
Spatially Segregated Transmission of Co-Occluded Baculoviruses Limits Virus–Virus Interactions Mediated by Cellular Coinfection during Primary Infection



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Insects
2.2. Construction of Recombinant Viruses
2.3. Viral Titration
2.4. Flow Cytometry
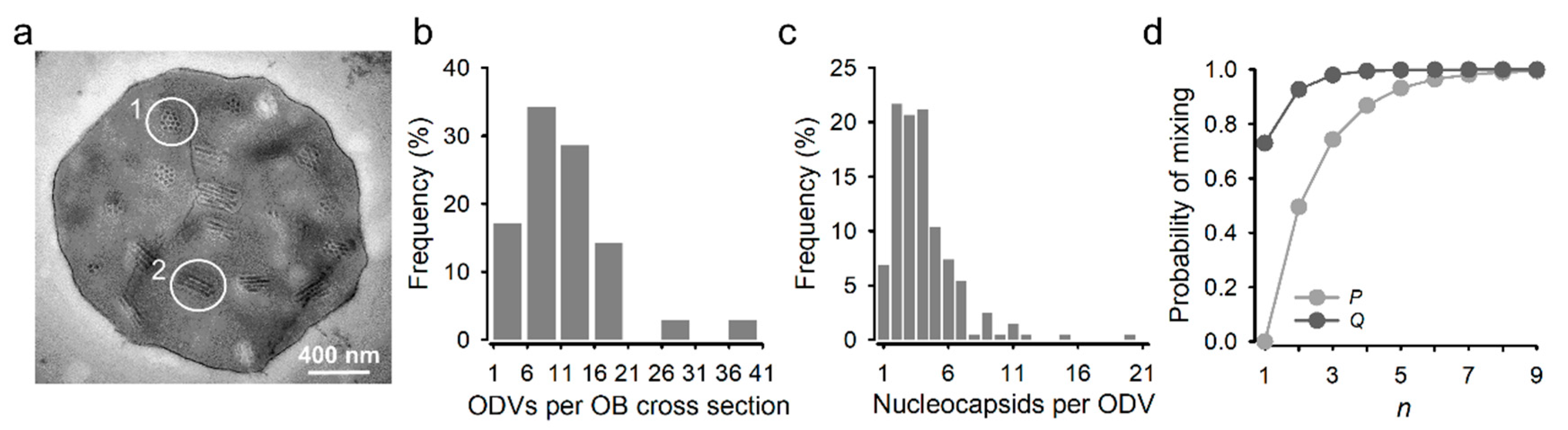
2.5. OB Preparation
2.6. Transmission Electron Microscopy
2.7. Midgut Infection Analysis
2.8. Hemolymph-Derived BVs Extraction
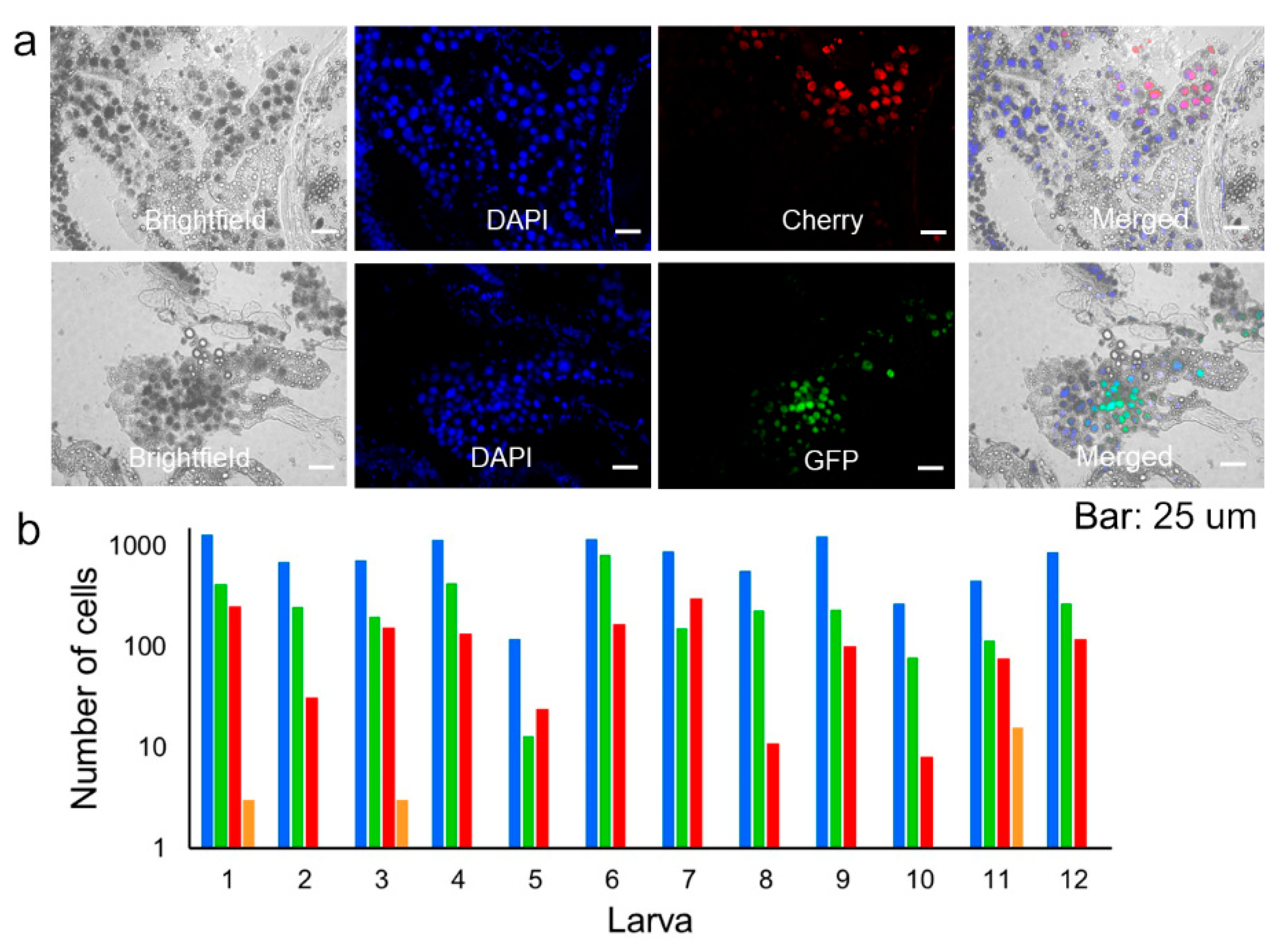
2.9. Determination of Coinfected Cells in Whole Larvae
2.10. PCR Detection of Spontaneous Recombinants
2.11. Foci Assay-Based Detection of Spontaneous Recombinants
2.12. Calculation of Expected Coinfection Rates under Free Assortment
3. Results
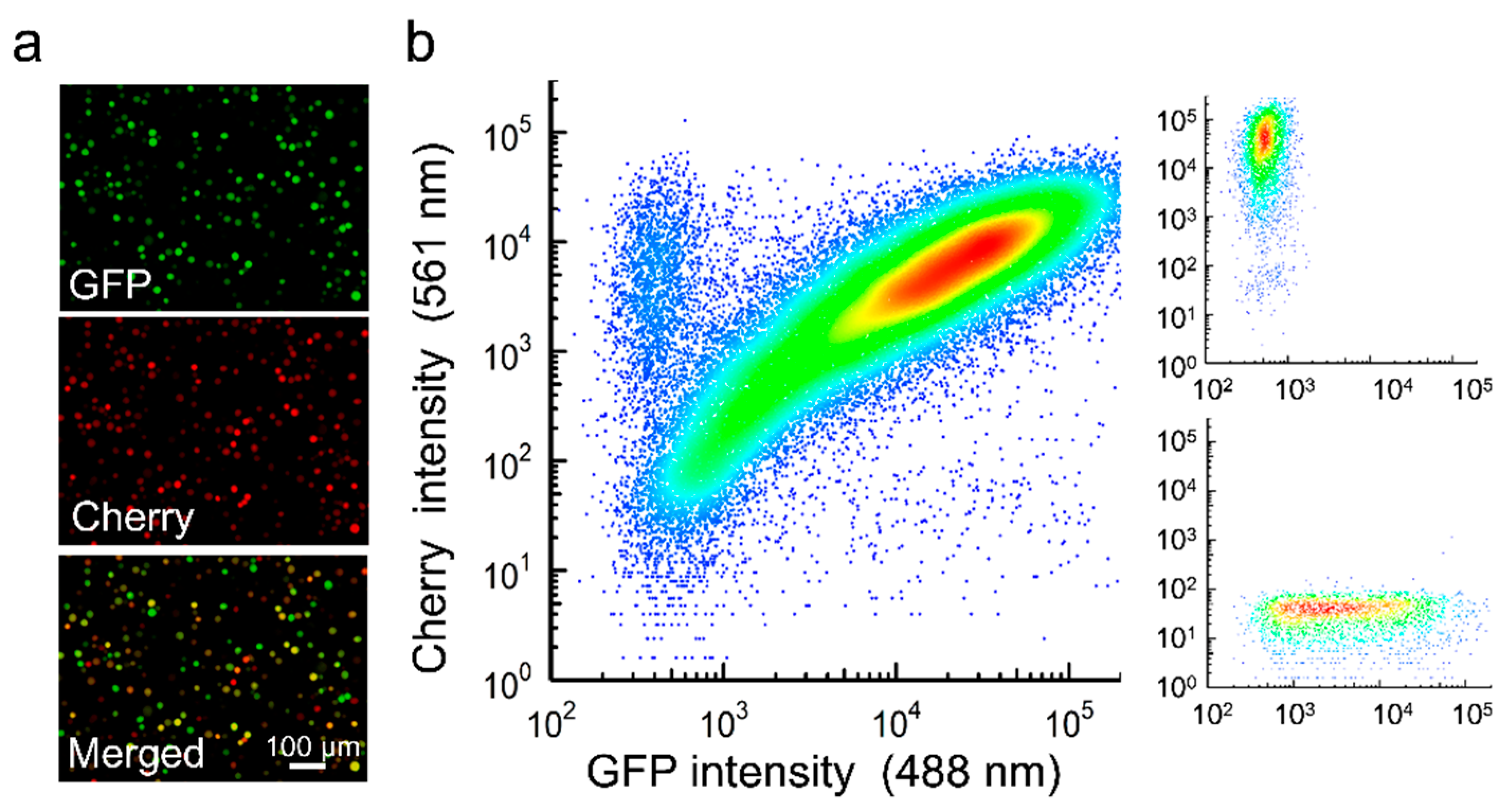
3.1. Production of OBs from Cells Coinfected with GFP- and Cherry-Encoding Viruses
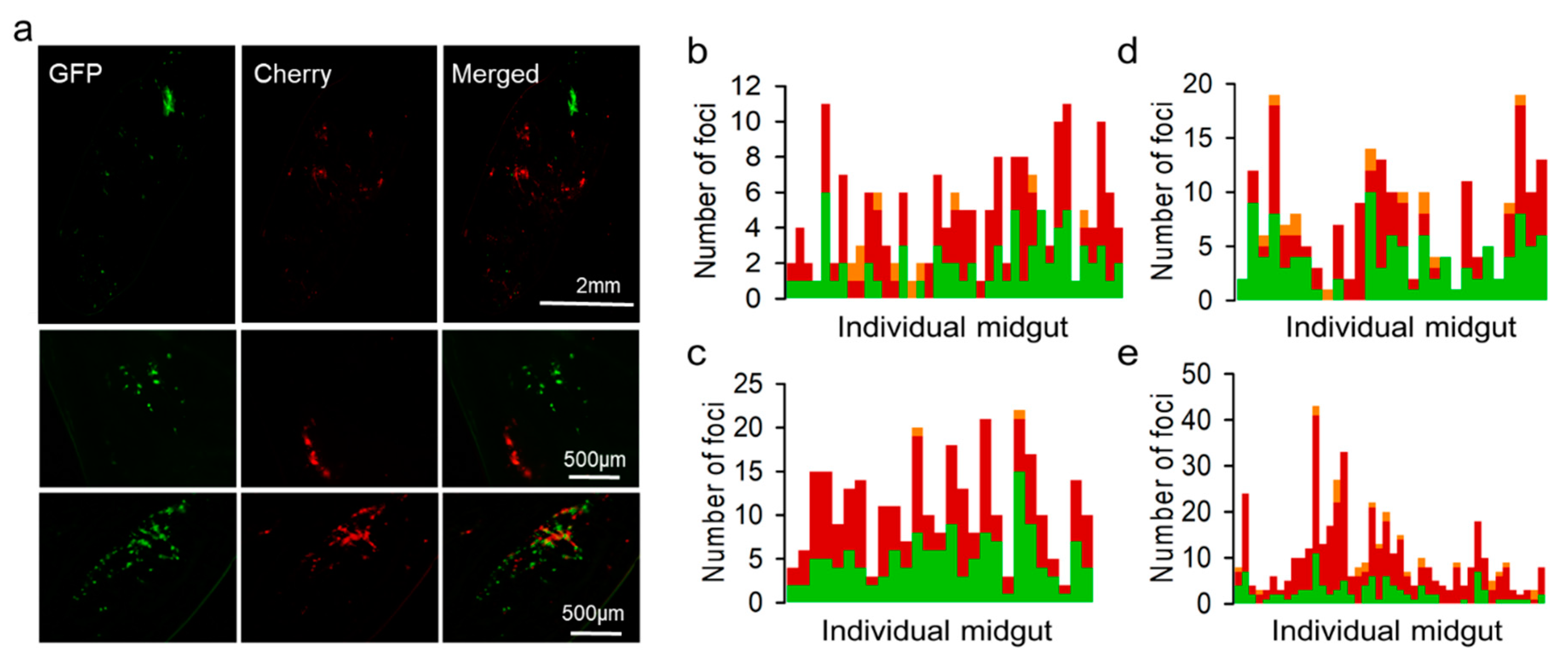
3.2. Spatial Segregation of GFP and Cherry Variants In Vivo
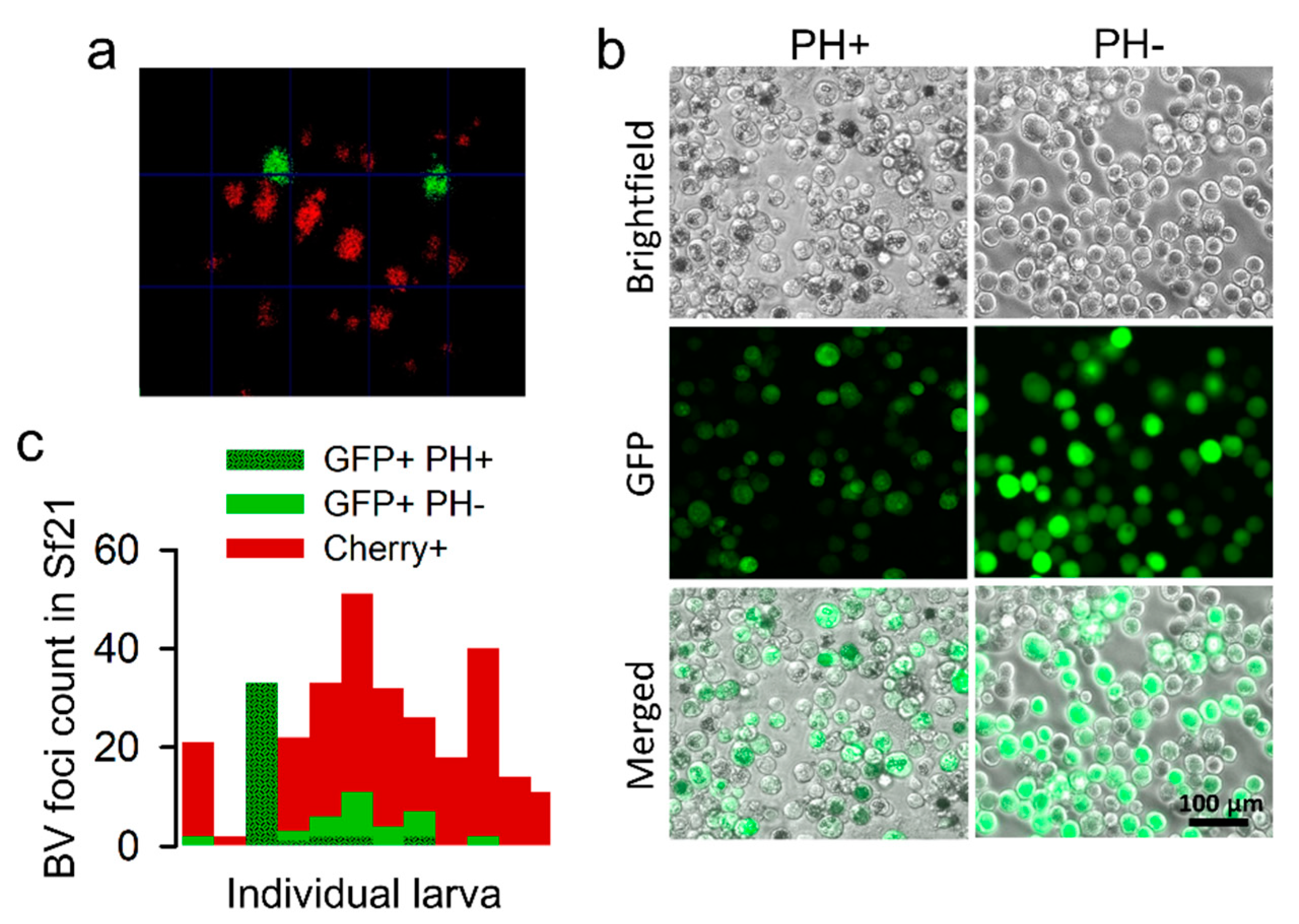
3.3. Ability of a Polyhedrin-Defective Virus to Exploit OBs from a Functional Virus
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sanjuán, R. Collective Infectious Units in Viruses. Trends Microbiol. 2017, 25, 402–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanjuán, R.; Thoulouze, M.-I. Why Viruses Sometimes Disperse in Groups. Virus Evol. 2019, 5, vez014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urbanelli, L.; Buratta, S.; Tancini, B.; Sagini, K.; Delo, F.; Porcellati, S.; Emiliani, C. The Role of Extracellular Vesicles in Viral Infection and Transmission. Vaccines 2019, 7, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguilera, E.R.; Pfeiffer, J.K. Strength in Numbers: Mechanisms of Viral Co-Infection. Virus Res. 2019, 265, 43–46. [Google Scholar] [CrossRef]
- Altan-Bonnet, N.; Perales, C.; Domingo, E. Extracellular Vesicles: Vehicles of En Bloc Viral Transmission. Virus Res. 2019, 265, 143–149. [Google Scholar] [CrossRef]
- Ikeda, M.; Hamajima, R.; Kobayashi, M. Baculoviruses: Diversity, Evolution and Manipulation of Insects. Entomol. Sci. 2015, 18, 1–20. [Google Scholar] [CrossRef]
- Slack, J.; Arif, B.M. The Baculoviruses Occlusion-Derived Virus: Virion Structure and Function. Adv. Virus Res. 2006, 69, 99–165. [Google Scholar] [CrossRef]
- Rohrmann, G.F. Baculovirus Molecular Biology; National Center for Biotechnology Information: Bethesda, MD, USA, 2019; pp. 1–298. [Google Scholar]
- Rohrmann, G.F. Baculovirus Nucleocapsid Aggregation (MNPV vs SNPV): An Evolutionary Strategy, or a Product of Replication Conditions? Virus Genes 2014, 49, 351–357. [Google Scholar] [CrossRef]
- Clem, R.J.; Passarelli, A.L. Baculoviruses: Sophisticated Pathogens of Insects. PLoS Pathog. 2013, 9, e1003729. [Google Scholar] [CrossRef]
- Williams, T.; López-Ferber, M.; Caballero, P. Nucleopolyhedrovirus Coocclusion Technology: A New Concept in the Development of Biological Insecticides. Front. Microbiol. 2022, 12. [Google Scholar] [CrossRef]
- Granados, R.R.; Lawler, K.A. In Vivo Pathway of Autographa Californica Baculovirus Invasion and Infection. Virology 1981, 108, 297–308. [Google Scholar] [CrossRef]
- Washburn, J.O.; Lyons, E.H.; Haas-Stapleton, E.J.; Volkman, L.E. Multiple Nucleocapsid Packaging of Autographa Californica Nucleopolyhedrovirus Accelerates the Onset of Systemic Infection in Trichoplusia Ni. J. Virol. 1999, 73, 411–416. [Google Scholar] [CrossRef] [Green Version]
- Simón, O.; Williams, T.; López-Ferber, M.; Caballero, P. Genetic Structure of a Spodoptera Frugiperda Nucleopolyhedrovirus Population: High Prevalence of Deletion Genotypes. Appl. Environ. Microbiol. 2004, 70, 5579–5588. [Google Scholar] [CrossRef] [Green Version]
- Chateigner, A.; Bézier, A.; Labrousse, C.; Jiolle, D.; Barbe, V.; Herniou, E.A. Ultra Deep Sequencing of a Baculovirus Population Reveals Widespread Genomic Variations. Viruses 2015, 7, 3625–3646. [Google Scholar] [CrossRef] [Green Version]
- Loiseau, V.; Herniou, E.A.; Moreau, Y.; Lévêque, N.; Meignin, C.; Daeffler, L.; Federici, B.; Cordaux, R.; Gilbert, C. Wide Spectrum and High Frequency of Genomic Structural Variation, Including Transposable Elements, in Large Double-Stranded DNA Viruses. Virus Evol. 2020, 6, 1–16. [Google Scholar] [CrossRef]
- Simón, O.; Williams, T.; Caballero, P.; López-Ferber, M. Dynamics of Deletion Genotypes in an Experimental Insect Virus Population. Proc. R. Soc. B Biol. Sci. 2006, 273, 783–790. [Google Scholar] [CrossRef] [Green Version]
- Clavijo, G.; Williams, T.; Muñoz, D.; Caballero, P.; Lopez-Ferber, M. Mixed Genotype Transmission Bodies and Virions Contribute to the Maintenance of Diversity in an Insect Virus. Proc. R. Soc. B Biol. Sci. 2010, 277, 943–951. [Google Scholar] [CrossRef] [Green Version]
- Bull, J.C.; Godfray, H.C.J.; O’Reilly, D.R. Persistence of an Occlusion-Negative Recombinant Nucleopolyhedrovirus in Trichoplusia Ni Indicates High Multiplicity of Cellular Infection. Appl. Environ. Microbiol. 2001, 67, 5204–5209. [Google Scholar] [CrossRef] [Green Version]
- Genoyer, E.; López, C.B. The Impact of Defective Viruses on Infection and Immunity. Annu. Rev. Virol. 2019, 6, 547–566. [Google Scholar] [CrossRef]
- Muñoz, D.; Castillejo, J.I.; Caballero, P. Naturally Occurring Deletion Mutants Are Parasitic Genotypes in a Wild- Type Nucleopolyhedrovirus Population of Spodoptera Exigua. Appl. Environ. Microbiol. 1998, 64, 4372–4377. [Google Scholar] [CrossRef] [Green Version]
- López-Ferber, M.; Simón, O.; Williams, T.; Caballero, P. Defective or Effective? Mutualistic Interactions between Virus Genotypes. Proc. R. Soc. B Biol. Sci. 2003, 270, 2249–2255. [Google Scholar] [CrossRef] [Green Version]
- Serrano, A.; Williams, T.; Simón, O.; López-Ferber, M.; Caballero, P.; Muñoz, D. Analagous Population Structures for Two Alphabaculoviruses Highlight a Functional Role for Deletion Mutants. Appl. Environ. Microbiol. 2013, 79, 1118–1125. [Google Scholar] [CrossRef]
- Clavijo, G.; Williams, T.; Simón, O.; Muñoz, D.; Cerutti, M.; López-Ferber, M.; Caballero, P. Mixtures of Complete and Pif1- and Pif2-Deficient Genotypes Are Required for Increased Potency of an Insect Nucleopolyhedrovirus. J. Virol. 2009, 83, 5127–5136. [Google Scholar] [CrossRef] [Green Version]
- Kikhno, I.; Gutiérrez, S.; Croizier, L.; Crozier, G.; López Ferber, M. Characterization of Pif, a Gene Required for the per Os Infectivity of Spodoptera Littoralis Nucleopolyhedrovirus. J. Gen. Virol. 2002, 83, 3013–3022. [Google Scholar] [CrossRef] [PubMed]
- Elvira, S.; Gorría, N.; Muñoz, D.; Williams, T.; Caballero, P. A Simplified Low-Cost Diet for Rearing Spodoptera Exigua (Lepidoptera: Noctuidae) and Its Effect on S. Exigua Nucleopolyhedrovirus Production. J. Econ. Entomol. 2010, 103, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Targovnik, A.M.; Ferrari, A.; Mc Callum, G.J.; Arregui, M.B.; Smith, I.; Bracco, L.F.; Alfonso, V.; López, M.G.; Martínez-Solís, M.; Herrero, S.; et al. Highly Efficient Production of Rabies Virus Glycoprotein G Ectodomain in Sf9 Insect Cells. 3 Biotech 2019, 9, 385. [Google Scholar] [CrossRef] [PubMed]
- Invitrogen life technologies. Instruction Manual. Guide to Baculovirus Expression Vector Systems (BEVS) and Insect Cell Culture Techniques. Available online: https://tools.thermofisher.com/content/sfs/manuals/bevtest.pdf (accessed on 19 May 2022).
- Caballero, P.; Lopez-Ferber, M.; Williams, T. Los Baculovirus y Sus Aplicaciones Como Bioinsecticidad En El Control Biológico de Plagas; Phytoma S.A: Valencia, Spain, 2001; pp. 490–493. ISBN 84-932056-0-5. [Google Scholar]
- Zwart, M.P.; van Oers, M.M.; Cory, J.S.; van Lent, J.W.M.; van der Werf, W.; Vlak, J.M. Development of a Quantitative Real-Time PCR for Determination of Genotype Frequencies for Studies in Baculovirus Population Biology. J. Virol. Methods 2008, 148, 146–154. [Google Scholar] [CrossRef]
- Smith, D.R.; Adams, A.P.; Kenney, J.L.; Wang, E.; Weaver, S.C. Venezuelan Equine Encephalitis Virus in the Mosquito Vector Aedes Taeniorhynchus: Infection Initiated by a Small Number of Susceptible Epithelial Cells and a Population Bottleneck. Virology 2008, 372, 176–186. [Google Scholar] [CrossRef] [Green Version]
- Summers, M.D.; Volkman, L.E. Comparison of Biophysical and Morphological Properties of Occluded and Extracellular Nonoccluded Baculovirus from in Vivo and in Vitro Host Systems. J. Virol. 1976, 17, 962–972. [Google Scholar] [CrossRef] [Green Version]
- Engelhard, E.K.; Volkman, L.E. Developmental Resistance in Fourth Instar Trichoplusia Ni Orally Inoculated with Autographa Californica M Nuclear Polyhedrosis Virus. Virology 1995, 209, 384–389. [Google Scholar] [CrossRef] [Green Version]
- Inoue, H.; Miyagawa, M. Regeneration of Midgut Epithelial Cell in the Silkworm, Bombyx Mori, Infected with Viruses. J. Invertebr. Pathol. 1978, 32, 373–380. [Google Scholar] [CrossRef]
- Shrestha, A.; Bao, K.; Chen, Y.-R.; Chen, W.; Wang, P.; Fei, Z.; Blissard, G.W. Global Analysis of Baculovirus Autographa Californica Multiple Nucleopolyhedrovirus Gene Expression in the Midgut of the Lepidopteran Host Trichoplusia Ni. J. Virol. 2018, 92. [Google Scholar] [CrossRef] [Green Version]
- Flipsen, J.T.; Martens, J.W.M.; Van Oers, M.M.; Vlak, J.M.; Van Lent, J.W.M. Passage of Autographa Californica Nuclear Polyhedrosis Virus through the Midgut Epithelium of Spodoptera Exigua Larvae. Virology 1995, 208, 328–335. [Google Scholar] [CrossRef] [Green Version]
- Zwart, M.P.; Erro, E.; van Oers, M.M.; de Visser, J.A.G.M.; Vlak, J.M. Low Multiplicity of Infection in Vivo Results in Purifying Selection against Baculovirus Deletion Mutants. J. Gen. Virol. 2008, 89, 1220–1224. [Google Scholar] [CrossRef]
- Bull, J.C.; Godfray, H.C.J.; O’Reilly, D.R. A Few-Polyhedra Mutant and Wild-Type Nucleopolyhedrovirus Remain as a Stable Polymorphism during Serial Coinfection in Trichoplusia Ni. Appl. Environ. Microbiol. 2003, 69, 2052–2057. [Google Scholar] [CrossRef] [Green Version]
- Hamblin, M.; Van Beek, N.A.M.; Hughes, P.R.; Wood, H.A. Co-Occlusion and Persistance of a Baculovirus Mutant Lacking the Polyhedrin Gene. Appl. Environ. Microbiol. 1990, 56, 3057–3062. [Google Scholar] [CrossRef] [Green Version]
- Beperet, I.; Simón, O.; López-Ferber, M.; van Lent, J.; Williams, T.; Caballero, P. Mixtures of Insect-Pathogenic Viruses in a Single Virion: Towards the Development of Custom-Designed Insecticides. Appl. Environ. Microbiol. 2021, 87, 1–15. [Google Scholar] [CrossRef]
- Arrizubieta, M.; Simon, O.; Ricarte-Bermejo, A.; López-Ferber, M.; Williams, T.; Caballero, P. Coocclusion of Helicoverpa Armigera Single Nucleopolyhedrovirus (HearSNPV) and Helicoverpa Armigera Multiple Nucleopolyhedrovirus (HearMNPV): Pathogenicity and Stability in Homologous and Heterologous Hosts. Viruses 2022, 14, 687. [Google Scholar] [CrossRef]
- Young, J.C.; MacKinnon, E.A.; Faulkner, P. The Architecture of the Virogenic Stroma in Isolated Nuclei of Spodoptera Frugiperda Cells in Vitro Infected by Autographa Californica Nuclear Polyhedrosis Virus. J. Struct. Biol. 1993, 110, 141–153. [Google Scholar] [CrossRef]
- Godfray, H.C.J.; O’Reilly, D.R.; Briggs, C.J. A Model of Nucleopolyhedrovirus (NPV) Population Genetics Applied to Co-Occlusion and the Spread of the Few Polyhedra (FP) Phenotype. Proc. R. Soc. B Biol. Sci. 1997, 264, 315–322. [Google Scholar] [CrossRef] [Green Version]
- Beperet, I.; Irons, S.L.; Simon, O.; King, L.A.; Williams, T.; Possee, R.D.; Lopez-Ferber, M.; Caballero, P. Superinfection Exclusion in Alphabaculovirus Infections Is Concomitant with Actin Reorganization. J. Virol. 2014, 88, 3548–3556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pijlman, G.P.; Van Den Born, E.; Martens, D.E.; Vlak, J.M. Autographa Californica Baculoviruses with Large Genomic Deletions Are Rapidly Generated in Infected Insect Cells. Virology 2001, 283, 132–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardner, A.; West, S.A.; Wild, G. The Genetical Theory of Kin Selection. J. Evol. Biol. 2011, 24, 1020–1043. [Google Scholar] [CrossRef] [PubMed]
- West, S.A.; Griffin, A.S.; Gardner, A.; Diggle, S.P. Social Evolution Theory for Microorganisms. Nat. Rev. Microbiol. 2006, 4, 597–607. [Google Scholar] [CrossRef]
- Domingo-Calap, P.; Mora-Quilis, L.; Sanjuán, R. Social Bacteriophages. Microorganisms 2020, 8, 533. [Google Scholar] [CrossRef] [Green Version]
- Bou, J.V.; Geller, R.; Sanjuán, R. Membrane-Associated Enteroviruses Undergo Intercellular Transmission as Pools of Sibling Viral Genomes. Cell Rep. 2019, 29, 714–723. [Google Scholar] [CrossRef] [Green Version]
- Law, K.M.; Komarova, N.L.; Yewdall, A.W.; Lee, R.K.; Herrera, O.L.; Wodarz, D.; Chen, B.K. In Vivo HIV-1 Cell-to-Cell Transmission Promotes Multicopy Micro-Compartmentalized Infection. Cell Rep. 2016, 15, 2771–2783. [Google Scholar] [CrossRef] [Green Version]
- Miyashita, S.; Ishibashi, K.; Kishino, H.; Ishikawa, M. Viruses Roll the Dice: The Stochastic Behavior of Viral Genome Molecules Accelerates Viral Adaptation at the Cell and Tissue Levels. PLoS Biol. 2015, 13, e1002094. [Google Scholar] [CrossRef]
- Andreu-Moreno, I.; Bou, J.V.; Sanjuán, R. Cooperative Nature of Viral Replication. Sci. Adv. 2020, 6, eabd4942. [Google Scholar] [CrossRef]
- Andreu-Moreno, I.; Sanjuán, R. Collective Infection of Cells by Viral Aggregates Promotes Early Viral Proliferation and Reveals a Cellular-Level Allee Effect. Curr. Biol. 2018, 28, 3212–3219. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pazmiño-Ibarra, V.; Herrero, S.; Sanjuan, R. Spatially Segregated Transmission of Co-Occluded Baculoviruses Limits Virus–Virus Interactions Mediated by Cellular Coinfection during Primary Infection. Viruses 2022, 14, 1697. https://doi.org/10.3390/v14081697
Pazmiño-Ibarra V, Herrero S, Sanjuan R. Spatially Segregated Transmission of Co-Occluded Baculoviruses Limits Virus–Virus Interactions Mediated by Cellular Coinfection during Primary Infection. Viruses. 2022; 14(8):1697. https://doi.org/10.3390/v14081697
Chicago/Turabian StylePazmiño-Ibarra, Verónica, Salvador Herrero, and Rafael Sanjuan. 2022. "Spatially Segregated Transmission of Co-Occluded Baculoviruses Limits Virus–Virus Interactions Mediated by Cellular Coinfection during Primary Infection" Viruses 14, no. 8: 1697. https://doi.org/10.3390/v14081697
APA StylePazmiño-Ibarra, V., Herrero, S., & Sanjuan, R. (2022). Spatially Segregated Transmission of Co-Occluded Baculoviruses Limits Virus–Virus Interactions Mediated by Cellular Coinfection during Primary Infection. Viruses, 14(8), 1697. https://doi.org/10.3390/v14081697