1. Introduction
Foot-and-mouth disease (FMD) is a highly contagious disease caused by the foot-and-mouth disease virus (FMDV). This disease is economically important and affects domestic livestock with cloven hooves worldwide [
1]. FMD control relies on two major strategies: culling of infected animals and vaccination [
2]. FMDV is a member of the
Picornaviridae family, which consists of both animal and human pathogens, such as enterovirus, rhinovirus, and poliovirus. The FMDV genome is single-stranded RNA with positive polarity. Viral RNA synthesis during replication and transcription is achieved by RNA-dependent RNA polymerase (RdRp) encoded by 3D, so-called 3D
pol [
3]. The 3D
pol of picornaviruses including FMDV and the RdRp of other RNA viruses share a common characteristic [
4,
5]. The three-dimensional structure of RdRp resembles a cupped right-handed configuration composed of three subdomains, the so-called thumb, fingers, and palm. The finger subdomain has extensions, termed fingertips, that bridge between the finger and the thumb subdomains to maintain the RdRp active site rearrangement and form the NTP entry site [
6]. Therefore, the topology of these subdomains is essential for RdRp’s catalytic activity during negative-stranded RNA synthesis by holding the RNA template in place, engaging NTPs, and promoting polymerization. The central region of the 3D
pol contains six conserved motifs labelled A, B, C, D, E, and F, which are located mostly in the palm subdomain and form the template-binding channel with the 3D
pol active site [
6]. Inhibition of 3D
pol activity can prevent viral genome replication. Therefore, because of its critical role in viral replication, 3D
pol is an attractive target for antiviral drug development.
As FMDV is highly contagious, broad-spectrum antiviral drugs as well as other novel natural and synthesized compounds might support the outbreak control by reducing spillover of the spreading virus into the environment. For example, the nucleoside analogue ribavirin (1-β-d-ribofuranosyl-1,2,4-triazole-3-carboxamide) is an antiviral drug licensed for the treatment of human and animal viral infections. However, ribavirin resistance caused by mutations within the RdRp coding sequences have been reported in many RNA viruses. For instance, poliovirus (PV) has substitution mutations, G64S and L420A, in 3D
pol while M296I was found in the FMDV 3D
pol [
7,
8]. Furthermore, a human hepatitis C virus (HCV) escape mutant contained the mutation Y415 in the NS5B gene [
9]. M296 of FMDV 3D
pol is in the NTP binding site. Thus, alteration at this position may affect nucleotide recognition by the 3D
pol [
10]. Previously, nucleoside analogues including favipiravir (T-705), T-1105 (3-oxo-3,4-dihydro-2-pyrazinecarboxamide derivative), and T-1106 were tested for their ability to inhibit FMDV infection in vitro and in vivo [
11]. T-1105 demonstrated greater potency than T-705 and T-1106 to inhibit FMDV replication [
11,
12]. In an experimental FMDV infection, a high dose of T-1105 at 200 mg/kg twice daily for seven consecutive days was required to prevent FMD clinical signs and reduce viral shedding [
12]. In a guinea pig model, the prophylactic effect of T-1105 was achieved at a high dose of 400 mg/kg/day twice daily for five consecutive days to give a similar protective level as the full-dose vaccination [
13]. Thus far, no antiviral agent has been approved for the treatment or prevention of FMDV infection.
Among antiviral agents, nucleoside-based inhibitors (NIs) are likely to exhibit off-target effects and reduction in their potency in clinical cases [
14]. Small molecules that target surface cavities or allosteric site of enzymes, so-called non-nucleoside inhibitors (NNIs), can be either natural or non-natural compounds [
15]. Their structures are more flexible and thus can fit in the nucleotide-binding pocket better than the NIs [
16]. Several NNI compounds have been reported to inhibit the RdRp activities of RNA viruses such as GPC-N114 in picornaviruses [
14], lycorine in MERS-CoV [
17], and NITD29 in Zika virus [
18]. In addition to RdRp, 3C proteases (3C
pro) are another attractive target for antiviral drug design as it is present and conserved across several positive-sense, single-stranded RNA viruses with picornavirus-like 3C
pro supercluster. It has been shown previously that three dipeptidyl-based synthetic molecules with different interactive chemical groups including an aldehyde (GC373), a bisulfite adduct (GC376), and an α-ketoamide (GC375) possessed broad-spectrum, potent antiviral effects [
19]. These compounds effectively inhibited protease (3C
pro or 3CL
pro) activities and replication of viruses from the families
Caliciviridae,
Coronaviridae, and
Picornaviridae, as determined using FRET protease assay and cell-based assays, respectively.
In this study, we screened small molecules that targeted FMDV 3D
pol by integrating computer-aided virtual screening and a cell-based antiviral assay to filter for potential antiviral compounds. We further determined the inhibitory activity of the selected compounds on FMDV 3D
pol using a recently established FMDV minigenome with the green fluorescence reporter protein in the cell-based 3D
pol inhibition assay [
20]. Taken together, we have identified four 3D
pol inhibitors and demonstrated the mechanism by which the small molecules could inhibit viral replication in FMDV-infected BHK-21 cells.
2. Materials and Methods
2.1. Virtual Screening of Small Molecules
The crystal structure of FMDV 3D
pol (FMDV RdRp) deposited in the PDB under a code 1wne.pdb was retrieved (
https://www.rcsb.org/structure/1WNE). Upon the 3D structure’s preparation for the virtual screening, we found that four amino acids (F34, A68, E144, and K148) on the protein structures differed from the deduced amino acid sequence of FMDV O189. Then, homology modeling based on the 3D
pol crystal structure (PDB: 1wne.pdb) was performed on the SWISS-MODEL server (
https://swissmodel.expasy.org/, accessed on 19 April 2021 [
20]) to prepare the complete protein structure of O189 3D
pol previously used for the plasmid construction [
20]. The structure quality was evaluated using MolProbity [
21], Q-MEAN [
22], and Ramachanadran plotting [
23], as described elsewhere [
20]. The FMDV 3D
pol model was prepared for the downstream process with the removal of ligands and water molecules and addition of hydrogen atoms. Molecular docking was achieved using the PyRx 0.9.8 virtual screening tool [
24].
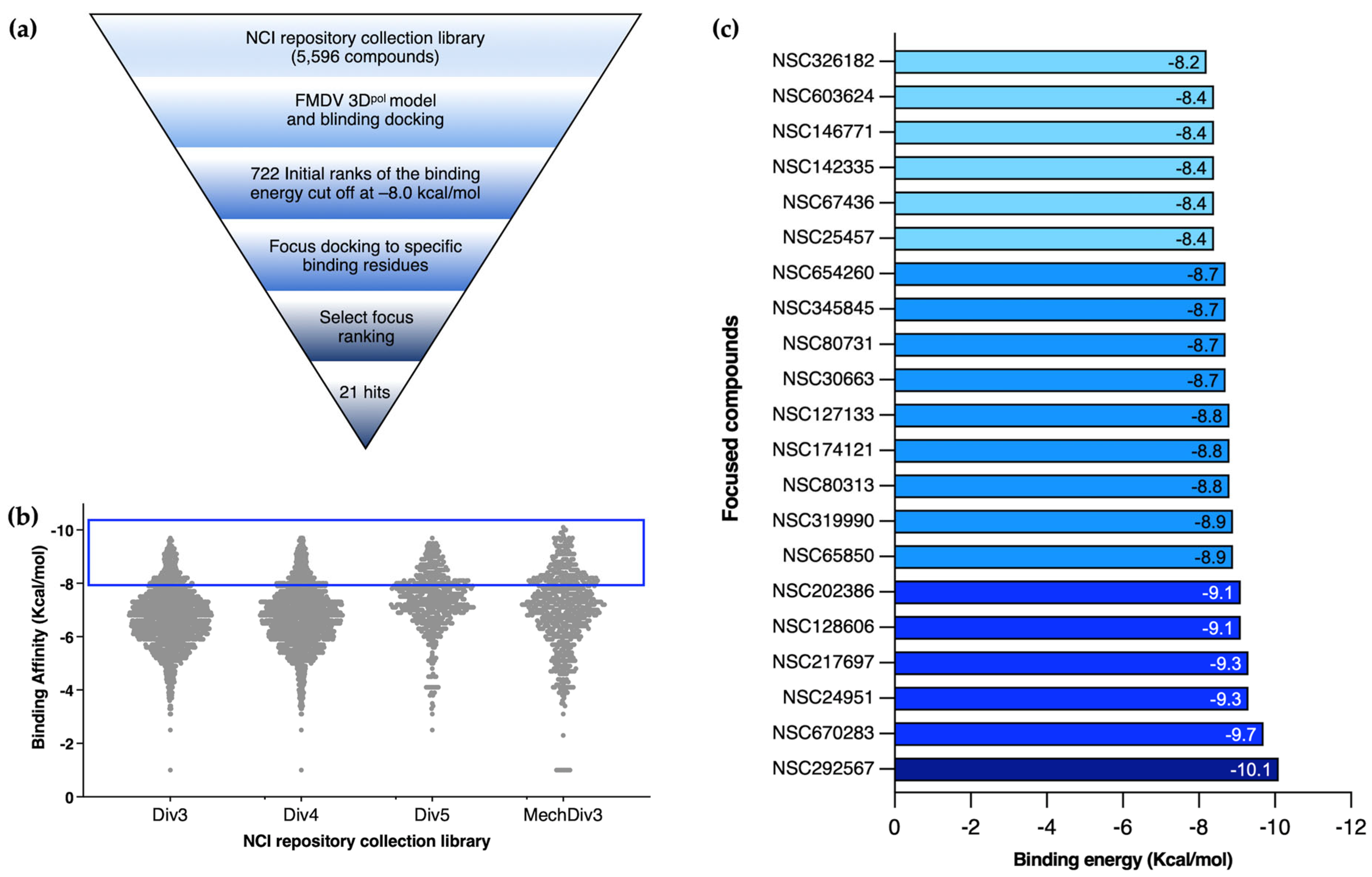
The compound libraries provided by the Developmental Therapeutics Program (DTP) Open Repository of the National Cancer Institute (NCI) were queried for a subset of the NCI repository collection comprising the NCI Diversity Set III, VI and V, and MechDiv3 libraries containing 5596 freely available models of compounds. Each subset of the compound models was assigned for docking using the Open Babel software toolbox [
25], including Gasteiger partial charges, addition of hydrogen atoms, optimization of hydrogen bonds, and removal of atomic clashes, to generate the pdbqt input format.
Virtual screening was performed using AutoDock Vina [
26] embedded in the PyRx 0.9.8 virtual screening tool [
24]. In the first virtual screening, the small molecules retrieved from the model libraries were blind-docked onto the 3D
pol structure, which yielded a set of ligands predicted to bind the 3D
pol active sites or the conserved amino acid residues important for 3D
pol’s function. Initially, the grid box was set at 65:70:65 (x, y, z dimensions in Angstroms), and the box was centered at 19.34:31.20:23.25 (x, y, z). The virtual screening outputs were presented as the predicted free energy in kcal/mol and the exhaustiveness value for the docking was set to 20. Subsequently, the top-ranked complexes were retrieved for the latter focus-docking. In the focus-docking, the top-ranked compounds with a binding energy lower than −8.0 kcal/mol were included, and the dockings were centered at specific sites or amino acid residues within the FMDV 3D
pol. Three sites on the palm subdomain of FMDV 3D
pol involved in the nucleotide binding and polymerizations [
6] were selected for the focus-docking. According to the 3D structure of FMDV 3D
pol, site 1 is composed of D240, Y241, and D245 of Motif A in the loop β8-α9; site 2 comprises M296, S298, G299, S301, and N307 of Motif B in the loop β9-α11; and site 3 contains Y336, D338, and D339 of Motif C in the loop β10-β11. The grid box of 25:25:25 was assigned to these specific sites and centered at 15.50:26.22:15.00 (x, y, z). The promising docking results were harvested for protein–ligand visualization using Discovery Studio Visualizer, version 2021 (BIOVIA, Dassault Systèmes, San Diego, CA, USA) and UCSF Chimera, version 1.16 (UCSF, San Francisco, CA, USA).
2.2. Sources and Preparations of Chemicals
The freely available compounds were provided by the Developmental Therapeutics Program (DTP) Open Repository of the National Cancer Institute (NCI). Ribavirin (Sigma-Aldrich, St. Louis, MO, USA) and rupintrivir (Sigma-Aldrich, St. Louis, MO, US) were used as RdRp and non-RdRp inhibitor controls. All compounds were prepared as 10-mM stock solutions in 100% dimethyl sulfoxide (DMSO) (Sigma-Aldrich, St. Louis, MO, USA), and stored at −20 °C for the following in vitro cell-based experiments.
2.3. Cells and Viruses
Baby Hamster Kidney (BHK−21) cells and HEK-293T cells were obtained from American Type Culture Collection (ATCC®, Manassas, VA, USA). The cells were maintained in a complete medium containing Minimum Essential Medium (MEM, InvitrogenTM, Carlsbad, CA, USA), 10% fetal bovine serum (FBS, InvitrogenTM, Carlsbad, CA, USA), 2 mM L-glutamine (InvitrogenTM, Carlsbad, CA, USA), and 1×Antibiotic-Antimycotic (InvitrogenTM, Carlsbad, CA, USA) at 37 °C with 5% CO2. FMDV serotype A (NP05) was propagated in BHK21 cells at 37 °C with 5% CO2 for 24 h. The virus stock with a titer of 1 × 109 TCID50/mL was stored at −80 °C in single-use aliquots. All works involving live FMDV were performed at biosafety level-2 with an enhanced facility.
2.4. Cytotoxicity Assay
BHK-21 cells were seeded at 1.8 × 10
4 cells per well into 96-well plates (Corning Incorporated., Corning, NY, USA) and incubated at 37 °C with 5% CO
2 overnight. The spent media was removed, and the cells were washed twice with 1× PBS. The compounds were serially diluted in serum-free MEM media containing DMSO at a final concentration of ≤0.1%. The diluted compounds were incubated with the cells or virus and the cells were further incubated at 37 °C for an additional 24 h. Two independent experiments were performed for all biological assays. Cytotoxicity was determined by measuring cell viability using MTS solution provided in the CellTiter 96
® Aqueous One Solution Cell Proliferation Assay (Promega, Madison, WI, USA) according to the manufacturer’s instructions. The absorbance of the solution in the experiment plates was measured at a wavelength of 490 nm using a multi-mode reader (Synergy H1 Hybrid Multi-Mode Reader, BioTek
®, Winooski, VT, USA). Cell viability was calculated using the following formula (1), where OD
treate denotes the absorbance of the uninfected BHK-21 cell wells containing cells, media, and compounds; OD
cell control denotes the absorbance of the cell control wells containing cells and media; and OD
dmso denotes the absorbance of the vehicle control wells containing cells, media, and 0.1% DMSO.
2.5. Antiviral Activity Assay
The compounds that targeted FMDV 3D
pol in the virtual screening and could inhibit viral replication were further investigated for the effects on the other processes of viral infection as mentioned in the previous study [
27]. Initially, we investigated the effects of compounds on viral entry including binding and penetration (pre-viral-entry experiment). The compound dilutions and FMDV at 10 TCID50 per wells were co-adsorbed onto the overnight-grown BHK-21 cells in a 96-well plate (Corning Incorporated., Corning, NY, USA) at 37 °C for 2 h. All virus–compound mixtures were removed, and the cells were washed twice with 1× PBS. Subsequently, fresh media was added to the cells which were further cultured at 37 °C for an additional 22 h. Secondly, in the post-viral-entry experiment, the BHK-21 cells in the 96-well plate were incubated with 10 TCID50 of FMDV per well at 37 °C for 2 h. Then, the cells were washed twice with 1× PBS, before treatment with the same compound dilutions that were used in the cytotoxicity assay at 37 °C for an additional 22 h to inhibit viral replication.
2.6. Immunoperoxidase Monolayer Assay (IPMA)
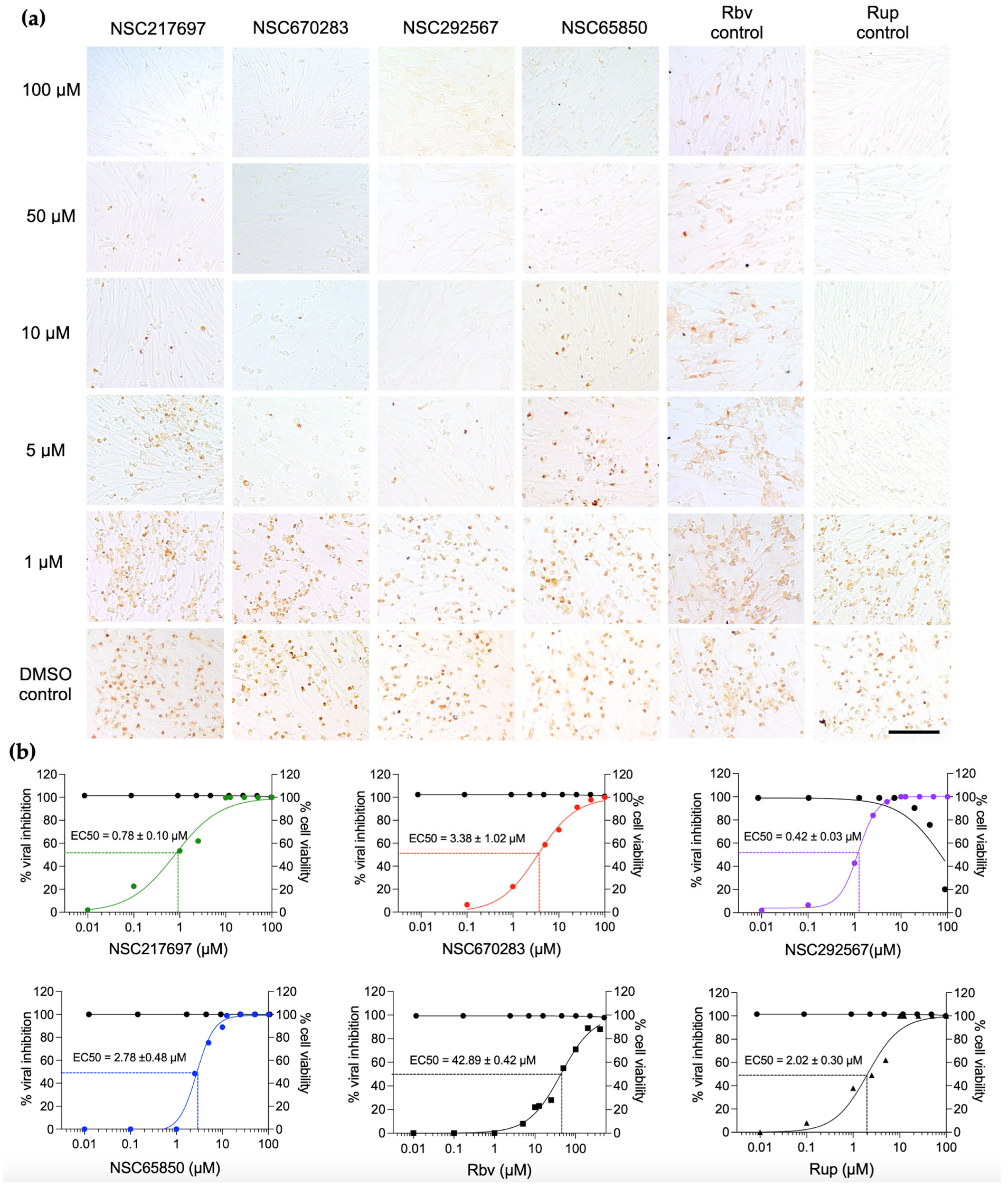
Immunoperoxidase monolayer assays (IPMA) were conducted as previously described [
27,
28]. Briefly, the BHK-21 cells, including no infection with and without compound and FMDV infection with and without compound, were fixed with cold methanol at room temperature for 20 min and washed with PBS plus 0.1% Tween 20 (1× PBST, Sigma Aldrich
®, St. Louis, MO, USA). The cells were then incubated with single-chain variable fragment with Fc fusion protein (scFv-Fc) specific to 3ABC of FMDV [
29] at 37 °C for 1 h for viral detection. After primary antibody incubation, the cells were washed with 1× PBST, and subsequently incubated with the protein G, HRP conjugate (dilution 1:1000, EMD Millipore corporation, Temecula, CA, USA) at 37 °C for 1 h. To visualize the antigen–antibody complex, the cells were stained with DAB substrate (DAKO, Santa Clara, CA, USA), and the dark-brown color of the infected FMDV cells was observed using a phase-contrast inverted microscope (Olympus IX73, Tokyo, Japan). The cell images were analyzed using CellProfiler image analysis v.4.2.0 [
29]. The resulting data were used to calculate the half-maximal effective concentration (EC50). The EC50 value represented the compound concentration at which the virus infection was reduced by 50% compared to the DMSO control (FMDV infection with 0.1% DMSO). To analyze the data, the DMSO control was set at 100% infection, and the EC50 value of each compound was calculated using GraphPad Software version 9.4.1 (Prism, San Diego, CA, USA). The Z’ factor value was analyzed using the following formula (2) to assess the assay performances both within a plate and across plates.
A Z’ factor between 0.5 and 1 indicates an acceptable range.
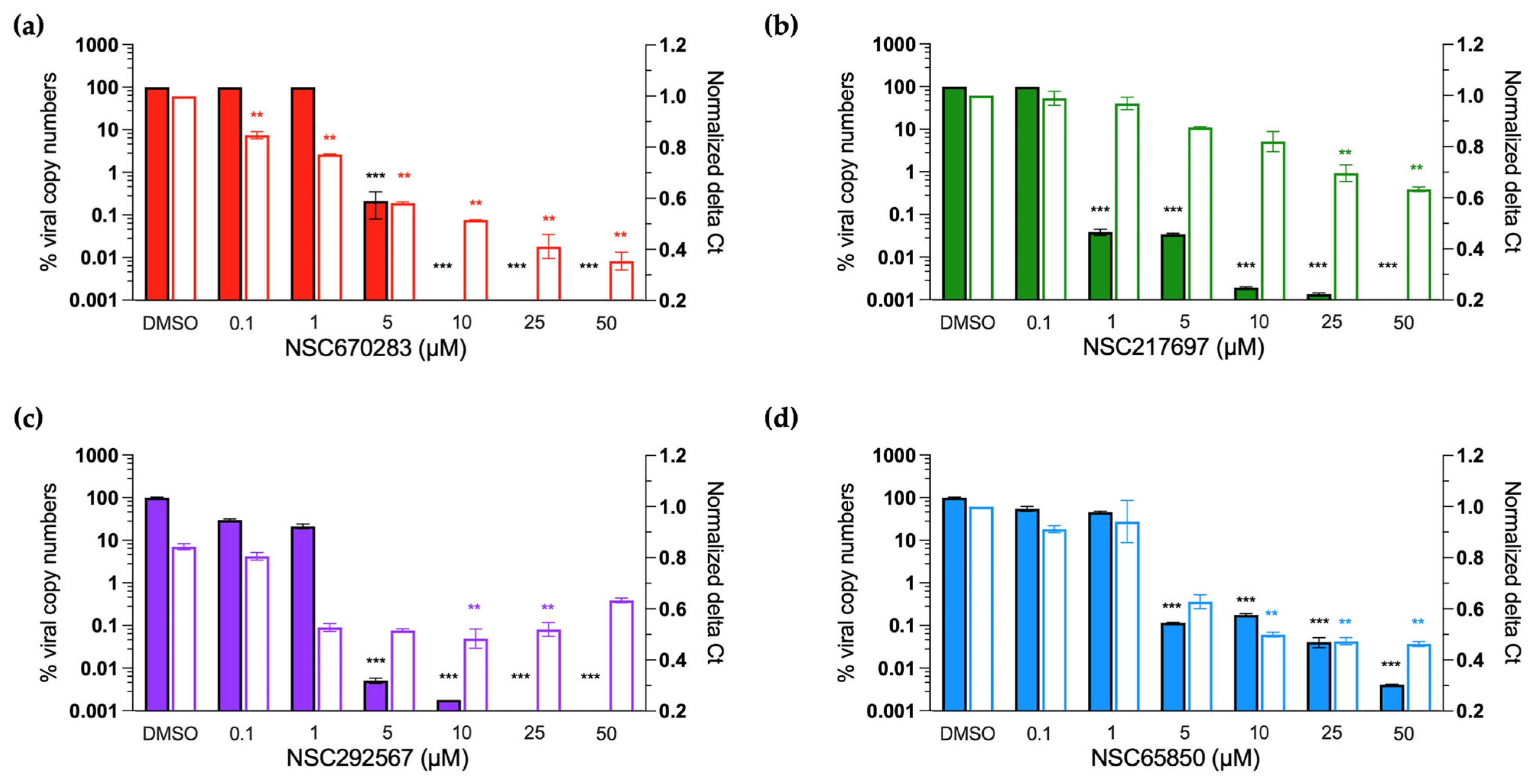
2.7. Real-Time RT-PCR Assay
The whole viral RNA and negative-strand specific RNA of FMDV were detected and quantitated using real-time RT-PCR after treatment with the compounds. The BHK-21 cells were seeded at 1.8 × 10
5 cells into each well of 24-well plates (Corning Incorporated., Corning, NY, USA) and cultured overnight. The cells were incubated with 10 TCID50 of FMDV for 2 h, and the inoculum was removed after viral adsorption. Then, the FMDV-infected cells were treated with serially diluted compounds at 37 °C as mentioned above. At 24 h post viral infection, intra- and extracellular viral RNA was isolated using Direct-zol
TM RNA MiniPrep (Zymo Research Corporation, Tustin, CA, USA) following the manufacturer’s instruction. The RNA was quantified using a NanoDrop
TM 2000c Spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA) and used as the templates for cDNA synthesis. The first-strand cDNAs derived from the whole viral RNA were generated using Random hexamers (Invitrogen
TM, Carlsbad, CA, USA) for whole viral RNA quantification. On the other hand, a primer specific to the negative-stranded RNA within the 3D
pol coding sequence (5′-AAGGGTTGATTGTTGACA-3′) was employed to amplify the first-strand cDNA for the negative-stranded RNA quantification [
30]. Both cDNA syntheses were performed with the enzyme SuperScript III Reverse Transcriptase (Invitrogen
TM, Carlsbad, CA, USA) according to the manufacturer’s instructions.
The cDNA templates were subjected to downstream DNA quantification with qPCR using iTaq Universal SYBR Green Supermix (Bio-Rad Laboratories, Hercules, CA, USA). The primer sets for the DNA amplifications are listed in
Table 1. Briefly, 5 µL mixture containing 0.5 µL of each target-specific forward and reverse primer was mixed with 2 µL cDNA and nuclease-free ddH
2O up to a final volume of 10 µL. The qPCR amplifications were conducted in a two-step method: first, denaturation at 95 °C for 30 sec, followed by 40 cycles of denaturation at 95 °C for 5 sec and annealing/extension at 55 °C for 30 sec. A melting curve was analyzed from 65 °C to 95 °C with 0.5 °C increments using a CFX96 touch Real-Time PCR detection system (Bio-Rad Laboratories, Hercules, CA, USA).
Viral RNA expression was determined using the absolute quantification method [
27]. A standard curve of DNA derived from the whole viral RNA was generated from the ten-fold serially diluted plasmid containing FMDV 5′UTR from 10
−2 to 10
−7 plasmid molecules/µL. Copy numbers of the DNA were calculated based on the standard curve. The negative-stranded RNA is normally produced by 3D
pol during viral replication. The negative-stranded RNA-derived DNA was quantitated based on the delta Ct values (cycle threshold) by subtracting the Ct values of the virus samples (FMDV-infected BHK-21 cells with compound treatment) from the Ct values of the virus control (FMDV-infected BHK-21 cells without compound treatment). The data were normalized using the following Equation (3).
The normalized data from three independent replications are presented as the means ± SD using GraphPad Prism version 9.4.1 (Prism, San Diego, CA, USA).
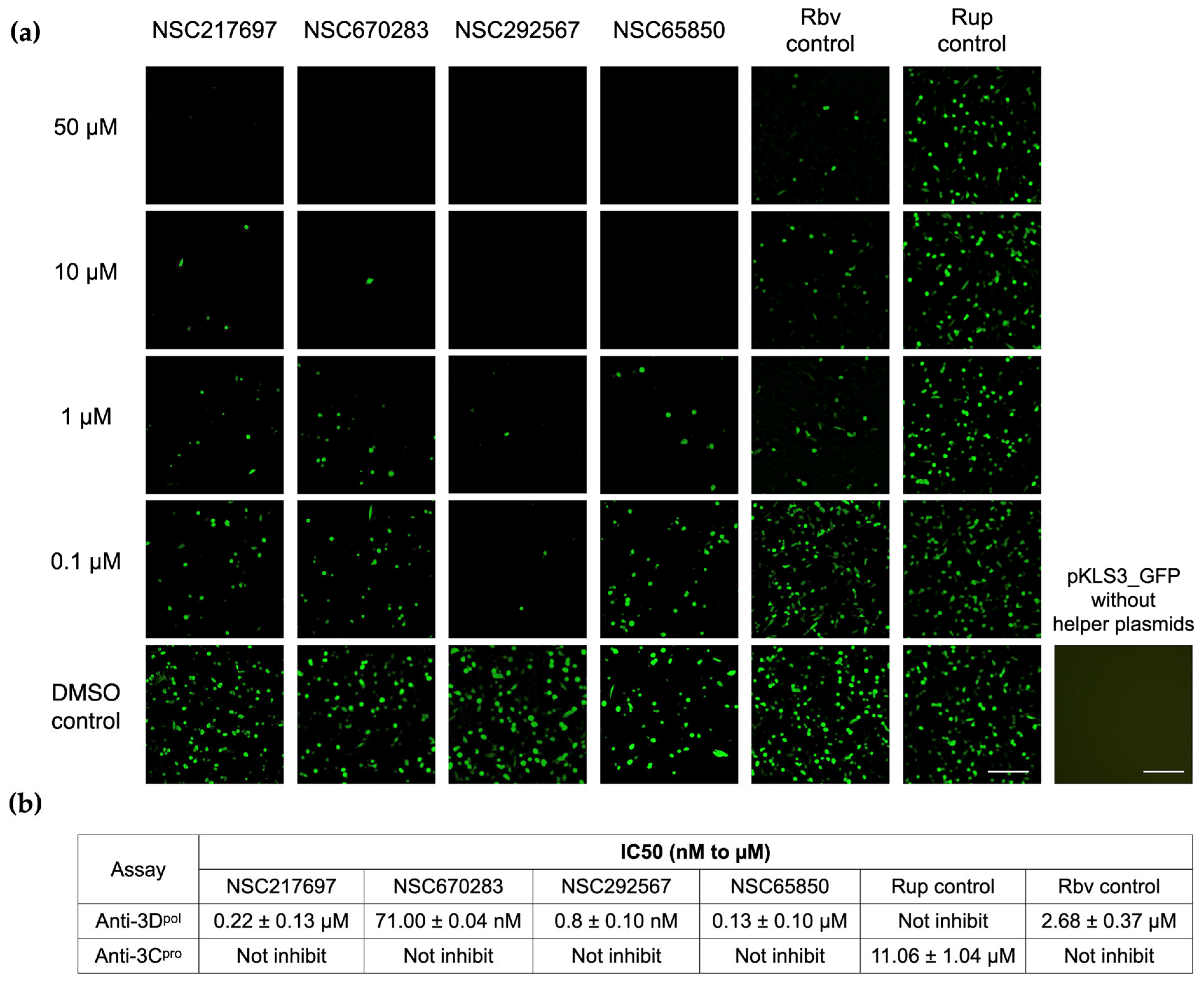
2.8. Cell-Based 3Dpol Inhibition Assay
Inhibitory effects of the compounds on FMDV 3D
pol were examined using plasmid pKLS3_GFP, an FMDV minigenome expressing GFP, and the two helper plasmids pCAGGS_T7 and pCAGGS_P3 [
20]. pKLS3_GFP contains the enhanced green fluorescent protein (GFP) gene inserted between FMDV O189 5′ and 3′UTRs required for FMDV transcription and translation, while pCAGGS_T7 and pCAGGS_P3 are mammalian protein expression plasmids containing T7 RNA polymerase and the FMDV P3 region essential for efficient generation of FMDV RNA, respectively.
The three plasmids were transfected onto the BHK-21 cells as described previously [
20]. Briefly, BHK-21 cells were seeded at 1 × 10
4 cells/well into 96-well plates and incubated at 37 °C overnight. On the transfection day, 40 ng of pKLS3_GFP, 120 ng of pCAGGS_T7, and 40 ng of pCAGGS_P3 were mixed with 0.6 µL of Fugene
® HD (Promega, Madison, WI, USA) in Opti-MEM™ I Reduced-Serum Medium (Thermo Fisher Scientific, Waltham, MA, USA) to make a final volume of 50 µL per well. Then, the spent media was removed and replaced with the transfection mixture and incubated at 37 °C for 4 h. Subsequently, the transfection mixture was removed and replaced with the serially diluted compounds in Opti-MEM
TM I Reduced-Serum Medium (Gibco
TM Thermo Fisher Scientific Inc., Waltham, MA, USA). In the vehicle control well, 0.1% DMSO was added in the wells instead of the compounds and it also served as the transfection positive control. pKLS3_GFP without helper plasmid transfection was also included as the plasmid control. The cells were incubated at 37 °C with 5% CO
2 and the fluorescent reporter expression signals were observed at 24–48 h post transfection using a phase-contrast inverted microscope (Olympus IX73, Tokyo, Japan).
The levels of FMDV 3D
pol activity corresponded to the numbers and intensity of the bright green fluorescent signal in the positive cells. The background was adjusted for contrast and brightness only. The fluorescent intensities of the images were measured, and the background was subtracted from each image using CellProfiler image analysis v.4.2.0 [
29]. The half-maximal inhibitory concentration (IC50) is the concentration at which the 3D
pol activity was reduced by 50% compared to that of the DMSO control. For the analysis, the signal of the DMSO control was set at 100% and the IC50 was calculated using GraphPad Software version 9.4.1 (Prism, San Diego, CA, USA).
2.9. Cell-Based FMDV 3Cpro Inhibition Assay
3C
pro is the main protease crucial for FMDV biology and is an attractive antiviral target. Therefore, we were interested to know whether these compounds could inhibit the 3C
pro activity. We determined the effects of the compounds on the main protease using a cell-based FMDV 3C
pro inhibition assay as described previously [
27,
31]. Briefly, HEK-293T cells were grown in 96-well plates at 1 × 10
3 cells per well. The cells were maintained in Opti-MEM I Reduced-Serum Medium (Gibco
TM Thermo Fisher Scientific Inc., Waltham, MA, USA). On the next day, the cells were transfected with plasmids pG5Luc (Promega, Madison, WI, USA) and pBV_3ABCD expressing intact 3C
pro or pBV_mu3ABCD expressing inactive 3C
pro [
32]. The 3ABCD and mu3ABCD genes were inserted between the Gal4-binding domain and the VP16-activation domain of plasmid pBV (Promega, Madison, WI, USA). pG5Luc is a reporter plasmid containing the GAL4 binding site upstream of VP16 and the firefly luciferase reporter gene sequences followed by the downstream
Renilla luciferase gene. The total 10 µL of co-transfection mixtures comprising 0.1 µg pBV_3ABCD or pBV_mu3ABCD, 0.1 µg pG5Luc, and 0.6 µL Fugene
® HD (Promega, Madison, WI, USA) were incubated with HEK-293 T cells at 37 °C for 2 h before adding the diluted compounds. At 16 h post transfection, the transfected cells were lysed with 20 µL passive lysis buffer (Promega, Madison, WI, USA) followed by firefly and
Renilla luminescence signal determination using the Dual-Glo Luciferase Assay System (Promega, Madison, WI, USA) in a Synergy H1 Hybrid Multi-Mode Microplate Reader (BioTek, Winooski, VT, USA). The intact 3C
pro expressed from pBV_3ABCD would separate the Gal4-binding domain from the VP16-activation domain, resulting in no expression of the firefly luciferase signal. When the compounds could suppress the 3C
pro, the Gal4-binding domain and the VP16-activation domain were in close proximity. Thus, GAL4 bound to the Gal4-binding domain and drove the firefly and
Renilla luciferase expressions. The data were recorded as an inverse correlation of the firefly/
Renilla luminescent (Fluc/Rluc) signal ratio from compound-treated wells compared to those obtained from wells transfected with pBV_mu3ABCD (3C
pro negative control) and pBV16 (empty plasmid control).
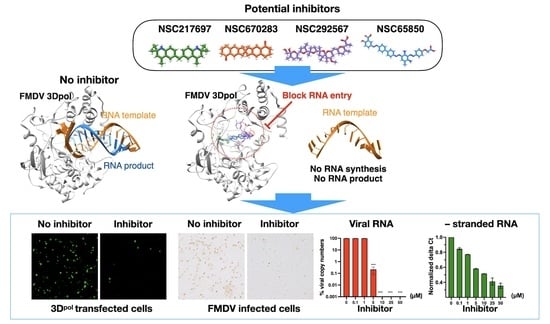
4. Discussion
RNA-dependent RNA polymerase (RdRp) is the enzyme essential for replication and transcription processes of RNA viruses including FMDV and other picornaviruses. Therefore, RdRp is an attractive target for novel inhibitors against RNA viruses. In the case of FMDV, the crystal structures of RdRp (3D
pol) have been resolved, and are similar to the right-handed architecture with three conserved subdomains—palm, finger, and thumb. This characteristic is common among picornaviruses [
5,
6]. Computational drug discovery has proven to be a guide for screening new drug candidates, and this method can directly provide scientific evidence for further in vitro and in vivo testing. In this study, we optimized the FMDV 3D
pol model for small-molecule screening processes. Initially, we performed the blind virtual screening of small molecules to filter the 3D
pol binding compounds for the second-round focused docking. The focused molecular docking specified the compounds that fit the active sites and involved functional areas of FMDV 3D
pol. We demonstrated that four out of 21 compounds obtained from the double virtual screening inhibited FMDV at the post-infection stage using a cell-based antiviral assay. Moreover, these candidates showed good inhibition specific to FMDV 3D
pol activity, but not 3C
pro. 3D
pol is one of the most important enzymes for viral replication processes and it functions after viral infection. In our study, ribavirin could inhibit FMDV 3D
pol as it is a well-known broad-spectrum anti-RNA virus and the complex of FMDV RdRp, RNA template, and ribavirin triphosphate has been solved and demonstrated [
10]. In addition, a ribavirin-resistant mutant, which contained M296I on the β9-α11 loop adjacent to the active site, induced misincorporation of guanosine monophosphate into the RNA chain leading to an error catastrophe [
8,
10,
33]. Therefore, it is suitable to be used as a positive RdRp inhibitor.
We have shown that the four compounds, NSC217697, NSC670283, NSC292567 and NSC65850, interacted with the active sites of FMDV 3D
pol to inhibit the enzyme function, and thus markedly reduced synthesis of viral RNA. NSC217697 (6,6′-methylene bis(2,2,4-trimethyl-1,2-dihydroquinoline)) is a quinoline, which contains fused rings of heterocyclic compounds. We found that this compound could inhibit FMDV replication after infection. Moreover, NSC217697 could inhibit 3D
pol with a micromolar concentration, but had no inhibitory effect on FMDV 3C
pro. This compound was predicted to bind the NTP binding residues (R168, K172, and R179) in the finger subdomain, which possibly masked an initiation site in the complex conformation. Recently, quinoline and quinazoline derivatives have been shown to inhibit SARS-CoV-2 RdRp [
34]. We assumed that NSC217697 could be an allosteric inhibitor of FMDV 3D
pol.
NSC670283 (2,2′-spirobi [3,6,7,8-tetrahydro-1H-cyclopenta[g]naphthalene]-5,5′-dione) is a spiro compound that contains a fused unit of 1,2,3,6,7,8-hexahydro-5
H-cyclopenta[
b]naphthalen-5-one at position 2. This spiro compound has been reported to predictably interact with hepatitis C virus E2 envelope glycoprotein (HCV E2). It could bind HCV E2 protein as detected by surface plasmon resonance [
35]. According to a viral specificity test, NSC670283 also inhibited RD114 glycoprotein (RD114pp) of endogenous feline retrovirus with IC50 of 3 µM in Huh-7 cells. However, antiviral activity against HCV replication in cell culture was not determined in the previous study. Indeed, we have shown that NSC670283 reacted to specific amino acid residues within Motifs C and E and the catalytic aspartic residue in the palm subdomain of FMDV 3D
pol. This compound could inhibit FMDV replication as well as FMDV 3D
pol activity with micromolar IC50 and EC50, suggesting a potential broad-spectrum antiviral.
Among the four compounds, NSC292567 (nigericin or pandavir) demonstrated the highest binding affinity. This compound is grouped with the ionophore antibiotics, which have inhibitory activities against drug-resistant strains of Gram-positive bacteria and coccidian protozoa. It has been used to treat coccidiosis in poultry [
36]. During the global COVID-19 crisis, the ionophore antibiotics were a group of interesting drugs that were repurposed for antiviral drug development. For example, monesin, a monovalent cation/proton antiporter, was found to inhibit MERS-CoV [
37] and SARS-CoV-2 [
38]. Salinomymin could inhibit influenza A and B viruses by blocking endosomal acidification and interfering with viral matrix protein 2 [
39]. In addition, nigericin is an ionophore that affects lipid-soluble molecules. It increases the permeability of an ion across a biological membrane as well as the lipid bilayer of the vesicular transport system that functions in cellular trafficking of protein macromolecules. Picornavirus replication requires maturation of these membranous vesicles in the vesicular compartment during plus- and minus-strand RNA syntheses. Disruption of this membranous vesicle by ionophores thus inhibits viral replication [
40]. Nigericin demonstrated a moderate inhibition of SARS-CoV-2 [
38]. In this study, nigericin was ranked in the first place of our virtual screening with a binding affinity of −10.1 kcal/mol, and it had a good inhibitory effect on FMDV replication with an EC50 value of 0.42 µM. Moreover, nigericin at 0.8 nM reduced FMDV 3D
pol activity by 50%, and the anti-3D
pol action of this compound was confirmed by the reduction of negative-stranded RNA synthesis. In our predicted model study, nigericin could occupy the finger and palm subdomains of 3D
pol. These results suggest that the ionophore antibiotics could be potential antivirals against FMDV infection by directly suppressing RNA synthesis of 3D
pol.
NSC65850 (5-((4′-((2,4-diamino-3-((4-(carboxymethoxy)phenyl)diazenyl)-5-methylphenyl)diazenyl)[1,1′-biphenyl]-4-yl)diazenyl)-2-hydroxybenzoic acid) is a compound in the NCI diversity set III. Thus far, its antimicrobial effect has not been reported in the available databases. Therefore, we further investigated the physicochemical properties of this compound. The predicted model based on the SwissAMDE analysis [
41] (accessed on 1 August 2022) demonstrated that this compound possessed a poor solubility (Log
S = −11.83), Log
Po/w of 7.17, low skin permeation (log
Kp = −5.14 cm/s), and low GI absorption. However, its synthetic accessibility score was 4.25, which is moderate for chemical modification in medicinal chemistry to improve its physicochemical property for druggable agents. Nonetheless, our experiments in the cell-based assay showed that this compound could inhibit 3D
pol, but not 3C
pro of FMDV.


 ,
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}