
Dietary Vitamin D Mitigates Coronavirus-Induced Lung Inflammation and Damage in Mice

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Virus Propagation and Plaque Assay
2.2. Primary Cultures
2.3. SARS-CoV-2 Analysis in Calu-3 Cells
2.4. Animals and In Vivo MHV-3 Infection
2.5. Sample Collection and Processing
2.6. Histopathology
2.7. Immunofluorescence and Confocal Microscopy
2.8. Cytokines, Calcium and 25(OH)D Measurements
2.9. Quantitative Reverse Transcription Polymerase Chain Reaction (RT-qPCR)
2.10. Statistical Analysis
3. Results
3.1. Vitamin D in Its Biologically Active form Reduces Replication of Coronaviruses In Vitro
3.2. Vitamin D Decreases Susceptibility of Wild-Type Mice to Severe MHV-3 Infection
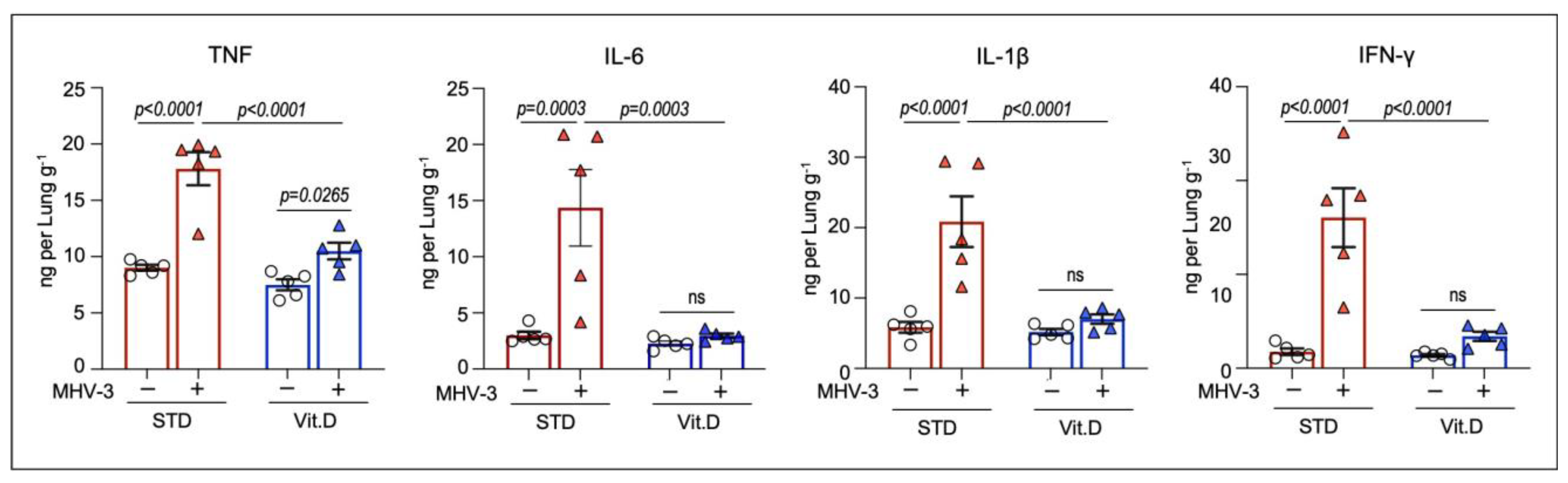
3.3. Vitamin D Mitigates Inflammation and Acute Lung Damage Induced by MHV-3
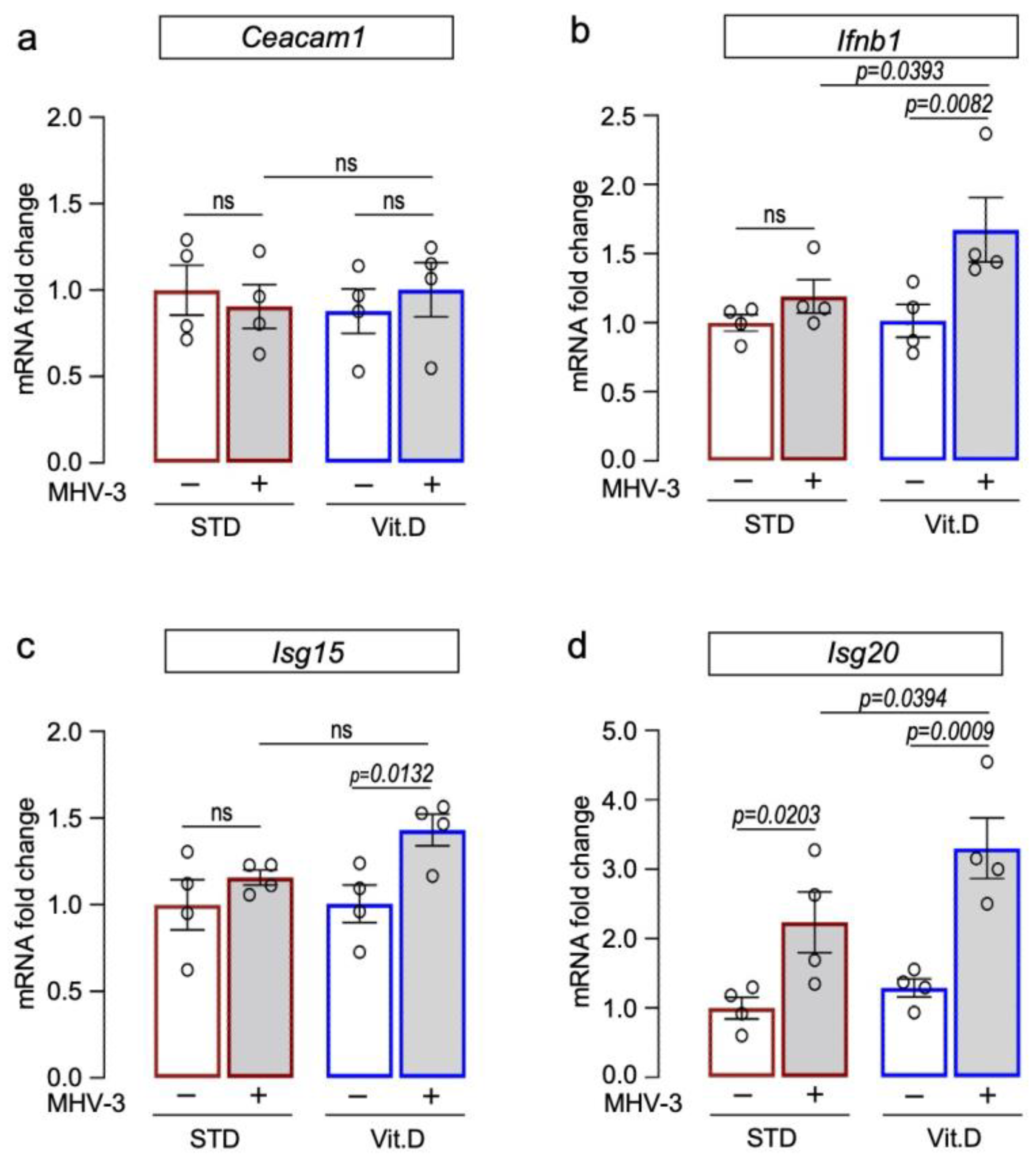
3.4. Vitamin D Supplementation Potentiates Type I Interferon Antiviral Responses against MHV-3
3.5. Vitamin D Reduces Extrapulmonary Damage Caused by MHV-3
4. Discussion
Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bikle, D.; Christakos, S. New Aspects of Vitamin D Metabolism and Action—Addressing the Skin as Source and Target. Nat. Rev. Endocrinol. 2020, 16, 234–252. [Google Scholar] [CrossRef]
- Dimitrov, V.; Barbier, C.; Ismailova, A.; Wang, Y.; Dmowski, K.; Salehi-Tabar, R.; Memari, B.; Groulx-Boivin, E.; White, J.H. Vitamin D-Regulated Gene Expression Profiles: Species-Specificity and Cell-Specific Effects on Metabolism and Immunity. Endocrinology 2021, 162, bqaa218. [Google Scholar] [CrossRef]
- Wang, T.-T.; Nestel, F.P.; Bourdeau, V.; Nagai, Y.; Wang, Q.; Liao, J.; Tavera-Mendoza, L.; Lin, R.; Hanrahan, J.W.; Mader, S.; et al. Cutting Edge: 1,25-Dihydroxyvitamin D3 Is a Direct Inducer of Antimicrobial Peptide Gene Expression. J. Immunol. 2004, 173, 2909–2912. [Google Scholar] [CrossRef] [PubMed]
- Gombart, A.F.; Borregaard, N.; Koeffler, H.P. Human Cathelicidin Antimicrobial Peptide (CAMP) Gene Is a Direct Target of the Vitamin D Receptor and Is Strongly Up-regulated in Myeloid Cells by 1,25-dihydroxyvitamin D 3. FASEB J. 2005, 19, 1067–1077. [Google Scholar] [CrossRef]
- White, J.H. Emerging Roles of Vitamin D-Induced Antimicrobial Peptides in Antiviral Innate Immunity. Nutrients 2022, 14, 284. [Google Scholar] [CrossRef] [PubMed]
- Coussens, A.K.; Wilkinson, R.J.; Hanifa, Y.; Nikolayevskyy, V.; Elkington, P.T.; Islam, K.; Timms, P.M.; Venton, T.R.; Bothamley, G.H.; Packe, G.E.; et al. Vitamin D Accelerates Resolution of Inflammatory Responses during Tuberculosis Treatment. Proc. Natl. Acad. Sci. USA 2012, 109, 15449–15454. [Google Scholar] [CrossRef]
- Hafezi, S.; Saheb Sharif-Askari, F.; Saheb Sharif-Askari, N.; Ali Hussain Alsayed, H.; Alsafar, H.; Al Anouti, F.; Hamid, Q.; Halwani, R. Vitamin D Enhances Type I IFN Signaling in COVID-19 Patients. Sci. Rep. 2022, 12, 17778. [Google Scholar] [CrossRef]
- Charoenngam, N.; Holick, M.F. Immunologic Effects of Vitamin d on Human Health and Disease. Nutrients 2020, 12, 2097. [Google Scholar] [CrossRef]
- Zhu, Z.; Zhu, X.; Gu, L.; Zhan, Y.; Chen, L.; Li, X. Association Between Vitamin D and Influenza: Meta-Analysis and Systematic Review of Randomized Controlled Trials. Front. Nutr. 2022, 8, 799709. [Google Scholar] [CrossRef]
- Leow, L.; Simpson, T.; Cursons, R.; Karalus, N.; Hancox, R.J. Vitamin D, Innate Immunity and Outcomes in Community Acquired Pneumonia. Respirology 2011, 16, 611–616. [Google Scholar] [CrossRef]
- Kim, J.H.; Park, J.-S.; Cho, Y.-J.; Yoon, H.-I.; Song, J.H.; Lee, C.-T.; Lee, J.H. Low Serum 25-Hydroxyvitamin D Level: An Independent Risk Factor for Tuberculosis? Clin. Nutr. 2014, 33, 1081–1086. [Google Scholar] [CrossRef]
- Jolliffe, D.A.; Camargo, C.A.; Sluyter, J.D.; Aglipay, M.; Aloia, J.F.; Ganmaa, D.; Bergman, P.; Bischoff-Ferrari, H.A.; Borzutzky, A.; Damsgaard, C.T.; et al. Vitamin D supplementation to prevent acute respiratory infections: A systematic review and meta-analysis of aggregate data from randomised controlled trials. Lancet Diabetes Endocrinol. 2021, 9, 276–292. [Google Scholar] [CrossRef]
- Petrelli, F.; Luciani, A.; Perego, G.; Dognini, G.; Colombelli, P.L.; Ghidini, A. Therapeutic and prognostic role of vitamin D for COVID-19 infection: A systematic review and meta-analysis of 43 observational studies. J. Steroid Biochem. Mol. Biol. 2021, 211, 105853. [Google Scholar] [CrossRef]
- Butler-Laporte, G.; Nakanishi, T.; Mooser, V.; Morrison, D.R.; Abdullah, T.; Adeleye, O.; Mamlouk, N.; Kimchi, N.; Afrasiabi, Z.; Rezk, N.; et al. Vitamin D and COVID-19 Susceptibility and Severity in the COVID-19 Host Genetics Initiative: A Mendelian Randomization Study. PLoS Med. 2021, 18, e1003605. [Google Scholar] [CrossRef]
- Villasis-Keever, M.A.; López-Alarcón, M.G.; Miranda-Novales, G.; Zurita-Cruz, J.N.; Barrada-Vázquez, A.S.; González-Ibarra, J.; Martínez-Reyes, M.; Grajales-Muñiz, C.; Santacruz-Tinoco, C.E.; Martínez-Miguel, B.; et al. Efficacy and Safety of Vitamin D Supplementation to Prevent COVID-19 in Frontline Healthcare Workers. A Randomized Clinical Trial. Arch. Med. Res. 2022, 53, 423–430. [Google Scholar] [CrossRef]
- Castillo, M.E.; Costa, L.M.E.; Barrios, J.M.V.; Díaz, J.F.A.; Miranda, J.L.; Bouillon, R.; Gomez, J.M.Q. Effect of Calcifediol Treatment and Best Available Therapy versus Best Available Therapy on Intensive Care Unit Admission and Mortality among Patients Hospitalized for COVID-19: A Pilot Randomized Clinical Study. J. Steroid Biochem. Mol. Biol. 2020, 203, 105751. [Google Scholar] [CrossRef]
- Murai, I.H.; Fernandes, A.L.; Sales, L.P.; Pinto, A.J.; Goessler, K.F.; Duran, C.S.C.; Silva, C.B.R.; Franco, A.S.; MacEdo, M.B.; Dalmolin, H.H.H.; et al. Effect of a Single High Dose of Vitamin D3 on Hospital Length of Stay in Patients with Moderate to Severe COVID-19: A Randomized Clinical Trial. JAMA—J. Am. Med. Assoc. 2021, 325, 1053–1060. [Google Scholar] [CrossRef]
- Jolliffe, D.A.; Holt, H.; Greenig, M.; Talaei, M.; Perdek, N.; Pfeffer, P.; Vivaldi, G.; Maltby, S.; Symons, J.; Barlow, N.L.; et al. Effect of a Test-and-Treat Approach to Vitamin D Supplementation on Risk of All Cause Acute Respiratory Tract Infection and COVID-19: Phase 3 Randomised Controlled Trial (CORONAVIT). BMJ 2022, 378, e071230. [Google Scholar] [CrossRef]
- Meng, J.; Li, X.; Liu, W.; Xiao, Y.; Tang, H.; Wu, Y.; Xiong, Y.; Gao, S. The role of vitamin D in the prevention and treatment of SARS-CoV-2 infection: A meta-analysis of randomized controlled trials. Clin. Nutr. 2023, 42, 2198–2206. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, J.; Yang, M.; Wang, Q. Effect of vitamin D supplementation on COVID-19 patients: A systematic review and meta-analysis. Front. Nutr. 2023, 10, 1131103. [Google Scholar] [CrossRef]
- Wang, C.; Wang, S.; Li, D.; Chen, P.; Han, S.; Zhao, G.; Chen, Y.; Zhao, J.; Xiong, J.; Qiu, J.; et al. Human Cathelicidin Inhibits SARS-CoV-2 Infection: Killing Two Birds with One Stone. ACS Infect. Dis. 2021, 7, 1545–1554. [Google Scholar] [CrossRef]
- Arora, J.; Patel, D.R.; Nicol, M.J.; Field, C.J.; Restori, K.H.; Wang, J.; Froelich, N.E.; Katkere, B.; Terwilliger, J.A.; Weaver, V.; et al. Vitamin D and the Ability to Produce 1,25(OH)2D Are Critical for Protection from Viral Infection of the Lungs. Nutrients 2022, 14, 3061. [Google Scholar] [CrossRef]
- Mok, C.K.; Ng, Y.L.; Ahidjo, B.A.; Aw, Z.Q.; Chen, H.; Wong, Y.H.; Lee, R.C.H.; Loe, M.W.C.; Liu, J.; Tan, K.S.; et al. Evaluation of In Vitro and In Vivo Antiviral Activities of Vitamin D for SARS-CoV-2 and Variants. Pharmaceutics 2023, 15, 925. [Google Scholar] [CrossRef]
- Andrade, A.C.d.S.P.; Campolina-Silva, G.H.; Queiroz-Junior, C.M.; de Oliveira, L.C.; Lacerda, L.d.S.B.; Gaggino, J.C.P.; de Souza, F.R.O.; Chaves, I.d.M.; Passos, I.B.; Teixeira, D.C.; et al. A Biosafety Level 2 Mouse Model for Studying Betacoronavirus-Induced Acute Lung Damage and Systemic Manifestations. J. Virol. 2021, 95, e0127621. [Google Scholar] [CrossRef]
- Queiroz-Junior, C.M.; Santos, A.C.; Gonçalves, M.R.; Brito, C.B.; Barrioni, B.; Almeida, P.J.; Gonçalves-Pereira, M.H.; Silva, T.; Oliveira, S.R.; Teixeira, M.; et al. Acute Coronavirus Infection Triggers a TNF-Dependent Osteoporotic Phenotype in Mice. Life Sci. 2023, 324, 121750. [Google Scholar] [CrossRef]
- Vieira-Alves, I.; Alves, A.R.P.; Souza, N.M.V.; Melo, T.L.D.; Coimbra Campos, L.M.D.C.; Lacerda, L.D.S.B.; Queiroz-Junior, C.M.; Andrade, A.C.D.S.P.; Barcelos, L.S.; Teixeira, M.M.; et al. TNF/INOS/NO Pathway Mediates Host Susceptibility to Endothelial-Dependent Circulatory Failure and Death Induced by Betacoronavirus Infection. Clin. Sci. 2023, 137, 543–559. [Google Scholar] [CrossRef]
- Garcia, A.B.; de Moraes, A.P.; Rodrigues, D.M.; Gilioli, R.; de Oliveira-Filho, E.F.; Durães-Carvalho, R.; Arns, C.W. Coding-Complete Genome Sequence of Murine Hepatitis Virus. Microbiol. Resour. Announc. 2021, 10, 10–1128. [Google Scholar] [CrossRef]
- Araujo, D.B.; Machado, R.R.G.; Amgarten, D.E.; Malta, F.d.M.; de Araujo, G.G.; Monteiro, C.O.; Candido, E.D.; Soares, C.P.; de Menezes, F.G.; Pires, A.C.C.; et al. SARS-CoV-2 Isolation from the First Reported Patients in Brazil and Establishment of a Coordinated Task Network. Mem. Inst. Oswaldo Cruz 2020, 115, e200342. [Google Scholar] [CrossRef]
- Marim, F.M.; Silveira, T.N.; Lima, D.S.; Zamboni, D.S. A Method for Generation of Bone Marrow-Derived Macrophages from Cryopreserved Mouse Bone Marrow Cells. PLoS ONE 2010, 5, e15263. [Google Scholar] [CrossRef]
- Campolina-Silva, G.H.; Maria, B.T.; Mahecha, G.A.B.; Oliveira, C.A. Reduced Vitamin D Receptor (VDR) Expression and Plasma Vitamin D Levels Are Associated with Aging-Related Prostate Lesions. Prostate 2018, 78, 532–546. [Google Scholar] [CrossRef]
- Taylor, S.C.; Nadeau, K.; Abbasi, M.; Lachance, C.; Nguyen, M.; Fenrich, J. The Ultimate QPCR Experiment: Producing Publication Quality, Reproducible Data the First Time. Trends Biotechnol. 2019, 37, 761–774. [Google Scholar] [CrossRef]
- Oh, J.; Riek, A.E.; Darwech, I.; Funai, K.; Shao, J.S.; Chin, K.; Sierra, O.L.; Carmeliet, G.; Ostlund, R.E.; Bernal-Mizrachi, C. Deletion of Macrophage Vitamin D Receptor Promotes Insulin Resistance and Monocyte Cholesterol Transport to Accelerate Atherosclerosis in Mice. Cell Rep. 2015, 10, 1872–1886. [Google Scholar] [CrossRef]
- Chu, H.; Chan, J.F.-W.; Yuen, T.T.-T.; Shuai, H.; Yuan, S.; Wang, Y.; Hu, B.; Yip, C.C.-Y.; Tsang, J.O.-L.; Huang, X.; et al. Comparative Tropism, Replication Kinetics, and Cell Damage Profiling of SARS-CoV-2 and SARS-CoV with Implications for Clinical Manifestations, Transmissibility, and Laboratory Studies of COVID-19: An Observational Study. Lancet Microbe 2020, 1, e14–e23. [Google Scholar] [CrossRef]
- Maj, E.; Trynda, J.; Maj, B.; Gębura, K.; Bogunia-Kubik, K.; Chodyński, M.; Kutner, A.; Wietrzyk, J. Differential Response of Lung Cancer Cell Lines to Vitamin D Derivatives Depending on EGFR, KRAS, P53 Mutation Status and VDR Polymorphism. J. Steroid Biochem. Mol. Biol. 2019, 193, 105431. [Google Scholar] [CrossRef]
- Tebben, P.J.; Singh, R.J.; Kumar, R. Vitamin D-Mediated Hypercalcemia: Mechanisms, Diagnosis, and Treatment. Endocr. Rev. 2016, 37, 521–547. [Google Scholar] [CrossRef]
- Otto, G.P.; Rathkolb, B.; Oestereicher, M.A.; Lengger, C.J.; Moerth, C.; Micklich, K.; Fuchs, H.; Gailus-Durner, V.; Wolf, E.; De Angelis, M.H. Clinical Chemistry Reference Intervals for C57BL/6J, C57BL/6N, and C3HeB/FeJ Mice (Mus Musculus). J. Am. Assoc. Lab. Anim. Sci. 2016, 55, 375–386. [Google Scholar]
- Alon, R.; Sportiello, M.; Kozlovski, S.; Kumar, A.; Reilly, E.C.; Zarbock, A.; Garbi, N.; Topham, D.J. Leukocyte Trafficking to the Lungs and beyond: Lessons from Influenza for COVID-19. Nat. Rev. Immunol. 2021, 21, 49–64. [Google Scholar] [CrossRef]
- Yang, Z.; Du, J.; Chen, G.; Zhao, J.; Yang, X.; Su, L.; Cheng, G.; Tang, H. Coronavirus MHV-A59 Infects the Lung and Causes Severe Pneumonia in C57BL/6 Mice. Virol. Sin. 2014, 29, 393–402. [Google Scholar] [CrossRef]
- De Albuquerque, N.; Baig, E.; Ma, X.; Zhang, J.; He, W.; Rowe, A.; Habal, M.; Liu, M.; Shalev, I.; Downey, G.P.; et al. MurineHepatitis Virus Strain 1 Produces a Clinically Relevant Model of Severe Acute Respiratory Syndrome in A/J Mice. J. Virol. 2006, 80, 10382–10394. [Google Scholar] [CrossRef]
- Gong, H.H.; Worley, M.J.; Carver, K.A.; Goldstein, D.R.; Deng, J.C. Neutrophils Drive Pulmonary Vascular Leakage in MHV-1 Infection of Susceptible A/J Mice. Front. Immunol. 2023, 13, 1089064. [Google Scholar] [CrossRef]
- Toro, A.; Arevalo, A.P.; Pereira-Gómez, M.; Sabater, A.; Zizzi, E.A.; Pascual, G.; Lage-Vickers, S.; Porfido, J.L.; Achinelli, I.; Seniuk, R.; et al. Coronavirus Pathogenesis in Mice Explains the SARS-CoV-2 Multi-Organ Spread by Red Blood Cells Hitch-Hiking. medRxiv 2023. [Google Scholar] [CrossRef]
- Liu, J.; Liu, Y.; Xiang, P.; Pu, L.; Xiong, H.; Li, C.; Zhang, M.; Tan, J.; Xu, Y.; Song, R.; et al. Neutrophil-to-Lymphocyte Ratio Predicts Critical Illness Patients with 2019 Coronavirus Disease in the Early Stage. J. Transl. Med. 2020, 18, 206. [Google Scholar] [CrossRef]
- Xu, H.; Li, H.; Cao, D.; Wu, Y.; Chen, Y. Tumor Necrosis Factor α (TNF-α) Receptor-I Is Required for TNF-α-Mediated Fulminant Virus Hepatitis Caused by Murine Hepatitis Virus Strain-3 Infection. Immunol. Lett. 2014, 158, 25–32. [Google Scholar] [CrossRef]
- Karki, R.; Sharma, B.R.; Tuladhar, S.; Williams, E.P.; Zalduondo, L.; Samir, P.; Zheng, M.; Sundaram, B.; Banoth, B.; Malireddi, R.K.S.; et al. Synergism of TNF-α and IFN-γ Triggers Inflammatory Cell Death, Tissue Damage, and Mortality in SARS-CoV-2 Infection and Cytokine Shock Syndromes. Cell 2021, 184, 149–168.e17. [Google Scholar] [CrossRef] [PubMed]
- Sarhan, N.M.; Warda, A.E.A.; Ibrahim, H.S.G.; Schaalan, M.F.; Fathy, S.M. Evaluation of Infliximab/Tocilizumab versus Tocilizumab among COVID-19 Patients with Cytokine Storm Syndrome. Sci. Rep. 2023, 13, 6456. [Google Scholar] [CrossRef] [PubMed]
- Hirai, A.; Ohtsuka, N.; Ikeda, T.; Taniguchi, R.; Blau, D.; Nakagaki, K.; Miura, H.S.; Ami, Y.; Yamada, Y.K.; Itohara, S.; et al. Role of Mouse Hepatitis Virus (MHV) Receptor Murine CEACAM1 in the Resistance of Mice to MHV Infection: Studies of Mice with Chimeric MCEACAM1a and MCEACAM1b. J. Virol. 2010, 84, 6654–6666. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Malireddi, R.K.S.; Bynigeri, R.R.; Mall, R.; Connelly, J.P.; Pruett-Miller, S.M.; Kanneganti, T.D. Inflammatory Cell Death, PANoptosis, Screen Identifies Host Factors in Coronavirus Innate Immune Response as Therapeutic Targets. Commun. Biol. 2023, 6, 1071. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.W.; Ning, Q.; Liu, M.F.; Lai, A.; Leibowitz, J.; Peltekian, K.M.; Cole, E.H.; Fung, L.S.; Holloway, C.; Marsden, P.A.; et al. Fulminant Hepatic Failure in Murine Hepatitis Virus Strain 3 Infection: Tissue-Specific Expression of a Novel Fgl2 Prothrombinase. J. Virol. 1998, 72, 3504. [Google Scholar] [CrossRef]
- Dissanayake, H.A.; de Silva, N.L.; Sumanatilleke, M.; de Silva, S.D.N.; Gamage, K.K.K.; Dematapitiya, C.; Kuruppu, D.C.; Ranasinghe, P.; Pathmanathan, S.; Katulanda, P. Prognostic and Therapeutic Role of Vitamin D in COVID-19: Systematic Review and Meta-Analysis. J. Clin. Endocrinol. Metab. 2022, 107, 1484–1502. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, T.; Gupta, G.K.; Agrawal, D.K. Vitamin D Supplementation Reduces Airway Hyperresponsiveness and Allergic Airway Inflammation in a Murine Model. Clin. Exp. Allergy 2013, 43, 672–683. [Google Scholar] [CrossRef] [PubMed]
- Fischer, K.D.; Hall, S.C.; Agrawal, D.K. Vitamin D Supplementation Reduces Induction of Epithelial-Mesenchymal Transition in Allergen Sensitized and Challenged Mice. PLoS ONE 2016, 11, e0149180. [Google Scholar] [CrossRef]
- Chen, Y.C.; Sung, H.C.; Chuang, T.Y.; Lai, T.C.; Lee, T.L.; Lee, C.W.; Lee, I.T.; Chen, Y.L. Vitamin D3 Decreases TNF-α-Induced Inflammation in Lung Epithelial Cells through a Reduction in Mitochondrial Fission and Mitophagy. Cell Biol. Toxicol. 2022, 38, 427–450. [Google Scholar] [CrossRef] [PubMed]
- Ray, A.D.; Personius, K.E.; Williamson, D.L.; Dungan, C.M.; Dhillon, S.S.; Hershberger, P.A. Vitamin D3 Intake Modulates Diaphragm but Not Peripheral Muscle Force in Young Mice. J. Appl. Physiol. 2016, 120, 1124–1131. [Google Scholar] [CrossRef] [PubMed]
- Verone-Boyle, A.R.; Shoemaker, S.; Attwood, K.; Morrison, C.D.; Makowski, A.J.; Battaglia, S.; Hershberger, P.A. Diet-Derived 25-Hydroxyvitamin D3 Activates Vitamin D Receptor Target Gene Expression and Suppresses EGFR Mutant Non-Small Cell Lung Cancer Growth in Vitro and in Vivo. Oncotarget 2016, 7, 995–1013. [Google Scholar] [CrossRef] [PubMed]
- Maestro, M.A.; Molnár, F.; Carlberg, C. Vitamin D and Its Synthetic Analogs. J. Med. Chem. 2019, 62, 6854–6875. [Google Scholar] [CrossRef] [PubMed]
- Cervantes-Barragán, L.; Kalinke, U.; Züst, R.; König, M.; Reizis, B.; López-Macías, C.; Thiel, V.; Ludewig, B. Type I IFN-Mediated Protection of Macrophages and Dendritic Cells Secures Control of Murine Coronavirus Infection. J. Immunol. 2009, 182, 1099–1106. [Google Scholar] [CrossRef]
- Sharma, L.; Peng, X.; Qing, H.; Hilliard, B.K.; Kim, J.; Swaminathan, A.; Tian, J.; Israni-Winger, K.; Zhang, C.; Habet, V.; et al. Distinct Roles of Type I and Type III Interferons during a Native Murine β Coronavirus Lung Infection. J. Virol. 2022, 96, e0124121. [Google Scholar] [CrossRef]
- Bastard, P.; Rosen, L.B.; Zhang, Q.; Michailidis, E.; Hoffmann, H.H.; Zhang, Y.; Dorgham, K.; Philippot, Q.; Rosain, J.; Béziat, V.; et al. Autoantibodies against Type I IFNs in Patients with Life-Threatening COVID-19. Science 2020, 370, eabd4585. [Google Scholar] [CrossRef]
- Zhang, Q.; Liu, Z.; Moncada-Velez, M.; Chen, J.; Ogishi, M.; Bigio, B.; Yang, R.; Arias, A.A.; Zhou, Q.; Han, J.E.; et al. Inborn Errors of Type I IFN Immunity in Patients with Life-Threatening COVID-19. Science 2020, 370, eabd4570. [Google Scholar] [CrossRef]
- Blanco-Melo, D.; Nilsson-Payant, B.E.; Liu, W.C.; Uhl, S.; Hoagland, D.; Møller, R.; Jordan, T.X.; Oishi, K.; Panis, M.; Sachs, D.; et al. Imbalanced Host Response to SARS-CoV-2 Drives Development of COVID-19. Cell 2020, 181, 1036–1045.e9. [Google Scholar] [CrossRef]
- Hadjadj, J.; Yatim, N.; Barnabei, L.; Corneau, A.; Boussier, J. Impaired Type I Interferon Activity and Inflammatory Responses in Severe COVID-19 Patients. Science 2020, 369, 718–724. [Google Scholar] [CrossRef]
- Li, R.; Pei, S.; Chen, B.; Song, Y.; Zhang, T.; Yang, W.; Shaman, J. Substantial Undocumented Infection Facilitates the Rapid Dissemination of Novel Coronavirus (SARS-CoV-2). Science 2020, 368, 489–493. [Google Scholar] [CrossRef]
- King, C.; Sprent, J. Dual Nature of Type I Interferons in SARS-CoV-2-Induced Inflammation. Trends Immunol. 2021, 42, 312–322. [Google Scholar] [CrossRef] [PubMed]
- Lucas, C.; Wong, P.; Klein, J.; Castro, T.B.R.; Silva, J.; Ellingson, M.K.; Mao, T.; Oh, J.E.; Israelow, B.; Takahashi, T.; et al. Longitudinal Analyses Reveal Immunological Misfiring in Severe COVID-19. Nature 2021, 584, 463–469. [Google Scholar] [CrossRef] [PubMed]
- Pelaia, C.; Tinello, C.; Vatrella, A.; De Sarro, G.; Pelaia, G. Lung under Attack by COVID-19-Induced Cytokine Storm: Pathogenic Mechanisms and Therapeutic Impications. Ther. Adv. Respir. Dis. 2020, 14, 1753466620933508. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M.; Williams, E.P.; Subbarao Malireddi, R.K.; Karki, R.; Banoth, B.; Burton, A.; Webby, R.; Channappanavar, R.; Jonsson, C.B.; Kanneganti, T.D. Impaired NLRP3 Inflammasome Activation/Pyroptosis Leads to Robust Inflammatory Cell Death via Caspase-8/RIPK3 during Coronavirus Infection. J. Biol. Chem. 2020, 295, 14040–14052. [Google Scholar] [CrossRef] [PubMed]
- Pan, P.; Shen, M.; Yu, Z.; Ge, W.; Chen, K.; Tian, M.; Xiao, F.; Wang, Z.; Wang, J.; Jia, Y.; et al. SARS-CoV-2 N Protein Promotes NLRP3 Inflammasome Activation to Induce Hyperinflammation. Nat. Commun. 2021, 12, 4664. [Google Scholar] [CrossRef]
- Zeng, J.; Xie, X.; Feng, X.L.; Xu, L.; Han, J.B.; Yu, D.; Zou, Q.C.; Liu, Q.; Li, X.; Ma, G.; et al. Specific Inhibition of the NLRP3 Inflammasome Suppresses Immune Overactivation and Alleviates COVID-19 like Pathology in Mice. eBioMedicine 2022, 75, 103803. [Google Scholar] [CrossRef]
- Körner, R.W.; Majjouti, M.; Alejandre Alcazar, M.A.; Mahabir, E. Of Mice and Men: The Coronavirus Mhv and Mouse Models as a Translational Approach to Understand Sars-Cov-2. Viruses 2020, 12, 880. [Google Scholar] [CrossRef]
- Amrein, K.; Scherkl, M.; Hoffmann, M.; Neuwersch-Sommeregger, S.; Köstenberger, M.; Tmava Berisha, A.; Martucci, G.; Pilz, S.; Malle, O. Vitamin D deficiency 2.0: An update on the current status worldwide. Eur. J. Clin. Nutr. 2020, 74, 1498–1513. [Google Scholar] [CrossRef]
- Qiao, W.; Lau, H.E.; Xie, H.; Poon, V.K.M.; Chan, C.C.S.; Chu, H.; Yuan, S.; Yuen, T.T.T.; Chik, K.K.H.; Tsang, J.O.L.; et al. SARS-CoV-2 infection induces inflammatory bone loss in golden Syrian hamsters. Nat. Commun. 2022, 13, 2539. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | NCBI Reference | Forward Primer | Reverse Primer |
|---|---|---|---|
| Gapdh | NM_001289726.2 | 5′-AGGTCGGTGTGAACGGATTTG-3′ | 5′-TGTAGACCATGTAGTTGAGGTCA-3′ |
| Actb | NM_007393.5 | 5′-ATGTTTGAGACCTTCAACA-3′ | 5′-CACGTCAGACTTCATGATGG-3′ |
| Ceacam1 | NM_001039185.1 | 5′-CTTGGAGCCTTTGCCTGGTA-3′ | 5′-ATCTCTCTGCCGCTGTATGC-3′ |
| Ifnb1 | NM_010510.2 | 5′-ACTCATGAAGTACAACAGCTACG-3′ | 5′-GGCATCAACTGACAGGTCTT-3′ |
| Isg15 | NM_015783.3 | 5′-CTGCAGCAATGGCCTGGGACCT-3′ | 5′-AGTTTGGTGGGCCAGGCGCT-3′ |
| Isg20 | NM_001113527.1 | 5′-GCCTGGAGGGCTGTTGGTTCTTG-3′ | 5′-CTGCCATGCTCCTTGGCGACC-3′ |
| Variable | Standard Diet (1000 IU Vitamin D3/kg Diet) | Vitamin D Rich Diet (10,000 IU Vitamin D3/kg Diet) | p Value |
|---|---|---|---|
| Serum 25(OH)D | 24.75 ± 3.39 ng/mL | 48.84 ± 4.80 ng/mL | 0.0060 |
| Serum calcium | 5.724 ± 0.35 mg/dL | 6.870 ± 0.57 mg/dL | 0.1230 |
| Mean food intake per mouse | |||
| Week 1 | 2.312 ± 0.09 g/day | 2.434 ± 0.09 g/day | 0.3659 |
| Week 2 | 2.545 ± 0.23 g/day | 2.651 ± 0.02 g/day | 0.6584 |
| Body weight (% from baseline) | |||
| Baseline | 17.52 ± 0.55 g (100.0) | 17.86 ± 0.59 g (100.0) | 0.6783 |
| Week 1 | 17.42 ± 0.50 g (99.43) | 17.41 ± 0.43 g (97.48) | 0.9907 |
| Week 2 | 18.30 ± 0.63 g (104.4) | 17.90 ± 0.46 g (100.2) | 0.6265 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Campolina-Silva, G.; Andrade, A.C.d.S.P.; Couto, M.; Bittencourt-Silva, P.G.; Queiroz-Junior, C.M.; Lacerda, L.d.S.B.; Chaves, I.d.M.; de Oliveira, L.C.; Marim, F.M.; Oliveira, C.A.; et al. Dietary Vitamin D Mitigates Coronavirus-Induced Lung Inflammation and Damage in Mice. Viruses 2023, 15, 2434. https://doi.org/10.3390/v15122434
Campolina-Silva G, Andrade ACdSP, Couto M, Bittencourt-Silva PG, Queiroz-Junior CM, Lacerda LdSB, Chaves IdM, de Oliveira LC, Marim FM, Oliveira CA, et al. Dietary Vitamin D Mitigates Coronavirus-Induced Lung Inflammation and Damage in Mice. Viruses. 2023; 15(12):2434. https://doi.org/10.3390/v15122434
Chicago/Turabian StyleCampolina-Silva, Gabriel, Ana Cláudia dos Santos Pereira Andrade, Manoela Couto, Paloma G. Bittencourt-Silva, Celso M. Queiroz-Junior, Larisse de Souza B. Lacerda, Ian de Meira Chaves, Leonardo C. de Oliveira, Fernanda Martins Marim, Cleida A. Oliveira, and et al. 2023. "Dietary Vitamin D Mitigates Coronavirus-Induced Lung Inflammation and Damage in Mice" Viruses 15, no. 12: 2434. https://doi.org/10.3390/v15122434
APA StyleCampolina-Silva, G., Andrade, A. C. d. S. P., Couto, M., Bittencourt-Silva, P. G., Queiroz-Junior, C. M., Lacerda, L. d. S. B., Chaves, I. d. M., de Oliveira, L. C., Marim, F. M., Oliveira, C. A., da Silva, G. S. F., Teixeira, M. M., & Costa, V. V. (2023). Dietary Vitamin D Mitigates Coronavirus-Induced Lung Inflammation and Damage in Mice. Viruses, 15(12), 2434. https://doi.org/10.3390/v15122434