



Molecular Signature of a Novel Alternanthera Yellow Vein Virus Variant Infecting the Ageratum conyzoides Weed in Oman

 ,
,  , and
, and 
Abstract
:1. Introduction
2. Material and Methods
2.1. Specimen Collection and DNA Extraction
2.2. Amplification, Cloning, and Sequencing
2.3. Bioinformatic Analysis
2.4. Population Study
3. Results
3.1. Alternanthera Yellow Vein Oman Virus Is Associated with Ageratum conyzoides
3.2. Tomato Leaf Curl Betasatellite Associated with the Disease
3.3. Recombination Analysis
3.4. Population Study
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Krupovic, M.; Varsani, A.; Kazlauskas, D.; Breitbart, M.; Delwart, E.; Rosario, K.; Zerbini, F.M. Cressdnaviricota: A virus phylum unifying seven families of rep-encoding viruses with single-stranded, circular DNA genomes. J. Virol. 2020, 94, e00582-20. [Google Scholar] [CrossRef] [PubMed]
- Dolja, V.V.; Krupovic, M.; Koonin, E.V. Deep roots and splendid boughs of the global plant virome. Annu. Rev. Phytopathol. 2020, 58, 23–53. [Google Scholar] [CrossRef] [PubMed]
- Gilbertson, R.L.; Batuman, O.; Webster, C.G.; Adkins, S. Role of the Insect Supervectors Bemisia tabaci and Frankliniella occidentalis in the Emergence and Global Spread of Plant Viruses. Annu. Rev. Virol. 2015, 2, 67–93. [Google Scholar] [CrossRef] [PubMed]
- Zerbini, F.M.; Briddon, R.W.; Idris, A.; Martin, D.P.; Moriones, E.; Navas-Castillo, J.; Ictv Report, C. ICTV Virus Taxonomy Profile: Geminiviridae. J. Gen. Virol. 2017, 98, 131–133. [Google Scholar] [CrossRef] [PubMed]
- Fiallo-Olive, E.; Navas-Castillo, J. Molecular and Biological Characterization of a New World Mono-/Bipartite Begomovirus/Deltasatellite Complex Infecting Corchorus siliquosus. Front. Microbiol. 2020, 11, 1755. [Google Scholar] [CrossRef] [PubMed]
- Idriss, M.; Abdallah, N.; Aref, N.; Haridy, G.; Madkour, M. Biotypes of the castor bean whitefly Trialeurodes ricini (Misra) (Hom., Aleyrodidae) in Egypt: Biochemical characterization and efficiency of geminivirus transmission. J. Appl. Entomol. 1997, 121, 501–509. [Google Scholar] [CrossRef]
- Walker, J.; Siddell, S.G.; Lefkowitz, E.J.; Mushegian, A.R.; Adriaenssens, E.M.; Alfenas-Zerbini, P.; Zerbini, F.M. Changes to virus taxonomy and to the International Code of Virus Classification and Nomenclature ratified by the International Committee on Taxonomy of Viruses (2021). Arch. Virol. 2021, 166, 2633–2648. [Google Scholar] [CrossRef]
- Gong, P.; Tan, H.; Zhao, S.; Li, H.; Liu, H.; Ma, Y.; Zhou, X. Geminiviruses encode additional small proteins with specific subcellular localizations and virulence function. Nat. Commun. 2021, 12, 4278. [Google Scholar] [CrossRef]
- Gnanasekaran, P.; Kishorekumar, R.; Bhattacharyya, D.; Vinoth Kumar, R.; Chakraborty, S. Multifaceted role of geminivirus associated betasatellite in pathogenesis. Mol. Plant Pathol. 2019, 20, 1019–1033. [Google Scholar] [CrossRef]
- Shahid, M.S.; Shafiq, M.; Ilyas, M.; Raza, A.; Al-Sadrani, M.N.; Al-Sadi, A.M.; Briddon, R.W. Frequent occurrence of Mungbean yellow mosaic India virus in tomato leaf curl disease affected tomato in Oman. Sci. Rep. 2019, 9, 16634. [Google Scholar] [CrossRef]
- Al Shihi, A.A.; Al Sadi, A.M.; Deadman, M.; Briddon, R.W.; Shahid, M.S. Identification of a distinct strain of Cotton leaf curl Gezira virus infecting tomato in Oman. J. Phytopathol. 2018, 166, 199–205. [Google Scholar] [CrossRef]
- Shahid, M.S.; Al-Sulaimani, H.; Al-Sadi, A.M. Squash Leaf Curl Virus: A New World Bipartite Begomovirus Threatening Squash Production in Oman. Plant Dis. 2020, 104, 2533. [Google Scholar] [CrossRef]
- Porebski, S.; Bailey, L.G.; Baum, B.R. Modification of a CTAB DNA extraction protocol for plants containing high polysaccharide and polyphenol components. Plant Mol. Biol. Report. 1997, 15, 8–15. [Google Scholar] [CrossRef]
- Al-Mabsli, S.S.; Al-Wahaibi, A.K.; Al-Sadi, A.M.; Shahid, M.S. Association of a monopartite begomovirus and associated betasatellite with yellow vein disease of a weed host, Senna italica Mill. in Oman. Virusdisease 2021, 32, 378–380. [Google Scholar] [CrossRef] [PubMed]
- Haible, D.; Kober, S.; Jeske, H. Rolling circle amplification revolutionizes diagnosis and genomics of geminiviruses. J. Virol. Methods 2006, 135, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Muhire, B.M.; Varsani, A.; Martin, D.P. SDT: A virus classification tool based on pairwise sequence alignment and identity calculation. PLoS ONE 2014, 9, e108277. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Martin, D.P.; Murrell, B.; Golden, M.; Khoosal, A.; Muhire, B. RDP4: Detection and analysis of recombination patterns in virus genomes. Virus Evol. 2015, 1, vev003. [Google Scholar] [CrossRef]
- Lima, A.T.M.; Silva, J.C.F.; Silva, F.N.; Castillo-Urquiza, G.P.; Silva, F.F.; Seah, Y.M.; Zerbini, F.M. The diversification of begomovirus populations is predominantly driven by mutational dynamics. Virus Evol. 2017, 3, vex005. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-Delbarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA sequence polymorphism analysis of large data sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Shafiq, M.; Sattar, M.N.; Shahid, M.S.; Al-Sadi, A.M.; Briddon, R.W. Interaction of watermelon chlorotic stunt virus with satellites. Australas. PlantPathol. 2021, 50, 117–128. [Google Scholar] [CrossRef]
- Marabi, R.S.; Das, S.B.; Tripathi, N.; Wada, T.; Noda, H. Identification of begomoviruses from legume crop and weed plants and viruliferous status of the whitefly Bemisia tabaci in Central India. Curr. Sci. 2021, 120, 1240–1246. [Google Scholar] [CrossRef]
- Brown, J.K.; Zerbini, F.M.; Navas-Castillo, J.; Moriones, E.; Ramos-Sobrinho, R.; Silva, J.C.; Varsani, A. Revision of Begomovirus taxonomy based on pairwise sequence comparisons. Arch. Virol. 2015, 160, 1593–1619. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.; Kulshreshtha, A.; Kumar, R.; Hallan, V. First report of natural infection of alternanthera yellow vein virus and cotton leaf curl Multan betasatellite on a new host Picrorhiza kurroa, an important endangered medicinal herb. J. Plant Pathol. 2019, 101, 149–153. [Google Scholar] [CrossRef]
- Murtaza, G.; Mubin, M.; Nawaz-Ul-Rehman, M.S.; Amrao, L. Genetic analysis of alternanthera yellow vein virus (Ayvv) infecting Eclipta prostrata plant in Pakistan. Pak. J. Agric. Sci. 2018, 55, 505–512. [Google Scholar]
- Nawaz-Ul-Rehman, M.; Liaqat, I.; Nahid, N.; Saleem, F.; Alkahtani, S.; Al Qahtani, A.; Mubin, M. Alternanthera yellow vein virus (AYVV); A betasatellite independent begomovirus infecting Sonchus palustris in Pakistan. Braz. J.Biol. 2022, 82, e262248. [Google Scholar] [CrossRef] [PubMed]
- Zaidi, S.S.; Shakir, S.; Farooq, M.; Amin, I.; Mansoor, S. First report of Alternanthera yellow vein virus from Eclipta prostrata in Pakistan. Plant Dis. 2017, 101, 266. [Google Scholar] [CrossRef]
- Zhang, J.; Jia, S.P.; Yang, C.X.; Liu, Z.; Wu, Z.J. Detection and molecular characterization of three begomoviruses associated with yellow vein disease of Eclipta prostrata in fujian, China. J. Plant Pathol. 2015, 97, 161–165. [Google Scholar]
- Mubin, M.; Shahid, M.S.; Tahir, M.N.; Briddon, R.W.; Mansoor, S. Characterization of begomovirus components from a weed suggests that begomoviruses may associate with multiple distinct DNA satellites. Virus Genes 2010, 40, 452–457. [Google Scholar] [CrossRef]
- Ding, M.; Yang, L.; Zhao, Z.W.; Zhang, Z.K. First report of alternanthera yellow vein virus in Eclipta prostrata in China. J. Plant Pathol. 2009, 91, 232. [Google Scholar]
- He, Z.F.; Mao, M.J.; Yu, H.; Wang, X.M.; Li, H.P. First report of a strain of Alternanthera yellow vein virus infecting Eclipta prostrate (L.) L. (compositae) in China. J. Phytopathol. 2008, 156, 496–498. [Google Scholar] [CrossRef]
- Huang, J.F.; Jiang, T.; Zhou, X.P. Molecular characterization of begomoviruses infecting Ludwigia hyssopifolia. J. Plant Pathol. 2006, 88, 83–88. [Google Scholar]
- Guo, X.; Zhou, X. Molecular characterization of Alternanthera yellow vein virus: A new Begomovirus species infecting Alternanthera philoxeroides. J. Phytopathol. 2005, 153, 694–696. [Google Scholar] [CrossRef]
- Rojas, M.R.; Hagen, C.; Lucas, W.J.; Gilbertson, R.L. Exploiting chinks in the plant’s armor: Evolution and emergence of geminiviruses. Annu. Rev. Phytopathol. 2005, 43, 361–394. [Google Scholar] [CrossRef] [PubMed]
- Idris, A.M.; Shahid, M.S.; Briddon, R.W.; Khan, A.; Zhu, J.-K.; Brown, J.K. An unusual alphasatellite associated with monopartite begomoviruses attenuates symptoms and reduces betasatellite accumulation. J. Gen.Virol. 2011, 92, 706–717. [Google Scholar] [CrossRef] [PubMed]
- Ha, C.; Coombs, S.; Revill, P.; Harding, R.; Vu, M.; Dale, J. Molecular characterization of begomoviruses and DNA satellites from Vietnam: Additional evidence that the New World geminiviruses were present in the Old World prior to continental separation. J. Gen. Virol. 2008, 89, 312–326. [Google Scholar] [CrossRef] [PubMed]
- Briddon, R.W.; Brown, J.K.; Moriones, E.; Stanley, J.; Zerbini, M.; Zhou, X.; Fauquet, C.M. Recommendations for the classification and nomenclature of the DNA-beta satellites of begomoviruses. Arch. Virol. 2008, 153, 763–781. [Google Scholar] [CrossRef]
- Briddon, R.W.; Markham, G. Complementation of bipartite begomovirus movement functions by topocuviruses and curtoviruses. Arch. Virol. 2001, 146, 1811–1819. [Google Scholar] [CrossRef]
- Hanley-Bowdoin, L.; Settlage, S.B.; Orozco, B.M.; Nagar, S.; Robertson, D. Geminiviruses: Models for plant DNA replication, transcription, and cell cycle regulation. Crit. Rev. Plant Sci. 1999, 18, 71–106. [Google Scholar] [CrossRef]
- Kharazmi, S.; Behjatnia, S.A.; Hamzehzarghani, H.; Niazi, A. Cotton leaf curl Multan betasatellite as a plant gene delivery vector trans-activated by taxonomically diverse geminiviruses. Arch. Virol. 2012, 157, 1269–1279. [Google Scholar] [CrossRef]
- Saunders, K. Analysis of geminivirus DNA replication by 2-D gel. Methods Mol. Biol. 2008, 451, 135–143. [Google Scholar]
- Nawaz-Ul-Rehman, M.S.; Nahid, N.; Mansoor, S.; Briddon, R.W.; Fauquet, C.M. Post-transcriptional gene silencing suppressor activity of two non-pathogenic alphasatellites associated with a begomovirus. Virology 2010, 405, 300–308. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
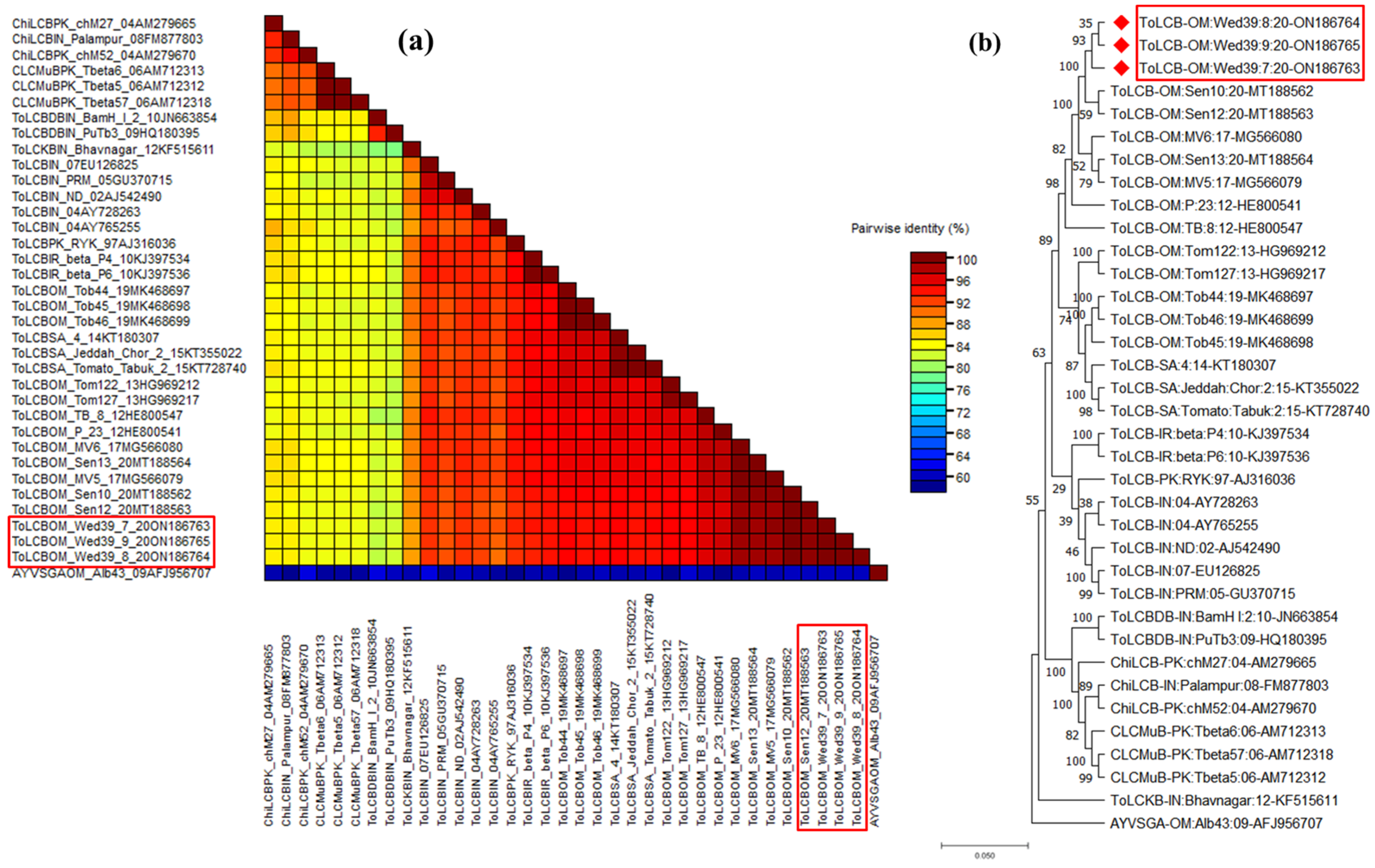{kind=link}
| Alternanthera Yellow Vein Virus Position of Genes (Coordinates)/No. of Amino Acids (Predicted Coding Capacity in kDa) | Tomato Leaf Curl Betasatellite | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Isolate | Acc. No. | Size (nt) | CP | V2 | Rep | TrAP | REn | C4 | Satellite | Acc. No. | Size (nt) | Position of BC1 Gene (Coordinates)/No. of Amino Acids (Predicted Coding Capacity in kDa) |
| Wed39-4 | ON186760 | 2745 | 314–1084 256 | 154–501 115 | 1533–2618 362 | 1226–1630 134 | 1081–1485 134 | 2171–2467 98 | Wed39-7 | ON186763 | 1360 | 189–545 118 (13) |
| Wed39-5 | ON186761 | 2747 | 314–1084 256 | 154–501 115 | 1533–2618 362 | 1226–1630 134 | 1081–1485 134 | 2171–2467 98 | Wed39-8 | ON186764 | 1360 | 189–545 118 (13) |
| Wed39-6 | ON186762 | 2747 | 314–1084 256 | 154–501 115 | 1533–2618 361 | 1226–1630 134 | 1081–1485 134 | 2171–2467 98 | Wed39-9 | ON186765 | 1360 | 189–545 118 (13) |
| Virus Name * | Isolate | Host Species | Family | Region/Country | GenBank Accession No. | Percent Nucleotide Sequence Identity |
|---|---|---|---|---|---|---|
| LUYVV | YN6153 | Ludwigia linifolia | Onagraceae | China | MN210345 | 74% |
| TbLCYnV | Y161 | tomato | Solanaceae | China | AJ566744 | 73.1% |
| TbLCYnV | Y143 | tobacco | Solanaceae | China | AJ512762 | 73.2% |
| TbLCYnV | Y3 | tobacco | Solanaceae | China | AF240674 | 73% |
| TbLCYnV | Y283 | Ageratum conyzoides | Asteraceae | China | AJ971267 | 73.2% |
| TbLCYnVCN | YN3095 | Ageratum conyzoides | Asteraceae | China | MN218668_ | 73.2% |
| TYLCShV | YN3383 | cowpea | Fabaceae | China | MT089918 | 73.1% |
| TYLCVNV | Vt-6 | tomato | Solanaceae | Vietnam | DQ641697 | 73.2% |
| AYVV | AFSP7d | Sauropus androgynus | Phyllanthaceae | Thailand | JN809824 | 73.3% |
| ToLCPatV | ToLCPatV | tomato | Solanaceae | India | KM383746 | 73.3% |
| PeLCV | Pk2 | Raphanus sativus | Brassicaceae | Pakistan | LT600729 | 72.5% |
| PeLCV | Mad-4 | soybean | Fabaceae | Pakistan | MN910265 | 72.2% |
| PeLCV | DP2 | papaya | Caricaceae | India | KX353622 | 75.5% |
| AlYVV | Jabalpur_A | Alternanthera sessilis | Amaranthaceae | India | LC316182 | 92.8% |
| AlYVV | Kangra | Synedrella sp. | Asteraceae | India | MG686552 | 92.6% |
| AlYVV | Rumex | Rumex nepalensis | Polygonaceae | India | LN795903 | 92.5% |
| AlYVV | Jabalpur_B | Alternanthera sessilis | Amaranthaceae | India | LC316183 | 92.5% |
| AlYVV | Mn5 | Eclipta prostrata | Asteraceae | Pakistan | KX906696 | 92.4% |
| AlYVV | Mn1 | Eclipta prostrata | Asteraceae | Pakistan | KX906695 | 91.6% |
| AlYVV | YN598 | Eclipta prostrata | Asteraceae | China | FJ015062 | 91.1% |
| AlYVV | G8 | Eclipta prostrata | Asteraceae | China | DQ375456 | 91.6% |
| AlYVV | F31 | Eclipta prostrata | Asteraceae | China | EU286798 | 91.1% |
| Virus | No of Seq | H | Hd | InDel Sites | S | Eta | Kt | π |
|---|---|---|---|---|---|---|---|---|
| AlYVOMV | 34 | 33 | 0.998 | 681 | 1309 | 2285 | 460 | 0.17 |
| ToLCB | 35 | 33 | 0.996 | 283 | 900 | 1200 | 138 | 0.12 |
| Virus Name | Isolate Name | Host Species | Family | Country or Region | Reference |
|---|---|---|---|---|---|
| Alternanthera yellow vein virus | AlYVV | Alternanthera sessilis | Amaranthaceae | India | [22] |
| Alternanthera yellow vein virus | AlYVV | Picrorhiza kurroa | Plantaginaceae | India | [24] |
| Alternanthera yellow vein virus | AlYVV | Eclipta prostrata | Asteraceae | Pakistan | [25] |
| Alternanthera yellow vein virus | AlYVV | Eclipta prostrata | Asteraceae | Pakistan | [26] |
| Alternanthera yellow vein virus | AlYVV | Eclipta prostrata | Asteraceae | Pakistan | [27] |
| Alternanthera yellow vein virus | AlYVV | Eclipta prostrata | Asteraceae | China | [28] |
| Alternanthera yellow vein virus | AlYVV | Sonchus arvensis | Asteraceae | Pakistan | [29] |
| Alternanthera yellow vein virus | AlYVV | Eclipta prostrata | Asteraceae | China | [30] |
| Alternanthera yellow vein virus | AlYVV | Eclipta prostrate (L.) | Compositae | China | [31] |
| Ludwigia yellow vein virus | LUYVV | Ludwigia hyssopifolia | Onagraceae | China | [32] |
| Alternanthera yellow vein virus | AlYVV | Alternanthera philoxeroides | Amaranthaceae | China | [33] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shafiq, M.; Ondrasek, G.; Al-Sadi, A.M.; Shahid, M.S. Molecular Signature of a Novel Alternanthera Yellow Vein Virus Variant Infecting the Ageratum conyzoides Weed in Oman. Viruses 2023, 15, 2381. https://doi.org/10.3390/v15122381
Shafiq M, Ondrasek G, Al-Sadi AM, Shahid MS. Molecular Signature of a Novel Alternanthera Yellow Vein Virus Variant Infecting the Ageratum conyzoides Weed in Oman. Viruses. 2023; 15(12):2381. https://doi.org/10.3390/v15122381
Chicago/Turabian StyleShafiq, Muhammad, Gabrijel Ondrasek, Abdullah Mohammed Al-Sadi, and Muhammad Shafiq Shahid. 2023. "Molecular Signature of a Novel Alternanthera Yellow Vein Virus Variant Infecting the Ageratum conyzoides Weed in Oman" Viruses 15, no. 12: 2381. https://doi.org/10.3390/v15122381
APA StyleShafiq, M., Ondrasek, G., Al-Sadi, A. M., & Shahid, M. S. (2023). Molecular Signature of a Novel Alternanthera Yellow Vein Virus Variant Infecting the Ageratum conyzoides Weed in Oman. Viruses, 15(12), 2381. https://doi.org/10.3390/v15122381