
Influenza D in Domestic and Wild Animals

Abstract
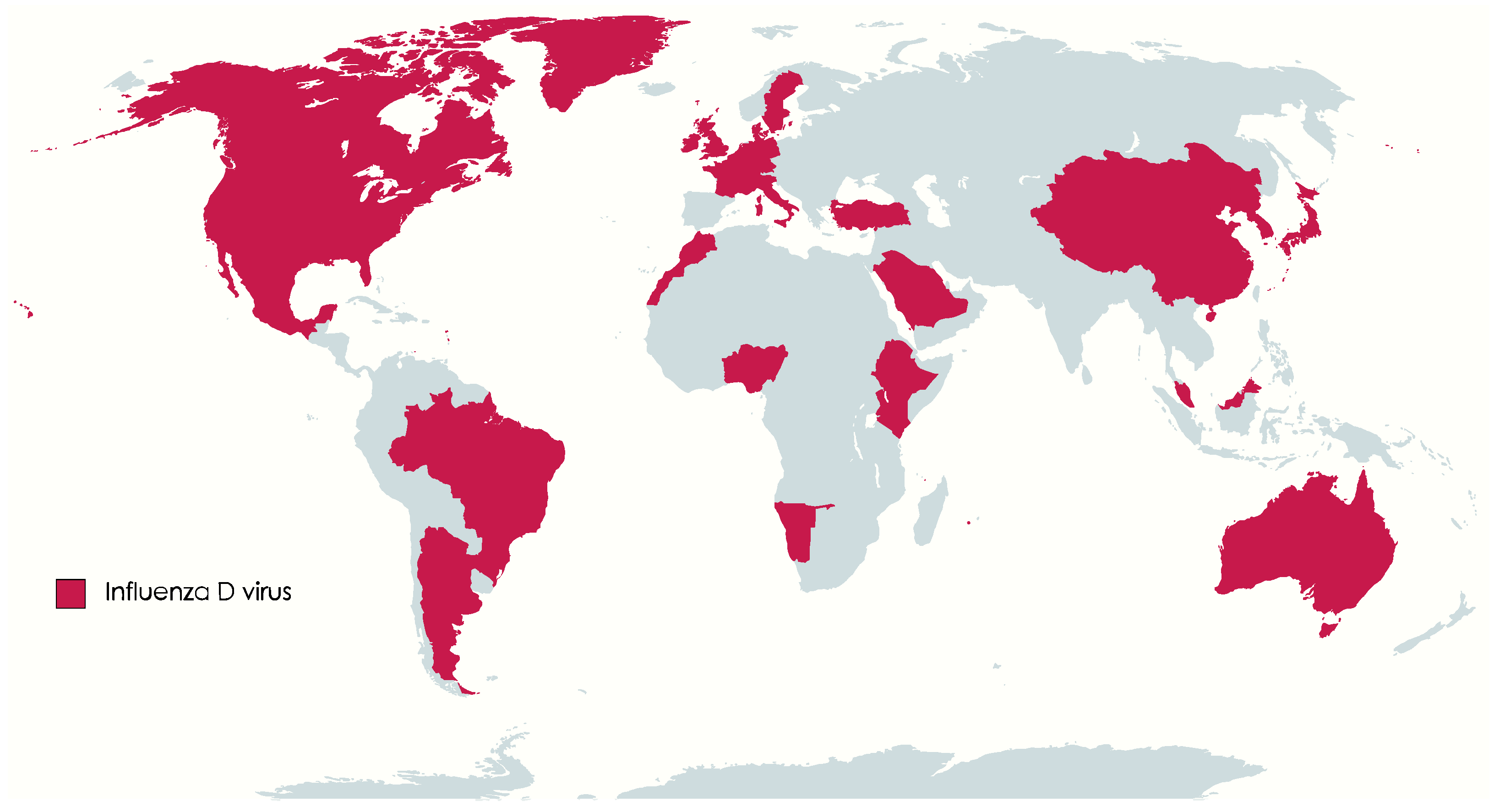
1. Introduction
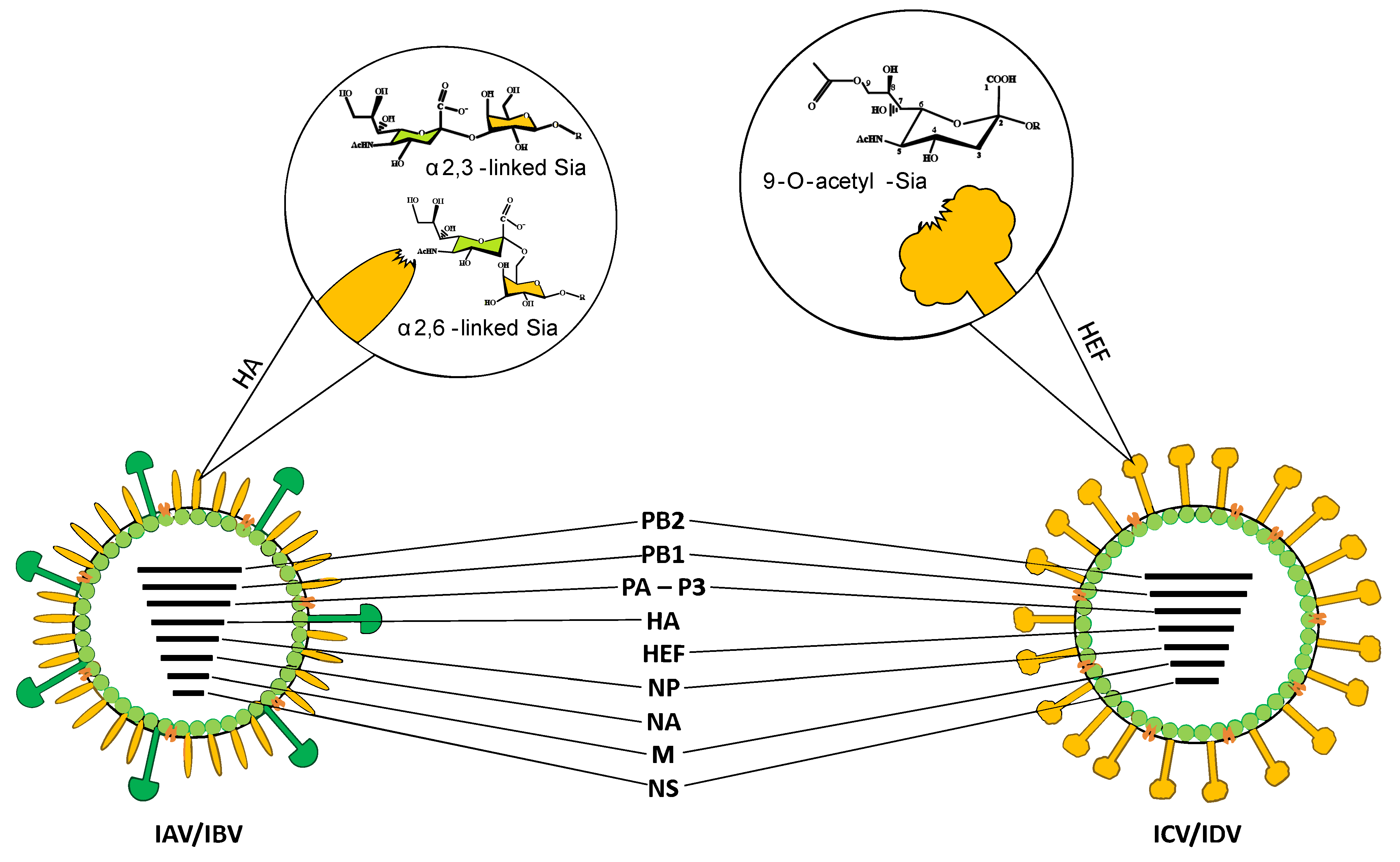
2. Molecular Structure and Receptor Specificity of IDV
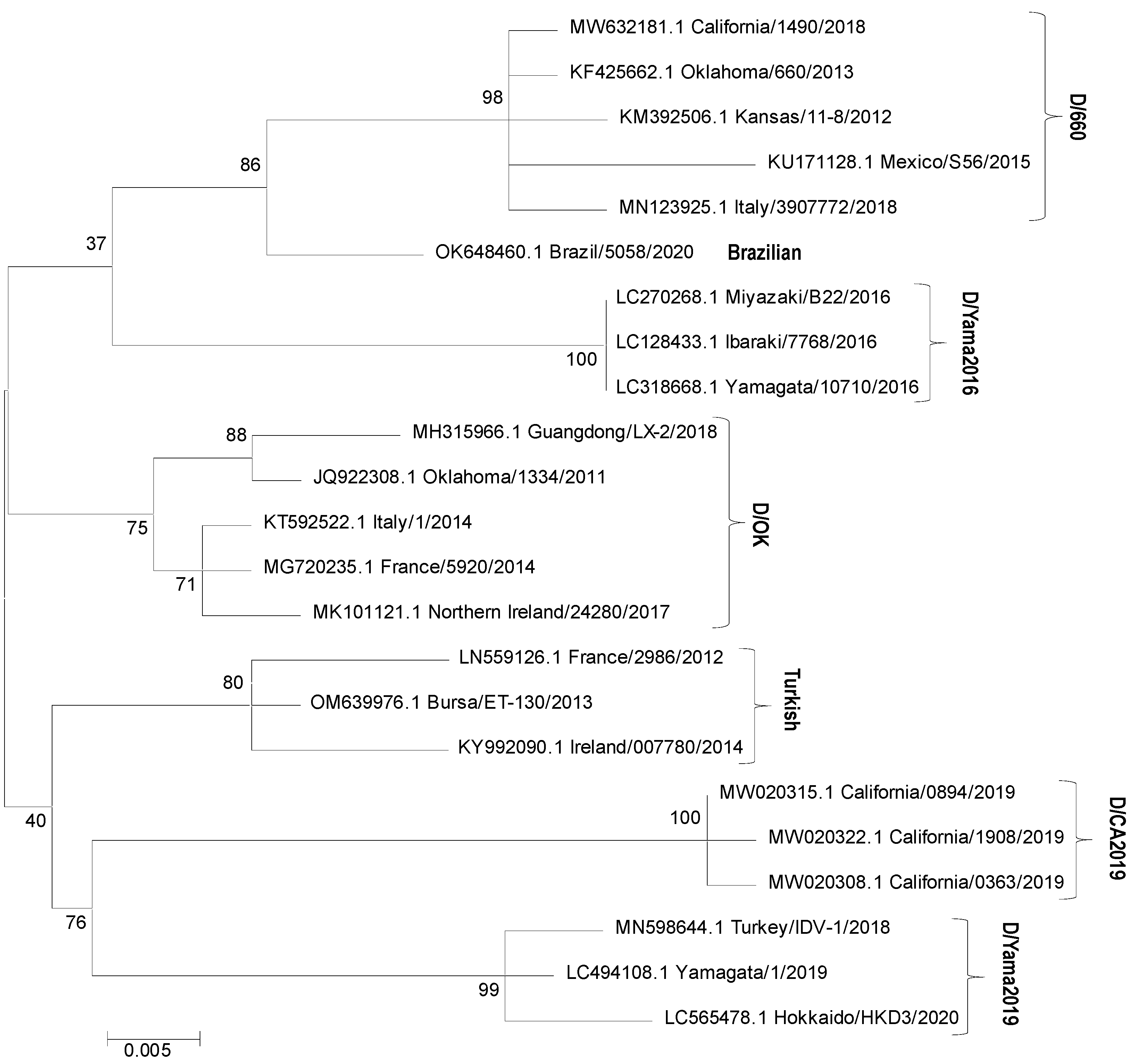
3. Phylogenetics
4. Clinical Signs, Diagnosis, and Potential Vaccines
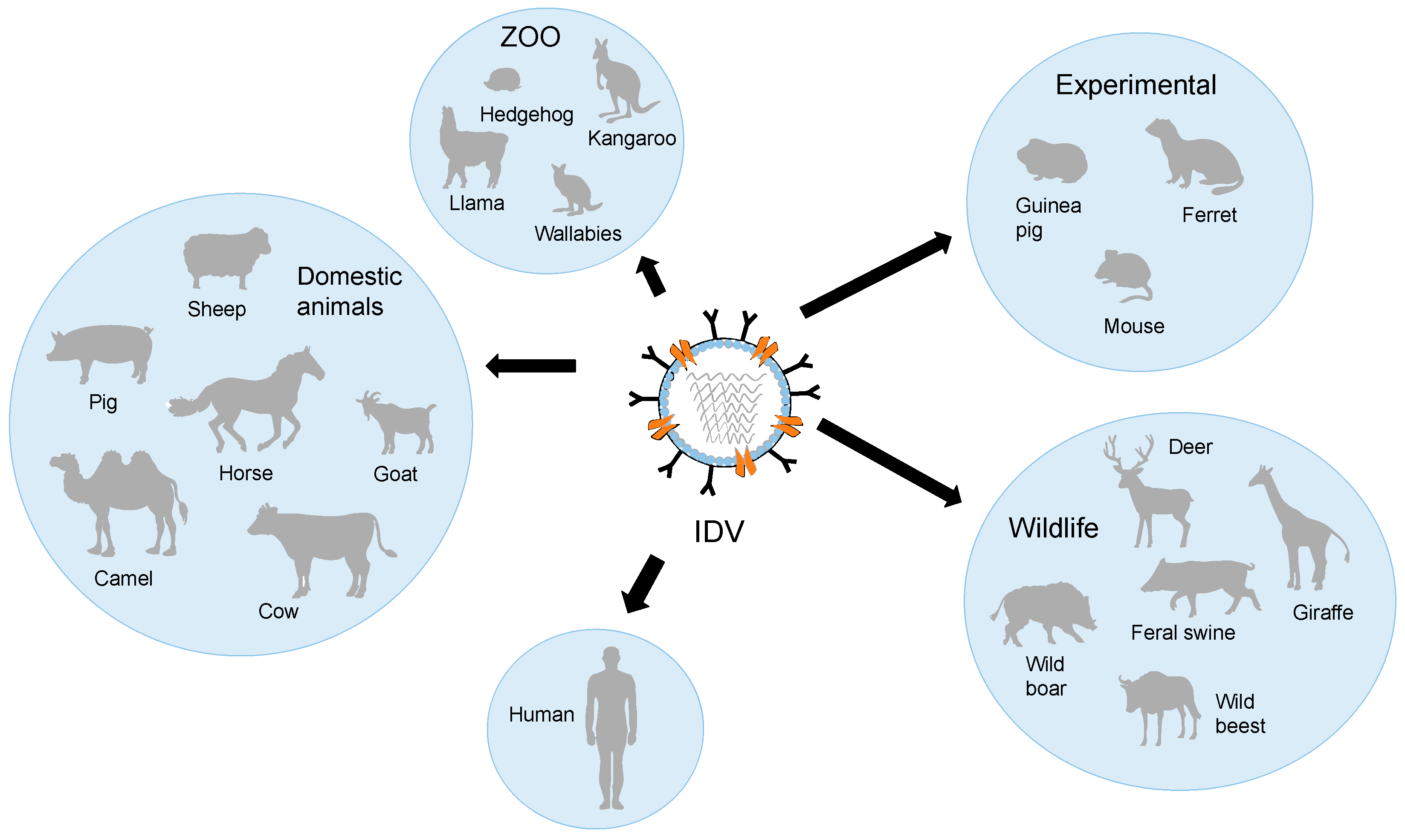
5. Host Range of IDV
5.1. Cattle
5.2. Swine
5.3. Camels
5.4. Small Ruminants
5.5. Horses
5.6. Wild Animals
5.7. Mice, Guinea Pigs, Ferrets, and Selected Cell Lines
6. Zoonotic Potential of IDV
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bouvier, N.M.; Palese, P. The Biology of Influenza Viruses. Vaccine 2008, 26, D49–D53. [Google Scholar] [CrossRef] [PubMed]
- Hause, B.M.; Collin, E.A.; Liu, R.; Huang, B.; Sheng, Z.; Lu, W.; Wang, D.; Nelson, E.A.; Li, F. Characterization of a Novel Influenza Virus in Cattle and Swine: Proposal for a New Genus in the Orthomyxoviridae Family. mBio 2014, 5, e00031-14. [Google Scholar] [CrossRef] [PubMed]
- Long, J.S.; Mistry, B.; Haslam, S.M.; Barclay, W.S. Host and Viral Determinants of Influenza A Virus Species Specificity. Nat. Rev. Microbiol. 2019, 17, 67–81. [Google Scholar] [CrossRef] [PubMed]
- Osterhaus, A.D.M.E.; Rimmelzwaan, G.F.; Martina, B.E.E.; Bestebroer, T.M.; Fouchier, R.A.M. Influenza B Virus in Seals. Science 2000, 288, 1051–1053. [Google Scholar] [CrossRef] [PubMed]
- Wolff, T.; Veit, M. Influenza B, C and D Viruses (Orthomyxoviridae). In Encyclopedia of Virology; Elsevier: Amsterdam, The Netherlands, 2021; pp. 561–574. ISBN 978-0-12-814516-6. [Google Scholar]
- Koutsakos, M.; Kent, S.J. Influenza B Viruses: Underestimated and Overlooked. Microbiol. Aust. 2021, 42, 110–115. [Google Scholar] [CrossRef]
- Horimoto, T.; Hiono, T.; Mekata, H.; Odagiri, T.; Lei, Z.; Kobayashi, T.; Norimine, J.; Inoshima, Y.; Hikono, H.; Murakami, K.; et al. Nationwide Distribution of Bovine Influenza D Virus Infection in Japan. PLoS ONE 2016, 11, e0163828. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Porter, E.; Lohman, M.; Lu, N.; Peddireddi, L.; Hanzlicek, G.; Marthaler, D.; Liu, X.; Bai, J. Influenza C Virus in Cattle with Respiratory Disease, United States, 2016–2018. Emerg. Infect. Dis. 2018, 24, 1926–1929. [Google Scholar] [CrossRef] [PubMed]
- Manuguerra, J.C.; Hannoun, C. Natural Infection of Dogs by Influenza C Virus. Res. Virol. 1992, 143, 199–204. [Google Scholar] [CrossRef]
- Kimura, H.; Abiko, C.; Peng, G.; Muraki, Y.; Sugawara, K.; Hongo, S.; Kitame, F.; Mizuta, K.; Numazaki, Y.; Suzuki, H.; et al. Interspecies Transmission of Influenza C Virus between Humans and Pigs. Virus Res. 1997, 48, 71–79. [Google Scholar] [CrossRef]
- Nedland, H.; Wollman, J.; Sreenivasan, C.; Quast, M.; Singrey, A.; Fawcett, L.; Christopher-Hennings, J.; Nelson, E.; Kaushik, R.S.; Wang, D.; et al. Serological Evidence for the Co-Circulation of Two Lineages of Influenza D Viruses in Equine Populations of the Midwest United States. Zoonoses Public Health 2018, 65, e148–e154. [Google Scholar] [CrossRef]
- Luo, J.; Ferguson, L.; Smith, D.R.; Woolums, A.R.; Epperson, W.B.; Wan, X.-F. Serological Evidence for High Prevalence of Influenza D Viruses in Cattle, Nebraska, United States, 2003–2004. Virology 2017, 501, 88–91. [Google Scholar] [CrossRef] [PubMed]
- Chiapponi, C.; Faccini, S.; De Mattia, A.; Baioni, L.; Barbieri, I.; Rosignoli, C.; Nigrelli, A.; Foni, E. Detection of Influenza D Virus among Swine and Cattle, Italy. Emerg. Infect. Dis. 2016, 22, 352–354. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, M.S.; Mosena, A.C.S.; Baumbach, L.; Demoliner, M.; Gularte, J.S.; Pavarini, S.P.; Driemeier, D.; Weber, M.N.; Spilki, F.R.; Canal, C.W. Cattle Influenza D Virus in Brazil Is Divergent from Established Lineages. Arch. Virol. 2022, 167, 1181–1184. [Google Scholar] [CrossRef] [PubMed]
- Brito, B.P.; Frost, M.J.; Anantanawat, K.; Jaya, F.; Batterham, T.; Djordjevic, S.P.; Chang, W.-S.; Holmes, E.C.; Darling, A.E.; Kirkland, P.D. Expanding the Range of the Respiratory Infectome in Australian Feedlot Cattle with and without Respiratory Disease Using Metatranscriptomics. Microbiome 2023, 11, 158. [Google Scholar] [CrossRef] [PubMed]
- Collin, E.A.; Sheng, Z.; Lang, Y.; Ma, W.; Hause, B.M.; Li, F. Cocirculation of Two Distinct Genetic and Antigenic Lineages of Proposed Influenza D Virus in Cattle. J. Virol. 2015, 89, 1036–1042. [Google Scholar] [CrossRef] [PubMed]
- Salem, E.; Cook, E.A.J.; Lbacha, H.A.; Oliva, J.; Awoume, F.; Aplogan, G.L.; Hymann, E.C.; Muloi, D.; Deem, S.L.; Alali, S.; et al. Serologic Evidence for Influenza C and D Virus among Ruminants and Camelids, Africa, 1991–2015. Emerg. Infect. Dis. 2017, 23, 1556–1559. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.-M.; Wang, S.-C.; Peng, C.; Yu, J.-M.; Zhuang, Q.-Y.; Hou, G.-Y.; Liu, S.; Li, J.-P.; Chen, J.-M. Identification of a Potential Novel Type of Influenza Virus in Bovine in China. Virus Genes 2014, 49, 493–496. [Google Scholar] [CrossRef]
- Oliva, J.; Eichenbaum, A.; Belin, J.; Gaudino, M.; Guillotin, J.; Alzieu, J.-P.; Nicollet, P.; Brugidou, R.; Gueneau, E.; Michel, E.; et al. Serological Evidence of Influenza D Virus Circulation Among Cattle and Small Ruminants in France. Viruses 2019, 11, 516. [Google Scholar] [CrossRef]
- Ruiz, M.; Puig, A.; Bassols, M.; Fraile, L.; Armengol, R. Influenza D Virus: A Review and Update of Its Role in Bovine Respiratory Syndrome. Viruses 2022, 14, 2717. [Google Scholar] [CrossRef]
- Hause, B.M.; Ducatez, M.; Collin, E.A.; Ran, Z.; Liu, R.; Sheng, Z.; Armien, A.; Kaplan, B.; Chakravarty, S.; Hoppe, A.D.; et al. Isolation of a Novel Swine Influenza Virus from Oklahoma in 2011 Which Is Distantly Related to Human Influenza C Viruses. PLoS Pathog. 2013, 9, e1003176. [Google Scholar] [CrossRef]
- Quast, M.; Sreenivasan, C.; Sexton, G.; Nedland, H.; Singrey, A.; Fawcett, L.; Miller, G.; Lauer, D.; Voss, S.; Pollock, S.; et al. Serological Evidence for the Presence of Influenza D Virus in Small Ruminants. Vet. Microbiol. 2015, 180, 281–285. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, L.; Luo, K.; Olivier, A.K.; Cunningham, F.L.; Blackmon, S.; Hanson-Dorr, K.; Sun, H.; Baroch, J.; Lutman, M.W.; Quade, B.; et al. Influenza D Virus Infection in Feral Swine Populations, United States. Emerg. Infect. Dis. 2018, 24, 1020–1028. [Google Scholar] [CrossRef] [PubMed]
- Guan, M.; Jacobson, O.; Sarafianos, G.; Baroch, J.; Deliberto, T.J.; Wan, X.-F. Exposure of White-Tailed Deer in North America to Influenza D Virus. Virology 2022, 573, 111–117. [Google Scholar] [CrossRef]
- Markowska-Daniel, I.; Mickiewicz, M.; Witkowski, L.; Kita, J. Prevalence of Infections with a New Type of Influenza Virus—Influenza D Virus in Humans and Animals. Med. Weter. 2016, 72, 659–665. [Google Scholar] [CrossRef][Green Version]
- Nemanichvili, N.; Berends, A.J.; Tomris, I.; Barnard, K.N.; Parrish, C.R.; Gröne, A.; Rijks, J.M.; Verheije, M.H.; De Vries, R.P. Influenza D Binding Properties Vary amongst the Two Major Virus Clades and Wildlife Species. Vet. Microbiol. 2022, 264, 109298. [Google Scholar] [CrossRef] [PubMed]
- Leibler, J.H.; Abdelgadir, A.; Seidel, J.; White, R.F.; Johnson, W.E.; Reynolds, S.J.; Gray, G.C.; Schaeffer, J.W. Influenza D Virus Exposure among US Cattle Workers: A Call for Surveillance. Zoonoses Public Health 2023, 70, 166–170. [Google Scholar] [CrossRef]
- Bailey, E.S.; Fieldhouse, J.K.; Alarja, N.A.; Chen, D.D.; Kovalik, M.E.; Zemke, J.N.; Choi, J.Y.; Borkenhagen, L.K.; Toh, T.-H.; Lee, J.S.Y.; et al. First Sequence of Influenza D Virus Identified in Poultry Farm Bioaerosols in Sarawak, Malaysia. Trop. Dis. Travel Med. Vaccines 2020, 6, 5. [Google Scholar] [CrossRef]
- King, A.M.Q. Virus Taxonomy: Classification and Nomenclature of Viruses Ninth Report of the International Committee on Taxonomy of Viruses; Elsevier/Academic Press: Amsterdam, The Netherlands, 2012; ISBN 978-0-12-384684-6. [Google Scholar]
- Nakatsu, S.; Murakami, S.; Shindo, K.; Horimoto, T.; Sagara, H.; Noda, T.; Kawaoka, Y. Influenza C and D Viruses Package Eight Organized Ribonucleoprotein Complexes. J. Virol. 2018, 92, e02084-17. [Google Scholar] [CrossRef]
- Dubois, J.; Terrier, O.; Rosa-Calatrava, M. Influenza Viruses and mRNA Splicing: Doing More with Less. mBio 2014, 5, e00070-14. [Google Scholar] [CrossRef]
- Noda, T.; Kawaoka, Y. Structure of Influenza Virus Ribonucleoprotein Complexes and Their Packaging into Virions. Rev. Med. Virol. 2010, 20, 380–391. [Google Scholar] [CrossRef]
- Yu, J.; Hika, B.; Liu, R.; Sheng, Z.; Hause, B.M.; Li, F.; Wang, D. The Hemagglutinin-Esterase Fusion Glycoprotein Is a Primary Determinant of the Exceptional Thermal and Acid Stability of Influenza D Virus. mSphere 2017, 2, e00254-17. [Google Scholar] [CrossRef] [PubMed]
- Varki, A. Multiple Changes in Sialic Acid Biology during Human Evolution. Glycoconj. J. 2009, 26, 231–245. [Google Scholar] [CrossRef] [PubMed]
- Matrosovich, M.; Herrler, G.; Klenk, H.D. Sialic Acid Receptors of Viruses. In SialoGlyco Chemistry and Biology II; Gerardy-Schahn, R., Delannoy, P., Von Itzstein, M., Eds.; Topics in Current Chemistry; Springer International Publishing: Cham, Switzerland, 2013; Volume 367, pp. 1–28. ISBN 978-3-319-21316-3. [Google Scholar]
- Liu, R.; Sreenivasan, C.; Yu, H.; Sheng, Z.; Newkirk, S.J.; An, W.; Smith, D.F.; Chen, X.; Wang, D.; Li, F. Influenza D Virus Diverges from Its Related Influenza C Virus in the Recognition of 9-O-Acetylated N-Acetyl- or N-Glycolyl-Neuraminic Acid-Containing Glycan Receptors. Virology 2020, 545, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Russell, C.J. Hemagglutinin Stability and Its Impact on Influenza A Virus Infectivity, Pathogenicity, and Transmissibility in Avians, Mice, Swine, Seals, Ferrets, and Humans. Viruses 2021, 13, 746. [Google Scholar] [CrossRef] [PubMed]
- Bogdanow, B.; Wang, X.; Eichelbaum, K.; Sadewasser, A.; Husic, I.; Paki, K.; Budt, M.; Hergeselle, M.; Vetter, B.; Hou, J.; et al. The Dynamic Proteome of Influenza A Virus Infection Identifies M Segment Splicing as a Host Range Determinant. Nat. Commun. 2019, 10, 5518. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Wang, R.; Zhang, L.; Shen, B.; Wang, N.; Xu, Q.; He, W.; He, W.; Li, G.; Su, S. Evolutionary Changes of the Novel Influenza D Virus Hemagglutinin-Esterase Fusion Gene Revealed by the Codon Usage Pattern. Virulence 2019, 10, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Li, T.; Wen, Z.; Wu, S.; Wang, Z.; Zheng, J.; Chen, M.; Chen, F.; Wei, W.-K.; Zhai, S.-L.; et al. Identification of D/Yama2019 Lineage-Like Influenza D Virus in Chinese Cattle. Front. Vet. Sci. 2022, 9, 939456. [Google Scholar] [CrossRef] [PubMed]
- Molini, U.; Curini, V.; Jacobs, E.; Tongo, E.; Berjaoui, S.; Hemberger, M.Y.; Puglia, I.; Jago, M.; Khaiseb, S.; Cattoli, G.; et al. First Influenza D Virus Full-Genome Sequence Retrieved from Livestock in Namibia, Africa. Acta Trop. 2022, 232, 106482. [Google Scholar] [CrossRef]
- Murakami, S.; Sato, R.; Ishida, H.; Katayama, M.; Takenaka-Uema, A.; Horimoto, T. Influenza D Virus of New Phylogenetic Lineage, Japan. Emerg. Infect. Dis. 2020, 26, 168–171. [Google Scholar] [CrossRef]
- Yu, J.; Li, F.; Wang, D. The First Decade of Research Advances in Influenza D Virus. J. Gen. Virol. 2021, 102, 1529. [Google Scholar] [CrossRef]
- Huang, C.; Yu, J.; Hause, B.M.; Park, J.Y.; Sreenivasan, C.; Uprety, T.; Sheng, Z.; Wang, D.; Li, F. Emergence of New Phylogenetic Lineage of Influenza D Virus with Broad Antigenicity in California, United States. Emerg. Microbes Infect. 2021, 10, 739–742. [Google Scholar] [CrossRef] [PubMed]
- Yesilbag, K.; Toker, E.B.; Ates, O. Recent Strains of Influenza D Virus Create a New Genetic Cluster for European Strains. Microb. Pathog. 2022, 172, 105769. [Google Scholar] [CrossRef] [PubMed]
- Su, S.; Fu, X.; Li, G.; Kerlin, F.; Veit, M. Novel Influenza D Virus: Epidemiology, Pathology, Evolution and Biological Characteristics. Virulence 2017, 8, 1580–1591. [Google Scholar] [CrossRef] [PubMed]
- Mekata, H.; Yamamoto, M.; Hamabe, S.; Tanaka, H.; Omatsu, T.; Mizutani, T.; Hause, B.M.; Okabayashi, T. Molecular Epidemiological Survey and Phylogenetic Analysis of Bovine Influenza D Virus in Japan. Transbound. Emerg. Dis. 2018, 65, e355–e360. [Google Scholar] [CrossRef] [PubMed]
- Gaudino, M.; Chiapponi, C.; Moreno, A.; Zohari, S.; O’Donovan, T.; Quinless, E.; Sausy, A.; Oliva, J.; Salem, E.; Fusade-Boyer, M.; et al. Evolutionary and Temporal Dynamics of Emerging Influenza D Virus in Europe (2009–22). Virus Evol. 2022, 8, veac081. [Google Scholar] [CrossRef] [PubMed]
- Nemanichvili, N.; Berends, A.; Wubbolts, R.; Gröne, A.; Rijks, J.; De Vries, R.; Verheije, M. Tissue Microarrays to Visualize Influenza D Attachment to Host Receptors in the Respiratory Tract of Farm Animals. Viruses 2021, 13, 586. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, L.; Olivier, A.K.; Genova, S.; Epperson, W.B.; Smith, D.R.; Schneider, L.; Barton, K.; McCuan, K.; Webby, R.J.; Wan, X.-F. Pathogenesis of Influenza D Virus in Cattle. J. Virol. 2016, 90, 5636–5642. [Google Scholar] [CrossRef] [PubMed]
- Salem, E.; Hägglund, S.; Cassard, H.; Corre, T.; Näslund, K.; Foret, C.; Gauthier, D.; Pinard, A.; Delverdier, M.; Zohari, S.; et al. Pathogenesis, Host Innate Immune Response, and Aerosol Transmission of Influenza D Virus in Cattle. J. Virol. 2019, 93, e01853-18. [Google Scholar] [CrossRef]
- Hause, B.M.; Huntimer, L.; Falkenberg, S.; Henningson, J.; Lechtenberg, K.; Halbur, T. An Inactivated Influenza D Virus Vaccine Partially Protects Cattle from Respiratory Disease Caused by Homologous Challenge. Vet. Microbiol. 2017, 199, 47–53. [Google Scholar] [CrossRef]
- Gorin, S.; Fablet, C.; Quéguiner, S.; Barbier, N.; Paboeuf, F.; Hervé, S.; Rose, N.; Simon, G. Assessment of Influenza D Virus in Domestic Pigs and Wild Boars in France: Apparent Limited Spread within Swine Populations Despite Serological Evidence of Breeding Sow Exposure. Viruses 2019, 12, 25. [Google Scholar] [CrossRef]
- Faccini, S.; De Mattia, A.; Chiapponi, C.; Barbieri, I.; Boniotti, M.B.; Rosignoli, C.; Franzini, G.; Moreno, A.; Foni, E.; Nigrelli, A.D. Development and Evaluation of a New Real-Time RT-PCR Assay for Detection of Proposed Influenza D Virus. J. Virol. Methods 2017, 243, 31–34. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, L.; Eckard, L.; Epperson, W.B.; Long, L.-P.; Smith, D.; Huston, C.; Genova, S.; Webby, R.; Wan, X.-F. Influenza D Virus Infection in Mississippi Beef Cattle. Virology 2015, 486, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Moreno, A.; Lelli, D.; Lavazza, A.; Sozzi, E.; Zanni, I.; Chiapponi, C.; Foni, E.; Capucci, L.; Brocchi, E. MAb-based Competitive ELISA for the Detection of Antibodies against Influenza D Virus. Transbound. Emerg. Dis. 2019, 66, 268–276. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.; Kang, G.; Sreenivasan, C.; Daharsh, L.; Zhang, J.; Fan, W.; Wang, D.; Moriyama, H.; Li, F.; Li, Q. A DNA Vaccine Expressing Consensus Hemagglutinin-Esterase Fusion Protein Protected Guinea Pigs from Infection by Two Lineages of Influenza D Virus. J. Virol. 2018, 92, e00110-18. [Google Scholar] [CrossRef] [PubMed]
- Ishida, H.; Murakami, S.; Kamiki, H.; Matsugo, H.; Katayama, M.; Sekine, W.; Ohira, K.; Takenaka-Uema, A.; Horimoto, T. Generation of a Recombinant Temperature-Sensitive Influenza D Virus. Sci. Rep. 2023, 13, 3806. [Google Scholar] [CrossRef] [PubMed]
- Taylor, J.D.; Fulton, R.W.; Lehenbauer, T.W.; Step, D.L.; Confer, A.W. The Epidemiology of Bovine Respiratory Disease: What Is the Evidence for Predisposing Factors? Can. Vet. J. 2010, 51, 1095–1102. [Google Scholar] [PubMed]
- Zhang, M.; Hill, J.E.; Fernando, C.; Alexander, T.W.; Timsit, E.; Meer, F.; Huang, Y. Respiratory Viruses Identified in Western Canadian Beef Cattle by Metagenomic Sequencing and Their Association with Bovine Respiratory Disease. Transbound. Emerg. Dis. 2019, 66, 1379–1386. [Google Scholar] [CrossRef]
- Zhang, X.; Outlaw, C.; Olivier, A.K.; Woolums, A.; Epperson, W.; Wan, X.-F. Pathogenesis of Co-Infections of Influenza D Virus and Mannheimia Haemolytica in Cattle. Vet. Microbiol. 2019, 231, 246–253. [Google Scholar] [CrossRef]
- Mitra, N.; Cernicchiaro, N.; Torres, S.; Li, F.; Hause, B.M. Metagenomic Characterization of the Virome Associated with Bovine Respiratory Disease in Feedlot Cattle Identified Novel Viruses and Suggests an Etiologic Role for Influenza D Virus. J. Gen. Virol. 2016, 97, 1771–1784. [Google Scholar] [CrossRef]
- Silveira, S.; Falkenberg, S.M.; Kaplan, B.S.; Crossley, B.; Ridpath, J.F.; Bauermann, F.B.; Fossler, C.P.; Dargatz, D.A.; Dassanayake, R.P.; Vincent, A.L.; et al. Serosurvey for Influenza D Virus Exposure in Cattle, United States, 2014–2015. Emerg. Infect. Dis. 2019, 25, 2074–2080. [Google Scholar] [CrossRef]
- Saegerman, C.; Gaudino, M.; Savard, C.; Broes, A.; Ariel, O.; Meyer, G.; Ducatez, M.F. Influenza D Virus in Respiratory Disease in Canadian, Province of Québec, Cattle: Relative Importance and Evidence of New Reassortment between Different Clades. Transbound. Emerg. Dis. 2022, 69, 1227–1245. [Google Scholar] [CrossRef]
- Ducatez, M.F.; Pelletier, C.; Meyer, G. Influenza D Virus in Cattle, France, 2011–2014. Emerg. Infect. Dis. 2015, 21, 368. [Google Scholar] [CrossRef] [PubMed]
- Rosignoli, C.; Faccini, S.; Merenda, M.; Chiapponi, C.; de Mattia, A.; Bufalo, G.; Garbarino, C.; Baioni, L.; Bolzoni, L.; Nigrelli, A.; et al. Influenza D Virus Infection in Cattle in Italy. Large Anim. Rev. 2017, 23, 123–128. [Google Scholar]
- O’Donovan, T.; Donohoe, L.; Ducatez, M.F.; Meyer, G.; Ryan, E. Seroprevalence of Influenza D Virus in Selected Sample Groups of Irish Cattle, Sheep and Pigs. Ir. Vet. J. 2019, 72, 11. [Google Scholar] [CrossRef] [PubMed]
- Snoeck, C.J.; Oliva, J.; Pauly, M.; Losch, S.; Wildschutz, F.; Muller, C.P.; Hübschen, J.M.; Ducatez, M.F. Influenza D Virus Circulation in Cattle and Swine, Luxembourg, 2012–2016. Emerg. Infect. Dis. 2018, 24, 1388–1389. [Google Scholar] [CrossRef]
- Flynn, O.; Gallagher, C.; Mooney, J.; Irvine, C.; Ducatez, M.; Hause, B.; McGrath, G.; Ryan, E. Influenza D Virus in Cattle, Ireland. Emerg. Infect. Dis. 2018, 24, 389–391. [Google Scholar] [CrossRef]
- Dane, H.; Duffy, C.; Guelbenzu, M.; Hause, B.; Fee, S.; Forster, F.; McMenamy, M.J.; Lemon, K. Detection of Influenza D Virus in Bovine Respiratory Disease Samples, UK. Transbound. Emerg. Dis. 2019, 66, 2184–2187. [Google Scholar] [CrossRef]
- Studer, E.; Schönecker, L.; Meylan, M.; Stucki, D.; Dijkman, R.; Holwerda, M.; Glaus, A.; Becker, J. Prevalence of BRD-Related Viral Pathogens in the Upper Respiratory Tract of Swiss Veal Calves. Animals 2021, 11, 1940. [Google Scholar] [CrossRef]
- Goecke, N.B.; Liang, Y.; Otten, N.D.; Hjulsager, C.K.; Larsen, L.E. Characterization of Influenza D Virus in Danish Calves. Viruses 2022, 14, 423. [Google Scholar] [CrossRef]
- Ng, T.F.F.; Kondov, N.O.; Deng, X.; Van Eenennaam, A.; Neibergs, H.L.; Delwart, E. A Metagenomics and Case-Control Study to Identify Viruses Associated with Bovine Respiratory Disease. J. Virol. 2015, 89, 5340–5349. [Google Scholar] [CrossRef]
- Alvarez, I.; Hägglund, S.; Näslund, K.; Eriksson, A.; Ahlgren, E.; Ohlson, A.; Ducatez, M.F.; Meyer, G.; Valarcher, J.-F.; Zohari, S. Detection of Influenza D-Specific Antibodies in Bulk Tank Milk from Swedish Dairy Farms. Viruses 2023, 15, 829. [Google Scholar] [CrossRef] [PubMed]
- Zhai, S.-L.; Zhang, H.; Chen, S.-N.; Zhou, X.; Lin, T.; Liu, R.; Lv, D.-H.; Wen, X.-H.; Wei, W.-K.; Wang, D.; et al. Influenza D Virus in Animal Species in Guangdong Province, Southern China. Emerg. Infect. Dis. 2017, 23, 1392–1396. [Google Scholar] [CrossRef] [PubMed]
- Hayakawa, J.; Masuko, T.; Takehana, T.; Suzuki, T. Genetic and Antigenic Characterization and Retrospective Surveillance of Bovine Influenza D Viruses Identified in Hokkaido, Japan from 2018 to 2020. Viruses 2020, 12, 877. [Google Scholar] [CrossRef] [PubMed]
- Lim, E.H.; Lim, S.-I.; Kim, M.J.; Kwon, M.; Kim, M.-J.; Lee, K.-B.; Choe, S.; An, D.-J.; Hyun, B.-H.; Park, J.-Y.; et al. First Detection of Influenza D Virus Infection in Cattle and Pigs in the Republic of Korea. Microorganisms 2023, 11, 1751. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, A.; Umar, S.; Turan, N.; Aydin, O.; Tali, H.E.; Oguzoglu, T.C.; Yilmaz, H.; Richt, J.A.; Ducatez, M.F. First Report of Influenza D Virus Infection in Turkish Cattle with Respiratory Disease. Res. Vet. Sci. 2020, 130, 98–102. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, I.J.; Fort, M.; Pasucci, J.; Moreno, F.; Gimenez, H.; Näslund, K.; Hägglund, S.; Zohari, S.; Valarcher, J.F. Seroprevalence of Influenza D Virus in Bulls in Argentina. J. Vet. Diagn. Investig. 2020, 32, 585–588. [Google Scholar] [CrossRef] [PubMed]
- Gaudino, M.; Moreno, A.; Snoeck, C.J.; Zohari, S.; Saegerman, C.; O’Donovan, T.; Ryan, E.; Zanni, I.; Foni, E.; Sausy, A.; et al. Emerging Influenza D Virus Infection in European Livestock as Determined in Serology Studies: Are We Underestimating Its Spread over the Continent? Transbound. Emerg. Dis. 2021, 68, 1125–1135. [Google Scholar] [CrossRef]
- Foni, E.; Chiapponi, C.; Baioni, L.; Zanni, I.; Merenda, M.; Rosignoli, C.; Kyriakis, C.S.; Luini, M.V.; Mandola, M.L.; Bolzoni, L.; et al. Influenza D in Italy: Towards a Better Understanding of an Emerging Viral Infection in Swine. Sci. Rep. 2017, 7, 11660. [Google Scholar] [CrossRef]
- Lauterbach, S.E.; Nelson, S.W.; Robinson, M.E.; Lorbach, J.N.; Nolting, J.M.; Bowman, A.S. Assessing Exhibition Swine as Potential Disseminators of Infectious Disease through the Detection of Five Respiratory Pathogens at Agricultural Exhibitions. Vet. Res. 2019, 50, 63. [Google Scholar] [CrossRef]
- Sparasci, O.A.; Rizzo, F.; Marchino, M.; Cerutti, F.; Guglielmetti, C.; Grosjacques, F.; Muratore, E.; Sona, B.; Biolatti, P.; Peletto, S.; et al. Influenza D Virus in Cattle and Swine in Piedmont Region, North-Western Italy. Int. J. Infect. Dis. 2022, 116, S92. [Google Scholar] [CrossRef]
- Sanogo, I.N.; Kouakou, C.; Batawui, K.; Djegui, F.; Byarugaba, D.K.; Adjin, R.; Adjabli, K.; Wabwire-Mangen, F.; Erima, B.; Atim, G.; et al. Serological Surveillance of Influenza D Virus in Ruminants and Swine in West and East Africa, 2017–2020. Viruses 2021, 13, 1749. [Google Scholar] [CrossRef] [PubMed]
- Murakami, S.; Odagiri, T.; Melaku, S.K.; Bazartseren, B.; Ishida, H.; Takenaka-Uema, A.; Muraki, Y.; Sentsui, H.; Horimoto, T. Influenza D Virus Infection in Dromedary Camels, Ethiopia. Emerg. Infect. Dis. 2019, 25, 1224–1226. [Google Scholar] [CrossRef] [PubMed]
- Hemida, M.G.; Perera, R.A.; Al Jassim, R.A.; Kayali, G.; Siu, L.Y.; Wang, P.; Chu, K.W.; Perlman, S.; Ali, M.A.; Alnaeem, A.; et al. Seroepidemiology of Middle East Respiratory Syndrome (MERS) Coronavirus in Saudi Arabia (1993) and Australia (2014) and Characterisation of Assay Specificity. Eurosurveillance 2014, 19, 20828. [Google Scholar] [CrossRef] [PubMed]
- Hemida, M.G.; Chu, D.K.W.; Poon, L.L.M.; Perera, R.A.P.M.; Alhammadi, M.A.; Ng, H.-Y.; Siu, L.Y.; Guan, Y.; Alnaeem, A.; Peiris, M. MERS Coronavirus in Dromedary Camel Herd, Saudi Arabia. Emerg. Infect. Dis. 2014, 20, 1231–1234. [Google Scholar] [CrossRef]
- Chan, S.M.S.; Damdinjav, B.; Perera, R.A.P.M.; Chu, D.K.W.; Khishgee, B.; Enkhbold, B.; Poon, L.L.M.; Peiris, M. Absence of MERS-Coronavirus in Bactrian Camels, Southern Mongolia, November 2014. Emerg. Infect. Dis. 2015, 21, 1269–1271. [Google Scholar] [CrossRef] [PubMed]
- Chu, D.K.; Oladipo, J.O.; Perera, R.A.; Kuranga, S.A.; Chan, S.M.; Poon, L.L.; Peiris, M. Middle East Respiratory Syndrome Coronavirus (MERS-CoV) in Dromedary Camels in Nigeria, 2015. Eurosurveillance 2015, 20, 30086. [Google Scholar] [CrossRef] [PubMed]
- Oliva, J. Eco-Epidemiology of Influenza D Virus: Assessment of Host Ranges and Emergence Risk. Ph.D. Dissertation, Université Paul Sabatier—Toulouse III, Toulouse, France, 2019. [Google Scholar]
- Mazzetto, E.; Bortolami, A.; Fusaro, A.; Mazzacan, E.; Maniero, S.; Vascellari, M.; Beato, M.S.; Schiavon, E.; Chiapponi, C.; Terregino, C.; et al. Replication of Influenza D Viruses of Bovine and Swine Origin in Ovine Respiratory Explants and Their Attachment to the Respiratory Tract of Bovine, Sheep, Goat, Horse, and Swine. Front. Microbiol. 2020, 11, 1136. [Google Scholar] [CrossRef]
- Daly, J.M.; Wu, G.; Norman, F.; Preston, T.; Kydd, J.H.; Jengarn, J.; Scott, S.D.; Temperton, N.; Rash, A.; Elton, D.M.; et al. Limited Evidence for Exposure of UK Horses to Influenza D Virus. Equine Vet. J. 2021, 53, 81. [Google Scholar] [CrossRef]
- Sreenivasan, C.C.; Uprety, T.; Reedy, S.E.; Temeeyasen, G.; Hause, B.M.; Wang, D.; Li, F.; Chambers, T.M. Experimental Infection of Horses with Influenza D Virus. Viruses 2022, 14, 661. [Google Scholar] [CrossRef]
- Wyckoff, A.C.; Henke, S.E.; Campbell, T.A.; Hewitt, D.G.; VerCauteren, K.C. Feral Swine Contact with Domestic Swine: A Serologic Survey and Assessment of Potential for Disease Transmission. J. Wildl. Dis. 2009, 45, 422–429. [Google Scholar] [CrossRef]
- Hall, J.S.; Minnis, R.B.; Campbell, T.A.; Barras, S.; DeYoung, R.W.; Pabilonia, K.; Avery, M.L.; Sullivan, H.; Clark, L.; McLean, R.G. Influenza Exposure in United States Feral Swine Populations. J. Wildl. Dis. 2008, 44, 362–368. [Google Scholar] [CrossRef] [PubMed]
- Parys, A.; Vereecke, N.; Vandoorn, E.; Theuns, S.; Van Reeth, K. Surveillance and Genomic Characterization of Influenza A and D Viruses in Swine, Belgium and the Netherlands, 2019–2021. Emerg. Infect. Dis. 2023, 29, 1459–1464. [Google Scholar] [CrossRef] [PubMed]
- Anjorin, A.-A.A.; Moronkeji, G.O.; Temenu, G.O.; Maiyegun, O.A.; Fakorede, C.O.; Ajoseh, S.O.; Salami, W.O.; Abegunrin, R.O.; Amisu, K.O.; Akinyemi, K.O. First Molecular Detection of Influenza D Virus in Cattle from Commercial Farm in Nigeria, Sub-Saharan Africa. One Health Bull. 2023, 3, 19. [Google Scholar] [CrossRef]
- Fusade-Boyer, M.; Pato, P.S.; Komlan, M.; Dogno, K.; Batawui, K.; Go-Maro, E.; McKenzie, P.; Guinat, C.; Secula, A.; Paul, M.; et al. Risk Mapping of Influenza D Virus Occurrence in Ruminants and Swine in Togo Using a Spatial Multicriteria Decision Analysis Approach. Viruses 2020, 12, 128. [Google Scholar] [CrossRef] [PubMed]
- Murakami, S.; Endoh, M.; Kobayashi, T.; Takenaka-Uema, A.; Chambers, J.K.; Uchida, K.; Nishihara, M.; Hause, B.; Horimoto, T. Influenza D Virus Infection in Herd of Cattle, Japan. Emerg. Infect. Dis. 2016, 22, 1517–1519. [Google Scholar] [CrossRef] [PubMed]
- Thielen, P.; Nolting, J.M.; Nelson, S.W.; Mehoke, T.S.; Howser, C.; Bowman, A.S. Complete Genome Sequence of an Influenza D Virus Strain Identified in a Pig with Subclinical Infection in the United States. Microbiol. Resour. Announc. 2019, 8, e01462-18. [Google Scholar] [CrossRef] [PubMed]
- Oliva, J.; Mettier, J.; Sedano, L.; Delverdier, M.; Bourgès-Abella, N.; Hause, B.; Loupias, J.; Pardo, I.; Bleuart, C.; Bordignon, P.J.; et al. Murine Model for the Study of Influenza D Virus. J. Virol. 2020, 94, e01662-19. [Google Scholar] [CrossRef]
- Skelton, R.M.; Shepardson, K.M.; Hatton, A.; Wilson, P.T.; Sreenivasan, C.; Yu, J.; Wang, D.; Huber, V.C.; Rynda-Apple, A. Contribution of Host Immune Responses Against Influenza D Virus Infection Toward Secondary Bacterial Infection in a Mouse Model. Viruses 2019, 11, 994. [Google Scholar] [CrossRef]
- Holwerda, M.; Kelly, J.; Laloli, L.; Stürmer, I.; Portmann, J.; Stalder, H.; Dijkman, R. Determining the Replication Kinetics and Cellular Tropism of Influenza D Virus on Primary Well-Differentiated Human Airway Epithelial Cells. Viruses 2019, 11, 377. [Google Scholar] [CrossRef]
- Trombetta, C.M.; Marchi, S.; Manini, I.; Kistner, O.; Li, F.; Piu, P.; Manenti, A.; Biuso, F.; Sreenivasan, C.; Druce, J.; et al. Influenza D Virus: Serological Evidence in the Italian Population from 2005 to 2017. Viruses 2019, 12, 30. [Google Scholar] [CrossRef]
- Song, H.; Qi, J.; Khedri, Z.; Diaz, S.; Yu, H.; Chen, X.; Varki, A.; Shi, Y.; Gao, G.F. An Open Receptor-Binding Cavity of Hemagglutinin-Esterase-Fusion Glycoprotein from Newly-Identified Influenza D Virus: Basis for Its Broad Cell Tropism. PLoS Pathog. 2016, 12, e1005411. [Google Scholar] [CrossRef]
- Bailey, E.S.; Choi, J.Y.; Fieldhouse, J.K.; Borkenhagen, L.K.; Zemke, J.; Zhang, D.; Gray, G.C. The Continual Threat of Influenza Virus Infections at the Human–Animal Interface. Evol. Med. Public Health 2018, 2018, 192–198. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.Y.; Zemke, J.; Philo, S.E.; Bailey, E.S.; Yondon, M.; Gray, G.C. Aerosol Sampling in a Hospital Emergency Room Setting: A Complementary Surveillance Method for the Detection of Respiratory Viruses. Front. Public Health 2018, 6, 174. [Google Scholar] [CrossRef] [PubMed]
- Borkenhagen, L.K.; Mallinson, K.A.; Tsao, R.W.; Ha, S.-J.; Lim, W.-H.; Toh, T.-H.; Anderson, B.D.; Fieldhouse, J.K.; Philo, S.E.; Chong, K.-S.; et al. Surveillance for Respiratory and Diarrheal Pathogens at the Human-Pig Interface in Sarawak, Malaysia. PLoS ONE 2018, 13, e0201295. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Country, Year | Animal Species | Method | References |
|---|---|---|---|
| EUROPE | |||
| Belgium (BE), 2019 | deer | HI | [90] |
| Denmark (DK), 2019 | bovine | real-time PCR | [72] |
| France (FR), 2009–2018 | swine and wild boar | HI and real-time PCR | [53] |
| France (FR), 2011–2018 | bovine, small ruminants | HI | [19,65] |
| France (FR), 2019 | hedgehogs | HI | [90] |
| Germany (DE), 2019 | deer | HI | [90] |
| Ireland (IE), 2014–2016 | bovine, swine, and sheep | HI and real-time PCR | [69] |
| Italy (IT), 2012–2019 | bovine and swine | PCR and real-time PCR | [13] |
| Luxembourg (LU), 2012–2016 | bovine and swine | HI | [68] |
| Netherlands (NL), 2021 | swine | real-time PCR | [96] |
| Sweden (SE), 2019 | bulk tank milk | ELISA | [74] |
| Switzerland (CH), 2016 | bovine | real-time PCR | [71] |
| United Kingdoms (UK), 2017 | bovine | real-time PCR | [70] |
| AFRICA | |||
| Benin (BJ), 2012–2014 | bovine | HI | [17] |
| Ethiopia (ET), 2019 | camels | HI | [85] |
| Kenya (KE), 2015 | camels | HI | [17] |
| Morocco (MA), 2015 | bovine | HI and MN | [17] |
| Namibia (NA), 2020 | wildebeest and giraffe | real-time PCR | [41] |
| Nigeria (NG), 2021 | bovine | real-time PCR | [97] |
| Nigeria (NG), 2019 | camels | HI | [90] |
| Togo (TG), 2017–2019 | bovine | HI | [98] |
| ASIA | |||
| Japan, Yamagata (JP-06), 2019 | bovine | real-time PCR | [99] |
| Japan, Hokkaido (JP-01), 2018 | bovine | real-time PCR | [76] |
| China, Guangdong (CN-44), 2016 | bovine, swine, and caprine | real-time PCR | [75] |
| China, Shandong (CN-37), 2014 | bovine | real-time PCR | [18] |
| Malaysia (MY), 2018 | swine | real-time PCR | [28] |
| Mongolia (MN), 2019 | camels | HI | [90] |
| Republic of Korea (ROK), 2019 | bovine and swine | HI and real-time PCR | [77] |
| Saudi Arabia (SA), 2019 | camels | HI | [90] |
| Turkey (TR), 2018 | bovine | Real-time PCR | [78] |
| SOUTH AMERICA | |||
| Argentina (ARG), 2013 | bovine | ELISA | [79] |
| Brazil (BR), 2020 | bovine | real-time PCR | [14] |
| NORTH AMERICA | |||
| USA, California (CA), 2018 | bovine | real-time PCR | [44] |
| USA, Hawaii (HI), North Carolina (NC), Oklahoma (OK), Texas (TX), 2012–2013 | feral swine | HI | [23] |
| USA, Kansas (KS), 2010–2012 | bovine | real-time PCR | [20] |
| USA, Kentucky (KY), 2017 | swine | next generation sequencing | [100] |
| USA (samples collected across the country)/2014, 2015 | bovine | HI | [60] |
| USA, Mississippi (MS), Michigan (MI), Minnesota (MN), and Oklahoma (OK), 2011–2017 | deer | HI | [24] |
| USA, Mississippi (MS), 2004–2014 | bovine | real-time PCR, HI | [55] |
| USA, Nebraska (NE), 2003–2004, 2012–2014 | bovine | HI | [12] |
| USA, Oklahoma (OK), 2011–2013 | swine | next generation sequencing | [21] |
| USA, 12 states, 2014 | bovine | real-time PCR | [16] |
| Canada, Quebec (QC), 2020 | bovine | real-time PCR | [64] |
| Mexico (MX), 2015 | bovine | real-time PCR | [62] |
| AUSTRALIA | |||
| Australia—New South Wales (AU), 2019 | bovine | transcriptomics | [15] |
| Australia (AU), 2019 | camels | HI | [90] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kwasnik, M.; Rola, J.; Rozek, W. Influenza D in Domestic and Wild Animals. Viruses 2023, 15, 2433. https://doi.org/10.3390/v15122433
Kwasnik M, Rola J, Rozek W. Influenza D in Domestic and Wild Animals. Viruses. 2023; 15(12):2433. https://doi.org/10.3390/v15122433
Chicago/Turabian StyleKwasnik, Malgorzata, Jerzy Rola, and Wojciech Rozek. 2023. "Influenza D in Domestic and Wild Animals" Viruses 15, no. 12: 2433. https://doi.org/10.3390/v15122433
APA StyleKwasnik, M., Rola, J., & Rozek, W. (2023). Influenza D in Domestic and Wild Animals. Viruses, 15(12), 2433. https://doi.org/10.3390/v15122433