
Atypical and Unique Transmission of Monkeypox Virus during the 2022 Outbreak: An Overview of the Current State of Knowledge


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
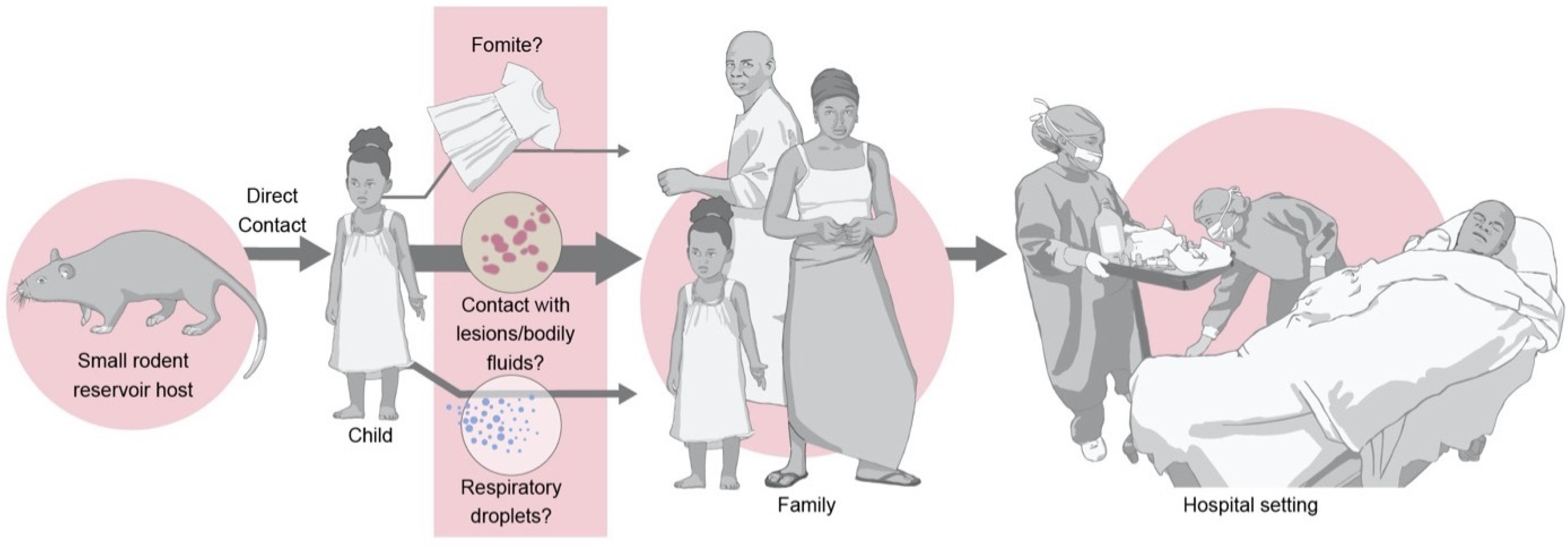
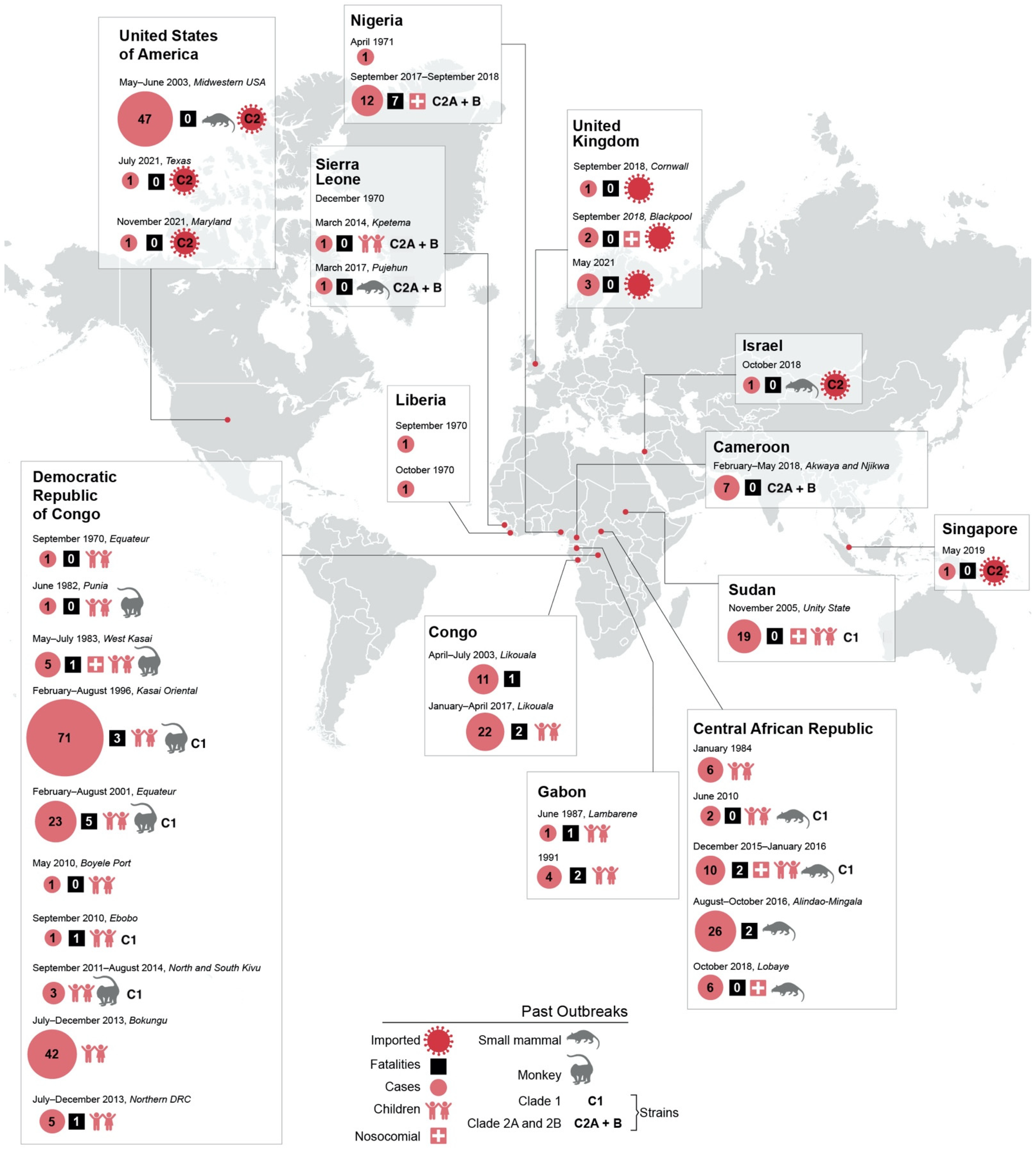
2. Ecology
3. Spillover
4. Human-To-Human Transmission
4.1. Epidemiological Dynamics
4.2. Routes
4.3. Determinants of Efficiency
4.4. Risk Factors
4.5. Countermeasures
5. Current Outbreak
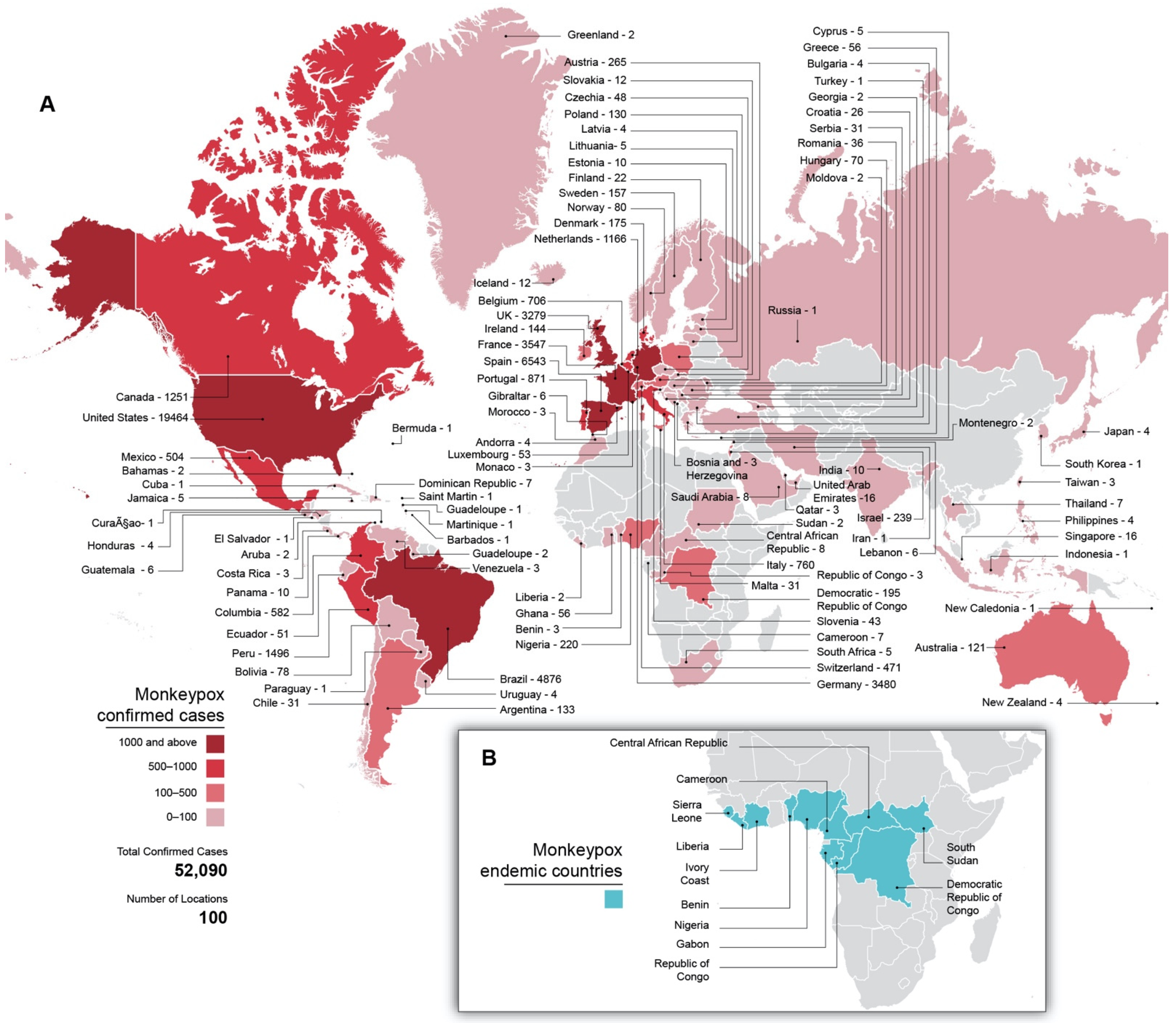
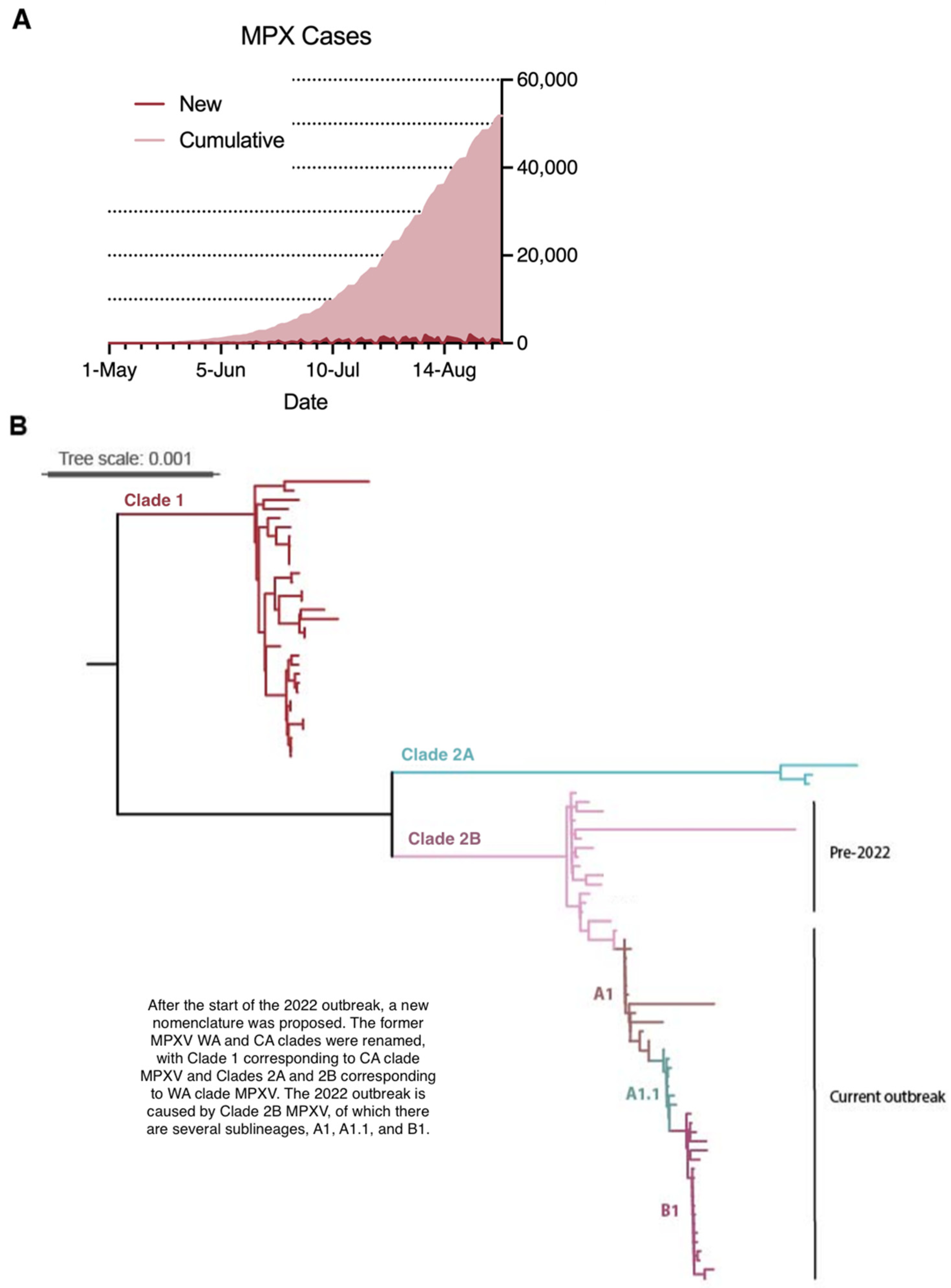
5.1. Epidemiology
5.2. Atypical Presentation
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Adalja, A.I.T. A Novel International Monkeypox Outbreak. Ann. Intern. Med. 2022, 175, 1175–1176. [Google Scholar] [CrossRef] [PubMed]
- CDC. Monkeypox and Orthopoxvirus Outbreak Global Map. Centers for Disease Control and Prevention. 2022. Available online: https://www.cdc.gov/poxvirus/monkeypox/response/2022/world-map.html (accessed on 1 September 2022).
- Essbauer, S.P.M.; Meyer, H. Zoonotic poxviruses. Vet. Microbiol. 2010, 140, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Brown, K.L.P. Human Monkeypox: Current State of Knowledge and Implications for the Future. Trop. Med. Infect. Dis. 2016, 1, 8. [Google Scholar] [CrossRef]
- Bunge, E.H.B.; Chen, L.; Lienert, F.; Weidenthaler, H.; Baer, L.; Steffen, R. The changing epidemiology of human monkeypox—A potential threat? A systematic review. PLoS Negl. Trop. Dis. 2022, 16, e0010141. [Google Scholar] [CrossRef]
- WHO. Multi-Country Monkeypox Outbreak in Non-Endemic Countries; World Health Organization: Geneva, Switzerland, 2022.
- IA, C.H.; Mbala, P.; Njouom, R.; Nakoune, E.; Happi, A.; Ndodo, N.; Ayansola, O.; Mboowa, G.; Bedford, T.; Neher, R.A.; et al. Urgent Need for a Non-Discriminatory and Non-Stigmatizing Nomenclature for Monkeypox Virus. 2022. Available online: https://virological.org/t/urgent-need-for-a-non-discriminatory-and-non-stigmatizing-nomenclature-for-monkeypox-virus/853/1 (accessed on 1 September 2022).
- Likos, A.M.; Sammons, S.A.; Olson, V.A.; Frace, A.M.; Li, Y.; Olsen-Rasmussen, M.; Davidson, W.; Galloway, R.; Khristova, M.L.; Reynolds, M.G.; et al. A tale of two clades: Monkeypox viruses. J. Gen. Virol. 2005, 86 Pt 10, 2661–2672. [Google Scholar] [CrossRef] [PubMed]
- Breman, J.R.K.; Steniowski, M.; Zanotta, E.; Gromyko, A.; Arita, I. Human monkeypox, 1970–1979. Bull. World Health Organ. 1980, 58, 165–182. [Google Scholar]
- Jezek, Z.M.S.; Mutumbo, M.; Nakano, J.; Paluku, K.; Szczeniowski, M. Human monkeypox: A study of 2510 contacts of 214 patients. J. Infect. Dis. 1986, 154, 551–555. [Google Scholar] [CrossRef]
- Hammarlund, E.L.M.; Carter, S.; Amanna, I.; Hansen, S.; Strelow, L.; Wong, S.; Yoshihara, P.; Hanifin, J.; Slifka, M. Multiple diagnostic techniques identify previously vaccinated individuals with protective immunity against monkeypox. Nat. Med. 2005, 11, 1005–1011. [Google Scholar] [CrossRef]
- Khodakevich, L.J.Z.; Kinzanzka, K. Isolation of monkeypox virus from wild squirrel infected in nature. Lancet 1986, 327, 98–99. [Google Scholar] [CrossRef]
- Hutin, Y.W.R.; Malfait, P.; Pebody, R.; Loparev, V.; Ropp, S.; Rodriguez, M.; Knight, J.; Tshioko, F.; Khan, A.; Szczeniowski, M.; et al. Outbreak of Human Monkeypox, Democratic Republic of Congo, 1996–1997. Emerg. Infect. Dis. 2001, 7, 434–438. [Google Scholar] [CrossRef]
- Doty, J.M.J.; Kalemba, L.; Stanley, W.; Monroe, B.; Nakazawa, Y.; Mauldin, M.; Bakambana, T.; Liyandja, T.; Braden, Z.; Wallace, R.; et al. Assessing Monkeypox Virus Prevalence in Small Mammals at the Human-Animal Interface in the Democratic Republic of the Congo. Viruses 2017, 9, 283. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, M.C.D.; Olson, V.; Hughes, C.; Galley, J.; Likos, A.; Montgomery, J.; Suu-Ire, R.; Kwasi, M.; Root, J.; Braden, Z.; et al. A Silent Enzootic of an Orthopoxvirus in Ghana, West Africa: Evidence for Multi-Species Involvement in the Absence of Widespread Human Disease. Am. J. Trop. Med. Hyg. 2010, 82, 746–754. [Google Scholar] [CrossRef] [PubMed]
- Fuller, T.T.H.; Mulembakani, P.; Johnston, S.; Lloyd-Smith, J.; Kisalu, N.; Lutete, T.; Blumberg, S.; Fair, J.; Wolfe, N.; Shongo, R.; et al. Using Remote Sensing to Map the Risk of Human Monkeypox Virus in the Congo Basin. EcoHealth 2010, 8, 14–25. [Google Scholar] [CrossRef] [PubMed]
- Thomassen, H.F.T.; Asefi-Najafabady, S.; Shiplacoff, J.; Mulembakani, P.; Blumberg, S.; Johnston, S.; Kisalu, N.; Kinkela, T.; Fair, J.; Wolfe, N.; et al. Pathogen-Host Associations and Predicted Range Shifts of Human Monkeypox in Response to Climate Change in Central Africa. PLoS ONE 2013, 8, e66071. [Google Scholar] [CrossRef]
- Reynolds, M.D.J.; McCollum, A.; Olson, V.; Nakazawa, Y. Monkeypox re-emergence in Africa: A call to expand the concept and practice of One Health. Expert Rev. Anti-Infect. Ther. 2019, 17, 129–139. [Google Scholar] [CrossRef]
- Yinka-Ogunleye, A.A.O.; Dalhat, M.; Ongoina, D.; Ogoina, D.; McCollum, A.; Disu, Y.; Mamadu, I.; Akinpelu, A.; Ahmad, A.; Burga, J.; et al. Outbreak of human monkeypox in Nigeria in 2017–18: A clinical and epidemiological report. Lancet Infect. Dis. 2019, 19, 872–879. [Google Scholar] [CrossRef]
- Jezek, Z.; Nakano, J.H.; Arita, I.; Mutombo, M.; Szczeniowski, M.; Dunn, C. Serological survey for human monkeypox infections in a selected population in Zaire. J. Trop. Med. Hyg. 1987, 90, 31–38. [Google Scholar]
- Doshi, R.G.S.; Doty, J.; Babeaux, A.; Matheny, A.; Burgado, J.; Townsend, M.; Morgan, C.; Satheshkumar, S.; Ndakala, N.; Kanjingankolo, T.; et al. Epidemiologic and Ecologic Investigations of Monkeypox, Likouala Department, Republic of the Congo, 2017. Emerg. Infect. Dis. 2019, 25, 273–281. [Google Scholar] [CrossRef]
- Falendysz, E.L.J.; Doty, J.; Nakazawa, Y.; Crill, C.; Lorenzsonn, F.; Kalemba, L.; Ronderos, M.; Mejia, A.; Malekani, J.; Karem, K.; et al. Characterization of Monkeypox virus infection in African rope squirrels (Funisciurus sp.). PLoS Negl. Trop. Dis. 2017, 11, e0005809. [Google Scholar] [CrossRef]
- Falendysz, E.L.J.; Lorenzsonn, F.; Salzer, J.; Hutson, C.; Doty, J.; Gallardo-Romero, N.; Carroll, D.; Osorio, J.; Rocke, T. Further Assessment of Monkeypox Virus Infection in Gambian Pouched Rats (Cricetomys gambianus) Using In Vivo Bioluminescent Imaging. PLoS Negl. Trop. Dis. 2015, 9, e0004130. [Google Scholar] [CrossRef]
- Guarner, J.J.B.; Paddock, C.; Shieh, W.; Goldsmith, C.; Reynolds, M.; Damon, I.; Regnery, R.; Zaki, S. Monkeypox Transmission and Pathogenesis in Prairie Dogs. Emerg. Infect. Dis. 2004, 10, 426–431. [Google Scholar] [CrossRef] [PubMed]
- Radonic, A.M.S.; Dabrowski, P.; Couacy-Hymann, E.; Schuenadel, L.; Kurth, A.; Matz-Rensing, K.; Boesch, C.; Leendertz, F.; Nitsch, A. Fatal Monkeypox in Wild-Living Sooty Mangabey, Cote d’Ivoire, 2012. Emerg. Infect. Dis. 2014, 20, 1009–1011. [Google Scholar] [CrossRef] [PubMed]
- Patrono, L.P.K.; Samuni, L.; Ulrich, M.; Rothemeier, C.; Sachse, A.; Muschter, S.; Nitsche, A.; Couacy-Hymann, E.; Boesch, C.; Wittig, R.; et al. Monkeypox virus emergence in wild chimpanzees reveals distinct clinical outcomes and viral diversity. Nat. Microbiol. 2020, 5, 955–965. [Google Scholar] [CrossRef]
- Alakunle, E.M.U.; Nchinda, G.; Okeke, M. Monkeypox Virus in Nigeria: Infection Biology, Epidemiology, and Evolution. Viruses 2020, 12, 1257. [Google Scholar] [CrossRef] [PubMed]
- Salzer, J.C.D.; Rwego, I.; Li, Y.; Falendysz, E.; Shisler, J.; Karem, K.; Damon, I.; Gillespie, T. Serologic evidence for circulating orthopoxviruses in peridomestic rodents from rural Uganda. J. Wildl. Dis. 2013, 49, 125–131. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Erez, N.A.H.; Milrot, E.; Schwartz, Y.; Wiener-Nell, Y.; Paran, N.; Politi, B.; Tamir, H.; Israely, T.; Weiss, S.; Beth-Din, A.; et al. Diagnosis of Imported Monkeypox, Israel, 2018. Emerg. Infect. Dis. 2019, 25, 980–983. [Google Scholar] [CrossRef] [PubMed]
- Berthet, N.N.E.; Whist, E.; Selekon, B.; Pharm, A.; Manugerra, J.; Gessain, A.; Kazanji, M. Maculopapular lesions in the Central African Republic. Lancet 2011, 378, 1354. [Google Scholar] [CrossRef]
- Besombes, C.G.E.; Konamna, X.; Selekon, B.; Gessain, A.; Berthet, N.; Manuguerra, J.; Fontanet, A.; Nakoune, E. Intrafamily Transmission of Monkeypox Virus, Central African Republic, 2018. Emerg. Infect. Dis. 2019, 25, 1602–1604. [Google Scholar] [CrossRef]
- Khodakevich, L.J.Z.; Messinger, D. Monkeypox virus: Ecology and public health significance. Bull. World Health Organ. 1988, 66, 747–752. [Google Scholar]
- McCollum, A.N.Y.; Ndongala, G.; Pukuta, E.; Pukuta, E.; Karhemere, S.; Lushima, R.S.; Ilunga, B.K.; Kabamba, J.; Wilkins, K.; Gao, J.; et al. Human Monkeypox in the Kivus, a Conflict Region of the Democratic Republic of the Congo. Am. J. Trop. Med. Hyg. 2015, 93, 718–721. [Google Scholar] [CrossRef]
- Meyer, H.P.M.; Stemmler, M.; Emmerich, P.; Schmitz, H.; Varaine, F.; Shungu, R.; Tshioko, F.; Formenty, P. Outbreaks of Disease Suspected of Being Due to Human Monkeypox Virus Infection in the Democratic Republic of Congo in 2001. J. Clin. Microbiol. 2002, 40, 2919–2921. [Google Scholar] [CrossRef] [PubMed]
- Reed, K.M.J.; Graham, M.; Regnery, R.; Sotir, M.; Wegner, M.; Kazmierczak, J.; Stratman, E.; Li, Y.; Fairley, J.; Swain, G.; et al. The detection of monkeypox in humans in the Western Hemisphere. N. Engl. J. Med. 2004, 350, 342–350. [Google Scholar] [CrossRef] [PubMed]
- Nalca, A.L.V.; Garza, N.; Zumbrun, E.; Frick, O.; Chapman, J.; Hartings, J. Experimental Infection of Cynomolgus Macaques (Macaca fascicularis) with Aerosolized Monkeypox Virus. PLoS ONE 2010, 5, e12880. [Google Scholar] [CrossRef] [PubMed]
- Heberling, R.K.S. Induction, Course, and Transmissibilty of Monkeypox in the Baboon (Papio cynocephalus). J. Infect. Dis. 1971, 124, 33–38. [Google Scholar] [CrossRef]
- Rimoin, A.M.P.; Johnston, S.; Muyembe, J. Major increase in human monkeypox incidence 30 years after smallpox vaccination campaigns cease in the Democratic Republic of Congo. Proc. Natl. Acad. Sci. USA 2010, 107, 16262–16267. [Google Scholar] [CrossRef]
- Beer, E.R.B. A systematic review of the epidemiology of human monkeypox outbreaks and implications for outbreak strategy. PLoS Negl. Trop. Dis. 2019, 13, e0007791. [Google Scholar] [CrossRef]
- Fine, P.Z.J.; Grab, B.; Dixon, H. The Transmission Potential of Monkeypox Virus in Human Populations. Int. J. Epidemiol. 1988, 17, 643–650. [Google Scholar] [CrossRef]
- Simpson, K.; Heymann, D.; Brown, C.S.; Edmunds, W.J.; Elsgaard, J.; Fine, P.; Hochrein, H.; Hoff, N.A.; Green, A.; Ihekweazu, C.; et al. Human monkeypox–After 40 years, an unintended consequence of smallpox eradication. Vaccine 2020, 38, 5077–5081. [Google Scholar] [CrossRef]
- Nguyen, P.Y.; Ajisegiri, W.S.; Costantino, V.; Chughtai, A.A.; MacIntyre, C.R. Reemergence of Human Monkeypox and Declining Population Immunity in the Context of Urbanization, Nigeria, 2017–2020. Emerg. Infect. Dis. 2021, 27, 1007–1014. [Google Scholar] [CrossRef]
- Sadeuh-Mba, S.A.; Yonga, M.G.; Els, M.; Batejat, C.; Eyangoh, S.; Caro, V.; Etoundi, A.; Carniel, E.; Njouom, R. Monkeypox virus phylogenetic similarities between a human case detected in Cameroon in 2018 and the 2017–2018 outbreak in Nigeria. Infect. Genet. Evol. 2019, 69, 8–11. [Google Scholar] [CrossRef]
- Nakoune, E.L.E.; Ndjapou, S.; Janssens, C.; Zuniga, I.; Van Herp, M.; Fongbia, J.; Koyazegbe, T.D.; Selekon, B.; Komoyo, G.F.; Garba-Ouangole, S.M.; et al. A Nosocomial Outbreak of Human Monkeypox in the Central African Republic. Open Forum Infect. Dis. 2017, 4, ofx168. [Google Scholar] [CrossRef] [PubMed]
- Jezek, Z.G.B.; Szczeniowski, M.; Paluku, K.; Mutombo, M. Clinico-epidemiological features of monkeypox patients with an animal or human source of infection. Bull. World Health Organ. 1988, 66, 459–464. [Google Scholar] [PubMed]
- Nolen, L.O.L.; Katomba, J.; Likofata, J.; Mukadi, D.; Monroe, B.; Doty, J.; Kalemba, L.; Malekani, J.; Malekani, J.; Kabamba, J.; et al. Introduction of Monkeypox into a Community and Household: Risk Factors and Zoonotic Reservoirs in the Democratic Republic of the Congo. Am. J. Trop. Med. Hyg. 2015, 93, 410–415. [Google Scholar] [CrossRef] [PubMed]
- Quiner, C.M.C.; Monroe, B.; Nakazawa, Y.; Doty, J.; Hughes, C.; McCollum, A.; Ibata, S.; Malekani, J.; Okitolonda, E.; Carroll, D.; et al. Presumptive risk factors for monkeypox in rural communities in the Democratic Republic of the Congo. PLoS ONE 2017, 12, e0168664. [Google Scholar]
- Kile, J.F.A.; Beard, B. Transmission of monkeypox among persons exposed to infected prairie dogs in Indiana in 2003. JAMA Pediatr. 2005, 159, 1022–1025. [Google Scholar]
- Vaughan, A.A.E.; Astbury, J.; Balasegaram, S.; Balasegaram, S.; Beadsworth, M.; Beck, C.R.; Chand, M.; O’Connor, C.; Dunning, J.; Ghebrehewet, S.; et al. Two cases of monkeypox imported to the United Kingdom, September 2018. Eurosurveillance 2018, 23, 1800509. [Google Scholar] [CrossRef]
- Hobson, G.A.J.; Adler, H.; Firth, R.; Gould, S.; Houlihan, C.; Johnson, C.; Porter, D.; Rampling, T.; Ratcliffe, L.; Russell, K.; et al. Family cluster of three cases of monkeypox imported from Nigeria to the United Kingdom, May 2021. Eurosurveillance 2021, 26, 2100745. [Google Scholar] [CrossRef]
- Yong, S.N.O.; Ho, Z.; Mak, T.; Marimuthu, K.; Vasoo, S.; Yeo, T.; Ng, Y.; Cui, L.; Ferdous, Z.; Ferdous, Z.; et al. Imported Monkeypox, Singapore. Emerg. Infect. Dis. 2020, 26, 1826–1830. [Google Scholar] [CrossRef]
- Health, M.D. Travel-Associated Monkeypox Virus Infection Confirmed in Maryland Resident; Maryland Department of Health Newsroom: Baltimore, MD, USA, 2021. [Google Scholar]
- Grant, R.; Nguyen, L.L.; Breban, R. Modelling human-to-human transmission of monkeypox. Bull. World Health Organ 2020, 98, 638–640. [Google Scholar] [CrossRef]
- Leendertz, S.S.D.; Theophil, D.; Anoh, E.; Mossoun, A.; Schubert, G.; Wiersma, L.; Akoua-Koffi, C.; Couacy-Hymann, E.; Muyembe-Tamfum, J.; Karhemere, S.; et al. A Cross-Sectional Serosurvey of Anti-Orthopoxvirus Antibodies in Central and Western Africa. Viruses 2017, 9, 278. [Google Scholar] [CrossRef]
- Gani, S.L.R. Transmission potential of smallpox in contemporary populations. Nature 2001, 414, 748–751. [Google Scholar] [CrossRef] [PubMed]
- Eichner, M.D.K. Transmission Potential of Smallpox: Estimates Based on Detailed Data from an Outbreak. Am. J. Epidemiol. 2003, 158, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Lloyd-Smith, J.S.S.; Kopp, P.; Getz, W. Superspreading and the effect of individual variation on disease emergence. Nature 2005, 438, 355–359. [Google Scholar] [CrossRef]
- Mwamba, P.T.K.; Moudi, A.; Mukinda, V.; Mwema, G.; Messinger, D. Human Monkeypox–Kasai Oriental, Zaire, 1996–1997. MMWR Morb. Mortal. Wkly. Rep. 1997, 46, 304–307. [Google Scholar]
- Blumberg, S.; Lloyd-Smith, J.O. Inference of R(0) and transmission heterogeneity from the size distribution of stuttering chains. PLoS Comput. Biol. 2013, 9, e1002993. [Google Scholar] [CrossRef] [PubMed]
- Antinori, A.M.V.; Mazzotta, V.; Vita, S.; Carletti, F.; Tacconi, D.; Lapini, L.; D’Abramo, A.; Cicalini, S.; Lapa, D.; Pittalis, S.; et al. Epidemiological, clinical, and virological characteristics of four cases of monkeypox support transmission through sexual contact, Italy, May 2022. Eurosurveillance 2022, 27, 2200421. [Google Scholar] [CrossRef]
- Adler, H.G.S.; Hine, P.; Snell, L.; Wong, W.; Houlihan, C.; Osborne, J.; Rampling, T.; Rampling, T.; Beadsworth, M.B.; Duncan, C.J.; et al. Clinical features and management of human monkeypox: A retrospective observational study in the UK. Lancet Infect. Dis. 2022, 22, 1153–1162. [Google Scholar] [CrossRef]
- Peiró-Mestres, A.; Fuertes, I.; Camprubí-Ferrer, D.; Marcos, M.Á.; Vilella, A.; Navarro, M.; Rodriguez-Elena, L.; Riera, J.; Català, A.; Martínez, M.J.; et al. Frequent detection of monkeypox virus DNA in saliva, semen, and other clinical samples from 12 patients, Barcelona, Spain, May to June 2022. Eurosurveillance 2022, 27, 2200503. [Google Scholar] [CrossRef]
- Breman, J.H.D. Diagnosis and Management of Smallpox. N. Engl. J. Med. 2002, 346, 1300–1308. [Google Scholar] [CrossRef]
- CDC. How does smallpox spread? Centers for Disease Control and Prevention. 7 June 2016. Available online: https://www.cdc.gov/smallpox/transmission/index.html (accessed on 8 September 2022).
- Mbala, P.H.J.; Riu-Rovira, T.; Ahuka, S.; Mulembakani, P.; Rimoin, A.; Martin, J.; Muyembe, J. Maternal and Fetal Outcomes Among Pregnant Women With Human Monkeypox Infection in the Democratic Republic of Congo. J. Infect. Dis. 2017, 216, 824–828. [Google Scholar] [CrossRef]
- Millard, C. Aerial Convection from Smallpox Hospitals. Br. Med. J. 1944, 1, 628–629. [Google Scholar] [CrossRef] [PubMed]
- Wehrle, P.P.J.; Richter, K.; Henderson, D. An airborne outbreak of smallpox in a German hospital and its significance with respect to other recent outbreaks in Europe. Bull. World Health Organ. 1970, 43, 669–679. [Google Scholar] [PubMed]
- Hutson, C.C.D.; Gallardo-Romero, N.; Weiss, S.; Clemmons, C.; Hughes, C.; Salzer, J.; Olson, V.; Abel, J.; Karem, K.; Damon, I. Monkeypox Disease Transmission in an Experimental Setting: Prairie Dog Animal Model. PLoS ONE 2011, 6, e28295. [Google Scholar]
- Hutson, C.C.D.; Self, J.; Weiss, S.; Hughes, C.; Braden, Z.; Olson, V.; Smith, S.; Karem, K.; Regnery, R.; Damon, I. Dosage comparison of Congo Basin and West African strains of monkeypox virus using a prairie dog animal model of systemic orthopoxvirus disease. Virology 2010, 402, 72–82. [Google Scholar] [CrossRef]
- Hutson, C.G.-R.N.; Carroll, D.; Clemmons, C.; Salzer, J.; Nagy, T.; Hughes, C.; Olson, V.; Karem, K.; Damon, I. Transmissibility of the Monkeypox Virus Clades via Respiratory Transmission: Investigation Using the Prairie Dog-Monkeypox Virus Challenge System. PLoS ONE 2013, 8, e55488. [Google Scholar]
- Reynolds, M.Y.K.; Kuehnert, M.; Davidson, W.; Huhn, G.; Holman, R.; Damon, I. Clinical Manifestations of Human Monkeypox Influenced by Route of Infection. J. Infect. Dis. 2006, 194, 773–780. [Google Scholar] [CrossRef]
- Mack, T. Smallpox in Europe, 1950–1971. J. Infect. Dis. 1972, 125, 161–169. [Google Scholar] [CrossRef]
- Downie, L.S.V.A.; Meiklejohn, G.; Ratnakannan, N.; Rao, A.; Krishnan, G.; Kempe, C. Studies on the virus content of mouth washings in the acute phase of smallpox. Bull. World Health Organ. 1961, 25, 49–53. [Google Scholar]
- Nolen, L.O.L.; Katomba, J.; Likofata, J.; Mukadi, D.; Monroe, B.; Doty, J.; Hughes, C.; Kabamba, J.; Malekani, J.; Bomponda, P.L.; et al. Extended Human-to-Human Transmission during a Monkeypox Outbreak in the Democratic Republic of the Congo. Emerg. Infect. Dis. 2016, 22, 1014–1021. [Google Scholar] [CrossRef]
- Vaughan, A.A.E.; Astbury, J.; Brooks, T.; Chand, M.; Flegg, P.; Hardman, A.; Hardman, A.; Harper, N.; Jarvis, R.; Mawdsley, S.; et al. Human-to-Human Transmission of Monkeypox Virus, United Kingdom, October 2018. Emerg. Infect. Dis. 2020, 26, 782–785. [Google Scholar] [CrossRef]
- Ogoina, D.I.J.; Ogunleye, A.; Ederiane, E.; Anebonam, U.; Anebonam, U.; Neni, A.; Oyeyemi, A.; Etebu, E.N.; Ihekweazu, C. The 2017 human monkeypox outbreak in Nigeria-report of outbreak experience and response in the Niger Delta University Teaching Hospital, Bayelsa State, Nigeria. PLoS ONE 2019, 14, e0214229. [Google Scholar] [CrossRef] [PubMed]
- CDC. Smallpox–Stockholm, Sweden, 1963. In Morbidity and Mortality Weekly Report; Centers for Disease Control and Prevention: Atlanta, GA, USA, 1963. [Google Scholar]
- Kiang, K.K.M. Rates and risks of transmission of smallpox and mechanisms of prevention. J. Lab. Clin. Med. 2003, 142, 229–238. [Google Scholar] [CrossRef]
- Judson, S.M.V. Nosocomial Transmission of Emerging Viruses via Aerosol-Generating Medical Procedures. Viruses 2019, 11, 940. [Google Scholar] [CrossRef] [PubMed]
- Learned, L.A.; Reynolds, M.G.; Wassa, D.W.; Li, Y.; Olson, V.A.; Karem, K.; Stempora, L.L.; Braden, Z.H.; Kline, R.; Likos, A.; et al. Extended interhuman transmission of monkeypox in a hospital community in the Republic of the Congo, 2003. Am. J. Trop. Med. Hyg. 2005, 73, 428–434. [Google Scholar] [CrossRef] [PubMed]
- Monkeypox and Smallpox Vaccine Guidance. CDC. 2 June 2022. Available online: https://www.cdc.gov/poxvirus/monkeypox/clinicians/smallpox-vaccine.html (accessed on 8 September 2022).
- Considerations for Monkeypox Vaccination. CDC 22 August 2022. Available online: https://www.cdc.gov/poxvirus/monkeypox/health-departments/vaccine-considerations.html (accessed on 8 September 2022).
- Rao, A.K.; Petersen, B.W.; Whitehill, F.; Razeq, J.H.; Isaacs, S.N.; Merchlinsky, M.J.; Campos-Outcalt, D.; Morgan, R.L.; Damon, I.; Sánchez, P.J.; et al. Use of JYNNEOS (Smallpox and Monkeypox Vaccine, Live, Nonreplicating) for Preexposure Vaccination of Persons at Risk for Occupational Exposure to Orthopoxviruses: Recommendations of the Advisory Committee on Immunization Practices-United States, 2022. MMWR Morb. Mortal. Wkly. Rep. 2022, 71, 734–742. [Google Scholar] [CrossRef]
- Damon, I. Status of human monkeypox: Clinical disease, epidemiology, and research. Vaccine 2011, 29, D54–D59. [Google Scholar] [CrossRef]
- Epidemiological Update: Monkeypox Multi-Country Outbreak. European Centre for Disease Prevention and Control. 2022. Available online: https://www.ecdc.europa.eu/en/news-events/epidemiological-update-monkeypox-multi-country-outbreak-15-june (accessed on 8 September 2022).
- Guzzetta, G.; Mammone, A.; Ferraro, F.; Caraglia, A.; Rapiti, A.; Marziano, V.; Poletti, P.; Cereda, D.; Vairo, F.; Mattei, G.; et al. Early Estimates of Monkeypox Incubation Period, Generation Time, and Reproduction Number, Italy, May–June 2022. Emerg. Infect. Dis. 2022, 28. [Google Scholar] [CrossRef]
- Du, Z.; Shao, Z.; Bai, Y.; Wang, L.; Herrera-Diestra, J.L.; Fox, S.J.; Ertem, Z.; Lau, E.H.Y.; Cowling, B.J. Reproduction number of monkeypox in the early stage of the 2022 multi-country outbreak. J. Travel Med. 2022, taac099, Online ahead of print. [Google Scholar] [CrossRef]
- Isidro, J.B.V.; Pinto, M.; Sobral, D.; Santos, J.; Nunes, A.; Mixao, V.; Ferreira, R.; Santos, D.; Duarte, S. Phylogenomic characterization and signs of microevolution in the 2022 multi-country outbreak of monkeypox virus. Nat. Med. 2022, 28, 1569–1572. [Google Scholar] [CrossRef]
- Seang, S.; Burrel, S.; Todesco, E.; Leducq, V.; Monsel, G.; Le Pluart, D.; Cordevant, C.; Pourcher, V.; Palich, R. Evidence of human-to-dog transmission of monkeypox virus. Lancet 2022, 400, 658–659. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5, 113. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–w296. [Google Scholar] [CrossRef] [PubMed]
- Philpott, D.; Hughes, C.M.; Alroy, K.A.; Kerins, J.L.; Pavlick, J.; Asbel, L.; Crawley, A.; Newman, A.P.; Spencer, H.; Feldpausch, A.; et al. Epidemiologic and Clinical Characteristics of Monkeypox Cases-United States, 17 May–22 July, 2022. MMWR Morb. Mortal. Wkly. Rep. 2022, 71, 1018–1022. [Google Scholar] [CrossRef] [PubMed]
- Bragazzi, N.K.J.; Mahroum, N.; Tsigalou, C.; Khamisy-Farah, R.; Converti, M.; Wu, J. Epidemiological trends and clinical features of the ongoing mokeypox epidemic: A preliminary pooled data analysis and literature review. J. Med. Virol. 2022. [Google Scholar] [CrossRef] [PubMed]
- Shao, H.; McDonald, E.C.; Ginsberg, M.M.; Yee, L.M.; Montgomery, J.R.; Allan-Martinez, F.; Reynolds, M.G.; Tack, D.M.; Davidson, W.; Patel, N.; et al. Secondary and Tertiary Transmission of Vaccinia Virus after Sexual Contact with a Smallpox Vaccinee—San Diego, California, 2012. MMWR Morb. Mortal. Wkly. Rep. 2013, 62, 145–147. [Google Scholar]
- Lapa, D.; Carletti, F.; Mazzotta, V.; Matusali, G.; Pinnetti, C.; Meschi, S.; Gagliardini, R.; Colavita, F.; Mondi, A.; Minosse, C.; et al. Monkeypox virus isolation from a semen sample collected in the early phase of infection in a patient with prolonged seminal viral shedding. Lancet Infect. Dis. 2022, 22, 1267–1269. [Google Scholar] [CrossRef]
- Perez Duque, M.; Ribeiro, S.; Martins, J.V.; Casaca, P.; Leite, P.P.; Tavares, M.; Mansinho, K.; Duque, L.M.; Fernandes, C.; Cordeiro, R.; et al. Ongoing monkeypox virus outbreak, Portugal, 29 April to 23 May 2022. Eurosurveillance 2022, 27, 2200424. [Google Scholar] [CrossRef]
- CDC. Demographics of Patients Receiving TPOXX for Treatment of Monkeypox 2022. Available online: https://www.cdc.gov/poxvirus/monkeypox/response/2022/demographics-TPOXX.html (accessed on 31 August 2022).
- Tutu van Furth, A.M.; van der Kuip, M.; van Els, A.L.; Fievez, L.C.; van Rijckevorsel, G.G.; van den Ouden, A.; Jonges, M.; Welkers, M.R. Paediatric monkeypox patient with unknown source of infection, the Netherlands, June 2022. Eurosurveillance 2022, 27, 2200552. [Google Scholar] [CrossRef]
- Endo, A.M.H.; Abbott, S.; Ratnayake, R.; Pearson, C.; Edmunds, W.; Fearon, E.; Funk, S. Heavy-tailed sexual contact networks and the epidemiology of monkeypox outbreak in non-endemic regions, May 2022. MedRxiv 2022. [Google Scholar] [CrossRef]
- Bisanzio, D.R.R. Projected burden and duration of the 2022 Monkeypox outbreaks in non-endemic countries. Lancet Microbe 2022, 3, e643. [Google Scholar] [CrossRef]
- Heskin, J.B.A.; Milne, C.; Brown, N.; Walters, Y.; Scott, C.; Bracchi, M.; Moore, L.; Mughal, N.; Rampling, T.; Winston, A.; et al. Transmission of monkeypox virus through sexual contact–A novel route of infection. J. Infect. 2022, 85, 334–363. [Google Scholar] [CrossRef] [PubMed]
- Minhaj, F.O.Y.; Whitehill, F.; Schultz, J.; Foote, M.; Davidson, W.; Hughes, C.; Wilkins, K.; Bachmann, L.; Chatelain, R.; Donnelly, M.A.P.; et al. Monkeypox Outbreak–Nine States, May 2022. MMWR Morb. Mortal. Wkly. Rep. 2022, 71, 764–769. [Google Scholar] [CrossRef] [PubMed]
- Girometti, N.B.R.; Bracchi, M.; Heskin, J.; McOwan, A.; Tittle, V.; Gedela, K.; Scott, C.; Patel, S.; Gohil, J.; Nugent, D.; et al. Demographic and Clinical Characteristics of Confirmed Human Monkeypox Virus Cases in Individuals Attending a Sexual Health Centre in London, United Kingdom: An Observational Analysis. Lancet Inf. Dis. 2022, 22, 1321–1328. [Google Scholar] [CrossRef]
- Charniga, K.M.N.; Slayton, R.; Gosdin, L.; Minhaj, F.; Philpott, D.; Smith, D.; Gearhart, S.; Alvarado-Ramy, F.; Brown, C.; Waltenburg, M.A.; et al. Estimating the incubation period of monkeypox virus during the 2022 multi-national outbreak. medRxiv 2022. [Google Scholar] [CrossRef]
- Miura, F.E.C.; Backer, J.; Xiridou, M.; Franz, E.; Coul, E.; Brandwagt, D.; van Cleef, B.; van Rijckevorsel, G.; Swaan, C.; van den Hof, S.; et al. Estimated incubation period for monkeypox cases confirmed in the Netherlands, May 2022. Eurosurveillance 2022, 27, 200448. [Google Scholar] [CrossRef]
- Ogoina, D.; Yinka-Ogunleye, A. Sexual history of human monkeypox patients seen at a tertiary hospital in Bayelsa, Nigeria. Int. J. STD AIDS 2022, 1–5. [Google Scholar] [CrossRef]
- Ogoina, D.I.M.; James, H.; Oladokun, R.; Yinka-Ogunleye, A.; Wakama, P.; Otike-Odibi, B.; Usman, L.M.; Obazee, E.; Aruna, O.; Ihekweazu, C.; et al. Clinical Course and Outcome of Human Monkeypox in Nigeria. Clin. Infect. Dis. 2020, 71, 210–214. [Google Scholar] [CrossRef]
- Ferré, V.M.; Bachelard, A.; Zaidi, M.; Armand-Lefevre, L.; Descamps, D.; Charpentier, C.; Ghosn, J. Detection of Monkeypox Virus in Anorectal Swabs From Asymptomatic Men Who Have Sex With Men in a Sexually Transmitted Infection Screening Program in Paris, France. Ann. Intern. Med. 2022; Online ahead of print. [Google Scholar]
- National Technical Guidelines for Integrated Disease Surveillance and Response. Monrovia. 2021. Available online: https://www.afro.who.int/sites/default/files/2021-09/Final_IDSR%20Technical%20Guidelines_Final%20Version%20signed%20by%20%20Hon.%20DG%20and%20Hon.%20Minister.pdf (accessed on 8 September 2022).
- Wang, H.; de Paulo, K.J.D.A.; Gültzow, T.; Zimmermann, H.M.L.; Jonas, K.J. Monkeypox self-diagnosis abilities, determinants of vaccination and self-isolation intention after diagnosis among MSM, the Netherlands, July 2022. Eurosurveillance 2022, 27, 2200603. [Google Scholar] [CrossRef]
- Rueda, S.M.S.; Chen, S.; Gogolishvili, D.; Globerman, J.; Chambers, L.; Wilson, M.; Logie, C.; Shi, Q.; Morassaei, S.; Rourke, B. Examining the associations between HIV-related stigma and health outcomes in people living with HIV/AIDS: A series of meta-analyses. BMJ Open 2016, 6, e011453. [Google Scholar] [CrossRef]
- Mendos, L.R. State-Sponsored Homophobia 2019; International Lesbian, Gay, Bisexual, Trans and Intersex Association: Geneva, Switzerland, 2019. [Google Scholar]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Riopelle, J.C.; Munster, V.J.; Port, J.R. Atypical and Unique Transmission of Monkeypox Virus during the 2022 Outbreak: An Overview of the Current State of Knowledge. Viruses 2022, 14, 2012. https://doi.org/10.3390/v14092012
Riopelle JC, Munster VJ, Port JR. Atypical and Unique Transmission of Monkeypox Virus during the 2022 Outbreak: An Overview of the Current State of Knowledge. Viruses. 2022; 14(9):2012. https://doi.org/10.3390/v14092012
Chicago/Turabian StyleRiopelle, Jade C., Vincent J. Munster, and Julia R. Port. 2022. "Atypical and Unique Transmission of Monkeypox Virus during the 2022 Outbreak: An Overview of the Current State of Knowledge" Viruses 14, no. 9: 2012. https://doi.org/10.3390/v14092012
APA StyleRiopelle, J. C., Munster, V. J., & Port, J. R. (2022). Atypical and Unique Transmission of Monkeypox Virus during the 2022 Outbreak: An Overview of the Current State of Knowledge. Viruses, 14(9), 2012. https://doi.org/10.3390/v14092012