Scholar] [CrossRef]
- Li, Y.; Wang, X.; Tian, S.; Guo, C.-J.; Douglas, S.D.; Ho, W.-Z. Methadone Enhances Human Immunodeficiency Virus Infection of Human Immune Cells. J. Infect. Dis. 2002, 185, 118–122. [Google Scholar] [CrossRef][Green Version]
- Sharp, B.M.; Gekker, G.; Li, M.D.; Chao, C.C.; Peterson, P.K. δ-Opioid Suppression of Human Immunodeficiency Virus-1 Expression in T Cells (Jurkat). Biochem. Pharmacol. 1998, 56, 289–292. [Google Scholar] [CrossRef]
- Crooks, A.M.; Bateson, R.; Cope, A.B.; Dahl, N.P.; Griggs, M.K.; Kuruc, J.D.; Gay, C.L.; Eron, J.J.; Margolis, D.M.; Bosch, R.J.; et al. Precise Quantitation of the Latent HIV-1 Reservoir: Implications for Eradication Strategies. J. Infect. Dis. 2015, 212, 1361–1365. [Google Scholar] [CrossRef]
- Finzi, D.; Blankson, J.; Siliciano, J.D.; Margolick, J.B.; Chadwick, K.; Pierson, T.; Smith, K.; Lisziewicz, J.; Lori, F.; Flexner, C.; et al. Latent Infection of CD4 + T Cells Provides a Mechanism for Lifelong Persistence of HIV-1, Even in Patients on Effective Combination Therapy. Nat. Med. 1999, 5, 512–517. [Google Scholar] [CrossRef]
- Chun, T.W.; Stuyver, L.; Mizell, S.B.; Ehler, L.A.; Mican, J.A.; Baseler, M.; Lloyd, A.L.; Nowak, M.A.; Fauci, A.S. Presence of an Inducible HIV-1 Latent Reservoir during Highly Active Antiretroviral Therapy. Proc. Natl. Acad. Sci. USA 1997, 94, 13193–13197. [Google Scholar] [CrossRef]
- Chomont, N.; El-Far, M.; Ancuta, P.; Trautmann, L.; Procopio, F.A.; Yassine-Diab, B.; Boucher, G.; Boulassel, M.-R.; Ghattas, G.; Brenchley, J.M.; et al. HIV Reservoir Size and Persistence Are Driven by T Cell Survival and Homeostatic Proliferation. Nat. Med. 2009, 15, 893–900. [Google Scholar] [CrossRef] [PubMed]
- Vandergeeten, C.; Fromentin, R.; DaFonseca, S.; Lawani, M.B.; Sereti, I.; Lederman, M.M.; Ramgopal, M.; Routy, J.-P.; Sékaly, R.-P.; Chomont, N. Interleukin-7 Promotes HIV Persistence during Antiretroviral Therapy. Blood 2013, 121, 4321–4329. [Google Scholar] [CrossRef] [PubMed]
- Maldarelli, F.; Wu, X.; Su, L.; Simonetti, F.R.; Shao, W.; Hill, S.; Spindler, J.; Ferris, A.L.; Mellors, J.W.; Kearney, M.F.; et al. Specific HIV Integration Sites Are Linked to Clonal Expansion and Persistence of Infected Cells. Science 2014, 345, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Wagner, T.A.; McLaughlin, S.; Garg, K.; Cheung, C.Y.K.; Larsen, B.B.; Styrchak, S.; Huang, H.C.; Edlefsen, P.T.; Mullins, J.I.; Frenkel, L.M. Proliferation of Cells with HIV Integrated into Cancer Genes Contributes to Persistent Infection. Science 2014, 345, 570–573. [Google Scholar] [CrossRef]
- Siliciano, J.D.; Kajdas, J.; Finzi, D.; Quinn, T.C.; Chadwick, K.; Margolick, J.B.; Kovacs, C.; Gange, S.J.; Siliciano, R.F. Long-Term Follow-up Studies Confirm the Stability of the Latent Reservoir for HIV-1 in Resting CD4 + T Cells. Nat. Med. 2003, 9, 727–728. [Google Scholar] [CrossRef]
- Finzi, D.; Hermankova, M.; Pierson, T.; Carruth, L.M.; Buck, C.; Chaisson, R.E.; Quinn, T.C.; Chadwick, K.; Margolick, J.; Brookmeyer, R.; et al. Identification of a Reservoir for HIV-1 in Patients on Highly Active Antiretroviral Therapy. Science 1997, 278, 1295–1300. [Google Scholar] [CrossRef]
- Khoury, G.; Darcis, G.; Lee, M.Y.; Bouchat, S.; Van Driessche, B.; Purcell, D.F.J.; Van Lint, C. The Molecular Biology of HIV Latency. In HIV Vaccines and Cure: The Path Towards Finding an Effective Cure and Vaccine; Zhang, L., Lewin, S.R., Eds.; Advances in Experimental Medicine and Biology; Springer Singapore: Singapore, 2018; pp. 187–212. ISBN 9789811304842. [Google Scholar]
- Cohn, L.B.; Chomont, N.; Deeks, S.G. The Biology of the HIV-1 Latent Reservoir and Implications for Cure Strategies—ScienceDirect. Available online: https://www.sciencedirect.com/science/article/pii/S1931312820301785 (accessed on 25 April 2022).
- Börner, C.; Kraus, J. Inhibition of NF-ΚB by Opioids in T Cells. J. Immunol. 2013, 191, 4640–4647. [Google Scholar] [CrossRef]
- Börner, C.; Warnick, B.; Smida, M.; Hartig, R.; Lindquist, J.A.; Schraven, B.; Höllt, V.; Kraus, J. Mechanisms of Opioid-Mediated Inhibition of Human T Cell Receptor Signaling. J. Immunol. 2009, 183, 882–889. [Google Scholar] [CrossRef]
- Azzoni, L.; Metzger, D.; Montaner, L.J. Effect of Opioid Use on Immune Activation and HIV Persistence on ART. J. Neuroimmune Pharmacol. 2020, 15, 643–657. [Google Scholar] [CrossRef]
- Acharya, A.; Olwenyi, O.A.; Thurman, M.; Pandey, K.; Morsey, B.M.; Lamberty, B.; Ferguson, N.; Callen, S.; Fang, Q.; Buch, S.J.; et al. Chronic Morphine Administration Differentially Modulates Viral Reservoirs in a Simian Immunodeficiency Virus SIVmac251-Infected Rhesus Macaque Model. J. Virol. 2020. [Google Scholar] [CrossRef]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; American Psychiatric Publishing: Washington, DC, USA, 2013. [Google Scholar]
- Agosto, L.M.; Hirnet, J.B.; Michaels, D.H.; Shaik-Dasthagirisaheb, Y.B.; Gibson, F.C.; Viglianti, G.; Henderson, A.J. Porphyromonas Gingivalis-Mediated Signaling through TLR4 Mediates Persistent HIV Infection of Primary Macrophages. Virology 2016, 499, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Liszewski, M.K.; Yu, J.J.; O’Doherty, U. Detecting HIV-1 Integration by Repetitive-Sampling Alu-Gag PCR. Methods 2009, 47, 254–260. [Google Scholar] [CrossRef] [PubMed]
- Gagne, M.; Michaels, D.; Lester, G.M.S.; Gummuluru, S.; Wong, W.W.; Henderson, A.J. Strength of T Cell Signaling Regulates HIV-1 Replication and Establishment of Latency. PLoS Pathogens 2019, 15, e1007802. [Google Scholar] [CrossRef] [PubMed]
- Martins, L.J.; Bonczkowski, P.; Spivak, A.M.; De Spiegelaere, W.; Novis, C.L.; DePaula-Silva, A.B.; Malatinkova, E.; Trypsteen, W.; Bosque, A.; Vanderkerckhove, L.; et al. Modeling HIV-1 Latency in Primary T Cells Using a Replication-Competent Virus. AIDS Res. Hum. Retrovir. 2016, 32, 187–193. [Google Scholar] [CrossRef]
- Bruner, K.M.; Wang, Z.; Simonetti, F.R.; Bender, A.M.; Kwon, K.J.; Sengupta, S.; Fray, E.J.; Beg, S.A.; Antar, A.A.R.; Jenike, K.M.; et al. A Quantitative Approach for Measuring the Reservoir of Latent HIV-1 Proviruses. Nature 2019, 566, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Kuniholm, J.; Armstrong, E.; Bernabe, B.; Coote, C.; Berenson, A.; Patalano, S.D.; Olson, A.; He, X.; Lin, N.H.; Bass, J.I.F.; et al. Intragenic Proviral Elements Support Transcription of Defective HIV-1 Proviruses. PLoS Pathogens 2021, 17, e1009982. [Google Scholar] [CrossRef]
- Bruner, K.M.; Murray, A.J.; Pollack, R.A.; Soliman, M.G.; Laskey, S.B.; Capoferri, A.A.; Lai, J.; Strain, M.C.; Lada, S.M.; Hoh, R.; et al. Defective Proviruses Rapidly Accumulate during Acute HIV-1 Infection. Nat. Med. 2016, 22, 1043–1049. [Google Scholar] [CrossRef] [PubMed]
- Ho, Y.-C.; Shan, L.; Hosmane, N.N.; Wang, J.; Laskey, S.B.; Rosenbloom, D.I.S.; Lai, J.; Blankson, J.N.; Siliciano, J.D.; Siliciano, R.F. Replication-Competent Noninduced Proviruses in the Latent Reservoir Increase Barrier to HIV-1 Cure. Cell 2013, 155, 540–551. [Google Scholar] [CrossRef]
- Kirichenko, A.; Lapovok, I.; Baryshev, P.; van de Vijver, D.A.M.C.; van Kampen, J.J.A.; Boucher, C.A.B.; Paraskevis, D.; Kireev, D. Genetic Features of HIV-1 Integrase Sub-Subtype A6 Predominant in Russia and Predicted Susceptibility to INSTIs. Viruses 2020, 12, 838. [Google Scholar] [CrossRef]
- Kirichenko, A.; Kireev, D.; Lopatukhin, A.; Murzakova, A.; Lapovok, I.; Saleeva, D.; Ladnaya, N.; Gadirova, A.; Ibrahimova, S.; Safarova, A.; et al. Prevalence of HIV-1 Drug Resistance in Eastern European and Central Asian Countries. PLoS ONE 2022, 17, e0257731. [Google Scholar] [CrossRef]
- Thomson, M.M.; Vinogradova, A.; Delgado, E.; Rakhmanova, A.; Yakovlev, A.; Cuevas, M.T.; Muñoz, M.; Pinilla, M.; Vega, Y.; Pérez-Álvarez, L.; et al. Molecular Epidemiology of HIV-1 in St Petersburg, Russia: Predominance of Subtype A, Former Soviet Union Variant, and Identification of Intrasubtype Subclusters. JAIDS J. Acquir. Immune Defic. Syndr. 2009, 51, 332–339. [Google Scholar] [CrossRef] [PubMed]
- Simonetti, F.R.; White, J.A.; Tumiotto, C.; Ritter, K.D.; Cai, M.; Gandhi, R.T.; Deeks, S.G.; Howell, B.J.; Montaner, L.J.; Blankson, J.N.; et al. Intact Proviral DNA Assay Analysis of Large Cohorts of People with HIV Provides a Benchmark for the Frequency and Composition of Persistent Proviral DNA. Proc. Natl. Acad. Sci. USA 2020, 117, 18692–18700. [Google Scholar] [CrossRef]
- Yukl, S.A.; Kaiser, P.; Kim, P.; Telwatte, S.; Joshi, S.K.; Vu, M.; Lampiris, H.; Wong, J.K. HIV Latency in Isolated Patient CD4+ T Cells May Be Due to Blocks in HIV Transcriptional Elongation, Completion, and Splicing. Sci. Transl. Med. 2018, 10. [Google Scholar] [CrossRef] [PubMed]
- Elangovan, R.; Jenks, M.; Yun, J.; Dickson-Tetteh, L.; Kirtley, S.; Hemelaar, J.; WHO-UNAIDS Network for HIV Isolation and Characterisation; Abimiku, A.G.; Agwale, S.; Archibald, C.; et al. Global and Regional Estimates for Subtype-Specific Therapeutic and Prophylactic HIV-1 Vaccines: A Modeling Study. Front. Microbiol. 2021, 12, 690647. [Google Scholar] [CrossRef] [PubMed]
- Kirk, G.D.; Astemborski, J.; Mehta, S.H.; Ritter, K.D.; Laird, G.M.; Bordi, R.; Sekaly, R.; Siliciano, J.D.; Siliciano, R.F. Nonstructured Treatment Interruptions Are Associated With Higher Human Immunodeficiency Virus Reservoir Size Measured by Intact Proviral DNA Assay in People Who Inject Drugs. J. Infect. Dis. 2020. [Google Scholar] [CrossRef] [PubMed]
- Meijerink, H.; Indrati, A.; Utami, F.; Soedarmo, S.; Alisjahbana, B.; Netea, M.G.; van Crevel, R.; Wisaksana, R.; Ven, A.J. van der Heroin Use Is Associated with Suppressed Pro-Inflammatory Cytokine Response after LPS Exposure in HIV-Infected Individuals. PLoS ONE 2015, 10, e0122822. [Google Scholar] [CrossRef]
- Zhu, M.; Xu, Y.; Wang, H.; Shen, Z.; Xie, Z.; Chen, F.; Gao, Y.; Chen, X.; Zhang, Y.; Wu, Q.; et al. Heroin Abuse Results in Shifted RNA Expression to Neurodegenerative Diseases and Attenuation of TNFα Signaling Pathway. Sci Rep 2018, 8, 9231. [Google Scholar] [CrossRef]
- Karagiannis, T.T.; Cleary, J.P.; Gok, B.; Henderson, A.J.; Martin, N.G.; Yajima, M.; Nelson, E.C.; Cheng, C.S. Single Cell Transcriptomics Reveals Opioid Usage Evokes Widespread Suppression of Antiviral Gene Program. Nat. Commun. 2020, 11, 2611. [Google Scholar] [CrossRef]
- Wiese, A.D.; Griffin, M.R.; Schaffner, W.; Stein, C.M.; Greevy, R.A.; Mitchel, E.F.; Grijalva, C.G. Long-Acting Opioid Use and the Risk of Serious Infections: A Retrospective Cohort Study. Clin. Infect. Dis. 2019, 68, 1862–1869. [Google Scholar] [CrossRef]
- Prottengeier, J.; Koutsilieri, E.; Scheller, C. The Effects of Opioids on HIV Reactivation in Latently-Infected T-Lymphoblasts. AIDS Res. Ther. 2014, 11, 17. [Google Scholar] [CrossRef]
- Peterson, P.K.; Gekker, G.; Hu, S.; Anderson, W.R.; Kravitz, F.; Portoghese, P.S.; Balfour, H.H.; Chao, C.C. Morphine Amplifies HIV-1 Expression in Chronically Infected Promonocytes Cocultured with Human Brain Cells. J. Neuroimmunol. 1994, 50, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Bidlack, J.M. Detection and Function of Opioid Receptors on Cells from the Immune System. Clin. Diagn. Lab. Immunol. 2000, 7, 719–723. [Google Scholar] [CrossRef] [PubMed]
- Sharp, B.M. Opioid Receptor Expression and Intracellular Signaling by Cells Involved in Host Defense and Immunity; Landes Bioscience: Austin, TX, USA, 2013. [Google Scholar]
- Chuang, T.K.; Killam, K.F.; Chuang, L.F.; Kung, H.F.; Sheng, W.S.; Chao, C.C.; Yu, L.; Chuang, R.Y. Mu Opioid Receptor Gene Expression in Immune Cells. Biochem. Biophys. Res. Commun. 1995, 216, 922–930. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Belkowski, J.S.; Briscoe, T.; Rogers, T.J. Regulation of Mu Opioid Receptor Expression in Developing T Cells. J Neuroimmune Pharmacol 2012, 7, 835–842. [Google Scholar] [CrossRef]
- Williams, J.P.; Thompson, J.P.; McDonald, J.; Barnes, T.A.; Cote, T.; Rowbotham, D.J.; Lambert, D.G. Human Peripheral Blood Mononuclear Cells Express Nociceptin/Orphanin FQ, but Not μ, δ, or κ Opioid Receptors. Anesth. Analg. 2007, 105, 998–1005. [Google Scholar] [CrossRef]
- Al-Hashimi, M.; Scott, S.W.M.; Thompson, J.P.; Lambert, D.G. Opioids and Immune Modulation: More Questions than Answers. Br. J. Anaesth. 2013, 111, 80–88. [Google Scholar] [CrossRef]
- Anton, P.A.; Mitsuyasu, R.T.; Deeks, S.G.; Scadden, D.T.; Wagner, B.; Huang, C.; Macken, C.; Richman, D.D.; Christopherson, C.; Borellini, F.; et al. Multiple Measures of HIV Burden in Blood and Tissue Are Correlated with Each Other but Not with Clinical Parameters in Aviremic Subjects. AIDS 2003, 17, 53–63. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Assay | Primer Name | Sequence (5′ → 3′) |
|---|---|---|
| Alu-PCR | Albumin F | GCTGTCATCTCTTGTGGGCTGT |
| Albumin R | AAACTCATGGGAGCTGCTGGTT | |
| Albumin probe | CCTGTCATGCCCACACAAATCTCTCC | |
| Alu F | GCCTCCCAAACTGCTGGGATTACA | |
| Gag R | GCTCTCGCACCCATCTCTCTC | |
| HIV-1 ‘R’ F | GCCTCAATAAAGCTTGCCTTGA | |
| HIV-1 ‘U5’ R | TCCACACTGACTAAAAGGGTCTGA | |
| RU5 probe | CCAGAGTCACACAACAGACG |
| Assay | Target Gene | Primer Name | Sequence (5′ → 3′) |
|---|---|---|---|
| RT-qPCR | HIV Tat | Spliced Tat F | TCCCTCAGACCCTTTTAGTCAG |
| Spliced Tat R | CATCTGTCCTCTGTCAGTTTC | ||
| Mu-opioid receptor | OPRM F | TCTACTCCATCGTGTGCGTG | |
| OPRM R | CAGTCTTCATCTTGGTGTATCTGAC | ||
| Delta-opioid receptor | OPRD F | CGGCATCGTCCGGTACACTA | |
| OPRD R | CTTGGCACTCTGGAAAGGCA | ||
| Kappa-opioid receptor | OPRK F | CAGGTGATGCCAAGAGCTGA | |
| OPRK R | CCTGCGGCGCTATGGTT | ||
| RPL13a | RPL13a F | CAAGCGGATGAACACCAAC | |
| RPL13a R | CGCTTTTTCTTGTCGTAGGGG |
| Assay | Target Gene | Primer Name | Sequence (5′ → 3′) |
|---|---|---|---|
| Intact Proviral DNA Assay | HIV Psi | Ψ F | CAGGACTCGGCTTGCTGAGG |
| Ψ R | GCACCCATCTCTCTCCTTCTAGC | ||
| Ψ Probe | ATGGCGTACTCACCAGT | ||
| HIV Env | Env F | AGAGAGAAAAAAGAGCAGT | |
| Env R | GGCCTGTACCGTCAG | ||
| Env intact probe | AGCAGGAAGCACTATGGG | ||
| RPP30 | RPP30-1 F | GATTTGGACCTGCGAGC | |
| RPP30-1 R | GCGGCTGTCTCCACAAG | ||
| RPP30-1 probe | CTGACCTGAAGGCTCT | ||
| RPP30-2 F | GACACAATGTTTGGTACATGGTTAA | ||
| RPP30-2 R | CTTTGCTTTGTATGTTGGCAGAAA | ||
| RPP30-2 probe | CCATCTCACCAATCATTCTCCTTCCTTC |
| Assay | Primer Name | Sequence (5′ → 3′) |
|---|---|---|
| RT-ddPCR | R-U5 F | GCCTCAATAAAGCTTGCCTTGA - |
| Gag R | GGGCGCCACTGCTAGAGA | |
| R-U5/Gag probe | CCAGAGTCACACAACAGACGGGCACA |
| People with Current Opioid Use n = 8 | People without Current Opioid Use n = 11 | |
|---|---|---|
| Age Mean (SD) Median (IQR) | 37.8 (2.7) 38 (4) | 39.18 (10.78) 37 (4.5) |
| Gender Male (n, %) Female (n, %) | 5 (62.5%) 3 (37.5%) | 7 (63.63%) 4 (36.37%) |
| Education ≥9 grades (n, %) | 8 (100.0%) | 11 (100.0%) |
| CD4+ T cell count Mean (SD) Median (IQR) | 558 (312) 502 (393) | (information available for n = 8) 453 (148.6) 481.5 (145.5) |
| Time since first HIV diagnosis Mean (SD) Median (IQR) | 13.3 (6.0) 12.4 (7.5) | 10.52 (5.78) 10.2 (10.4) |
| Current ART Yes (n, %) No (n, %) | 8 (100.0%) 0 (0.0%) | 11 (100.0%) 0 (0.0%) |
| Time since ART initiation, yrs a Mean (SD) Median (IQR) | 0.6 (0.3) 0.6 (0.4) | 4.36 (2.85) 4.1 (2.95) |
| HIV viral load | <LOD * | <LOD ** |
| Opioid use at the time of enrollment, (n, %) | 8 (100.0%) | 0 (0.0%) |
| Other substance use (n, %) | ||
| Alcohol (AUDIT score) | ||
| Alcohol abstainer/low-risk consumption | 5 (62.5%) | 9 (81.81%) |
| Hazardous drinking | 1 (12.5%) | 1 (9.09%) |
| Harmful drinking | 0 (0.0%) | 0 (0.0%) |
| Likelihood of alcohol dependence | 2 (25.0%) | 1 (9.09%) |
| Current tobacco smokers (n, %) | 8 (100.0%) | 6 (54.54%) |
| Current cannabis, past 30 days | 0 (0.0%) | 0 (0.0%) |
| Predictor (Variables) | Odds Ratio (95% CI) | p Value |
|---|---|---|
| Intercept | 138.3 (1.593 to 2,598,796) | 0.1322 |
| Duration on ART (year) | 0.2478 (0.01881 to 0.7682) | 0.1051 |
| Current opioid use | 0.00951 (2.6964 × 10−7 to 0.1565) | 0.0395 |
| Intact provirus per 106 PBMCs | 1.00 (0.9972 to 1.003) | 0.7442 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Basukala, B.; Rossi, S.; Bendiks, S.; Gnatienko, N.; Patts, G.; Krupitsky, E.; Lioznov, D.; So-Armah, K.; Sagar, M.; Cheng, C.; et al. Virally Suppressed People Living with HIV Who Use Opioids Have Diminished Latency Reversal. Viruses 2023, 15, 415. https://doi.org/10.3390/v15020415
Basukala B, Rossi S, Bendiks S, Gnatienko N, Patts G, Krupitsky E, Lioznov D, So-Armah K, Sagar M, Cheng C, et al. Virally Suppressed People Living with HIV Who Use Opioids Have Diminished Latency Reversal. Viruses. 2023; 15(2):415. https://doi.org/10.3390/v15020415
Chicago/Turabian StyleBasukala, Binita, Sarah Rossi, Sally Bendiks, Natalia Gnatienko, Gregory Patts, Evgeny Krupitsky, Dmitry Lioznov, Kaku So-Armah, Manish Sagar, Christine Cheng, and et al. 2023. "Virally Suppressed People Living with HIV Who Use Opioids Have Diminished Latency Reversal" Viruses 15, no. 2: 415. https://doi.org/10.3390/v15020415
APA StyleBasukala, B., Rossi, S., Bendiks, S., Gnatienko, N., Patts, G., Krupitsky, E., Lioznov, D., So-Armah, K., Sagar, M., Cheng, C., & Henderson, A. J. (2023). Virally Suppressed People Living with HIV Who Use Opioids Have Diminished Latency Reversal. Viruses, 15(2), 415. https://doi.org/10.3390/v15020415