
The Emergence and Pathogenesis of Recombinant Viruses Associated with NADC34-like Strains and the Predominant Circulating Strains of Porcine Reproductive and Respiratory Syndrome Virus in Southern China

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
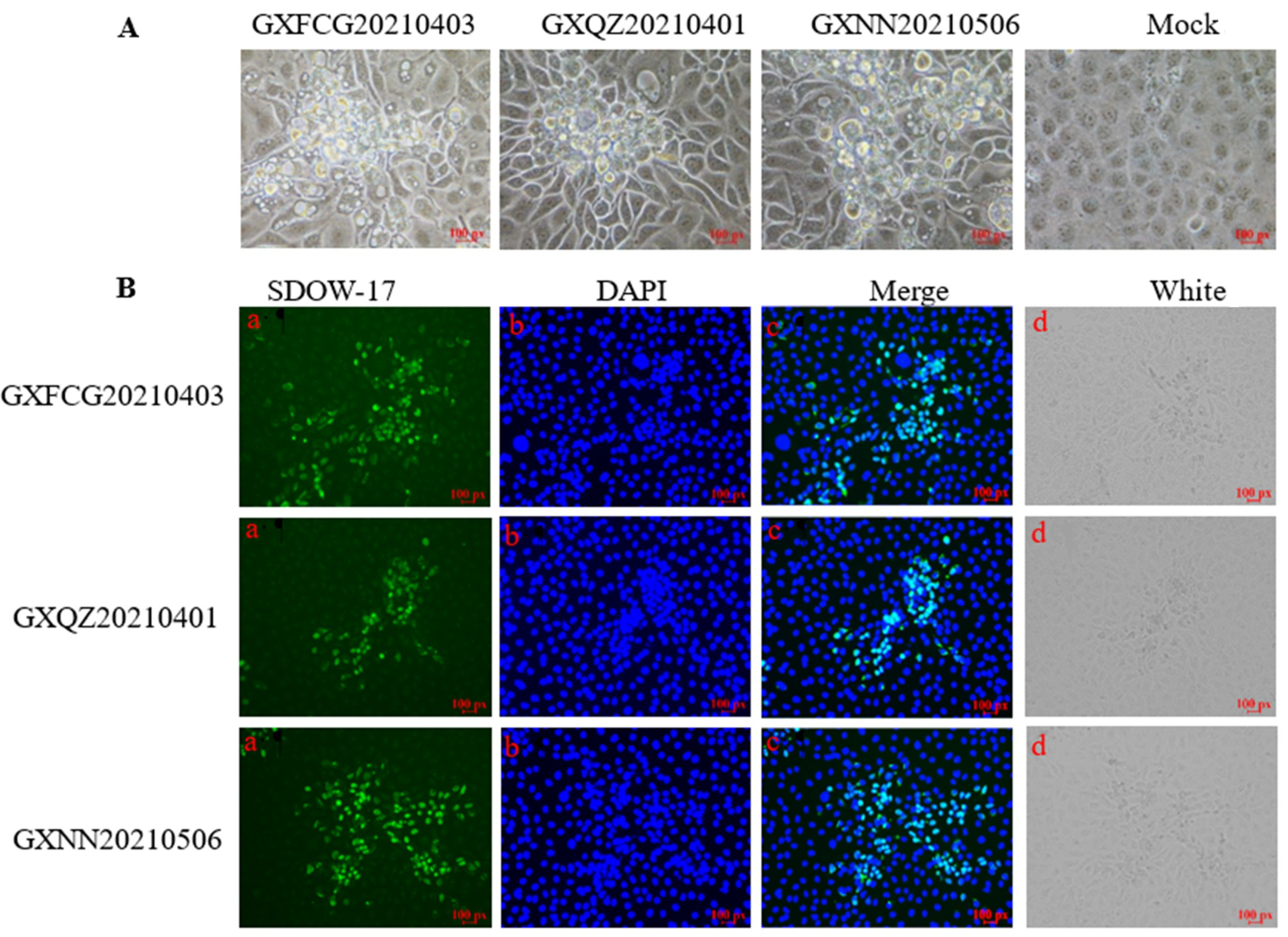
2.1. Clinical Samples, Cells and Virus Isolation
2.2. Immunofluorescence Assay
2.3. Viral RNA Extraction, Reverse Transcription and Complete Genome Determination
2.4. Sequence Comparison and Evolutionary Analysis
2.5. Recombination Analysis
2.6. Pathogenicity of the Recombinant PRRSV Strain in Piglets
2.7. Hematoxylin-Eosin and Immunohistochemistry
2.8. Statistical Analysis
3. Results
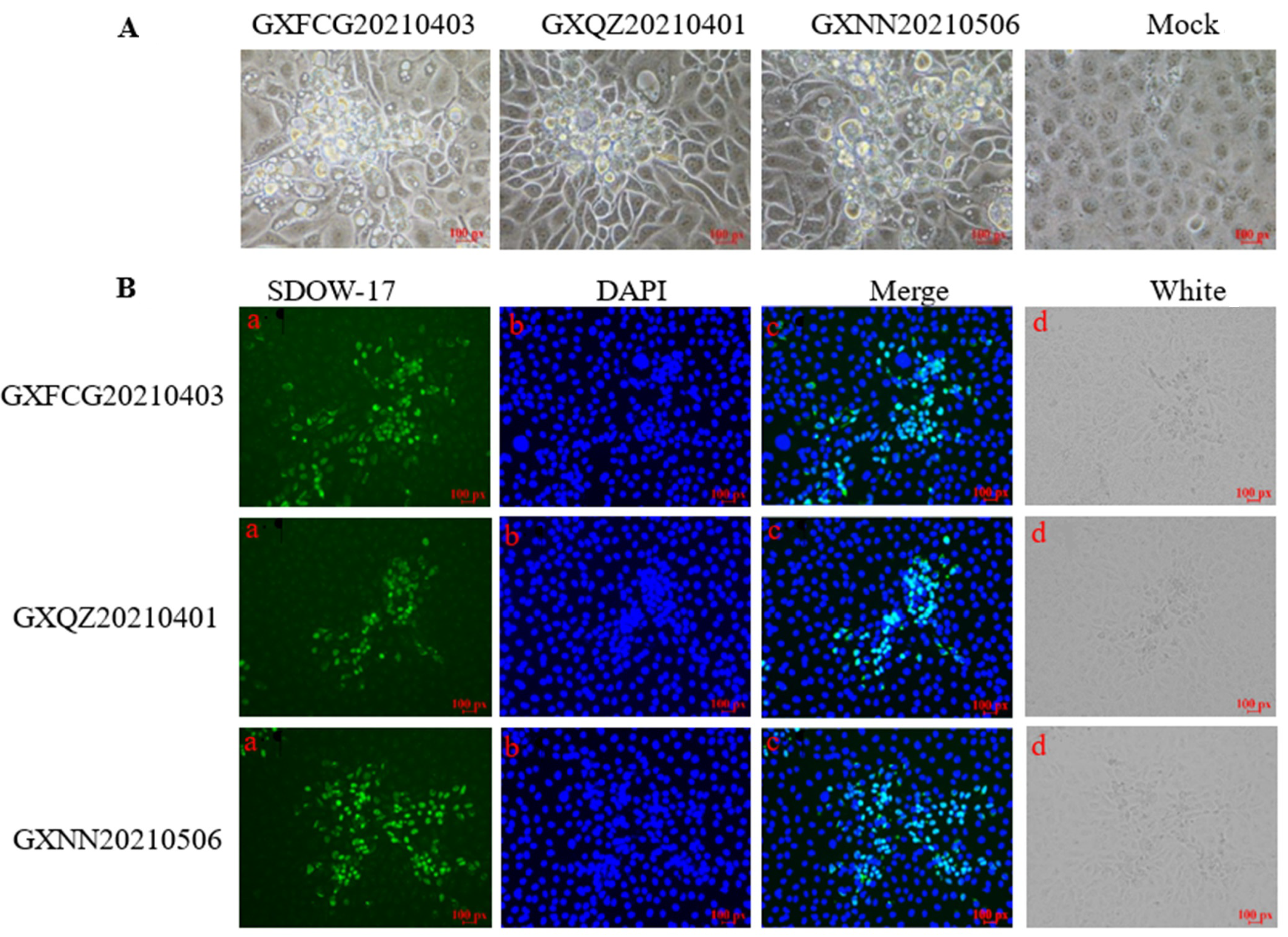
3.1. Isolation and Identification of PRRSV Strains
3.2. Complete Genome Sequence of PRRSV Strains
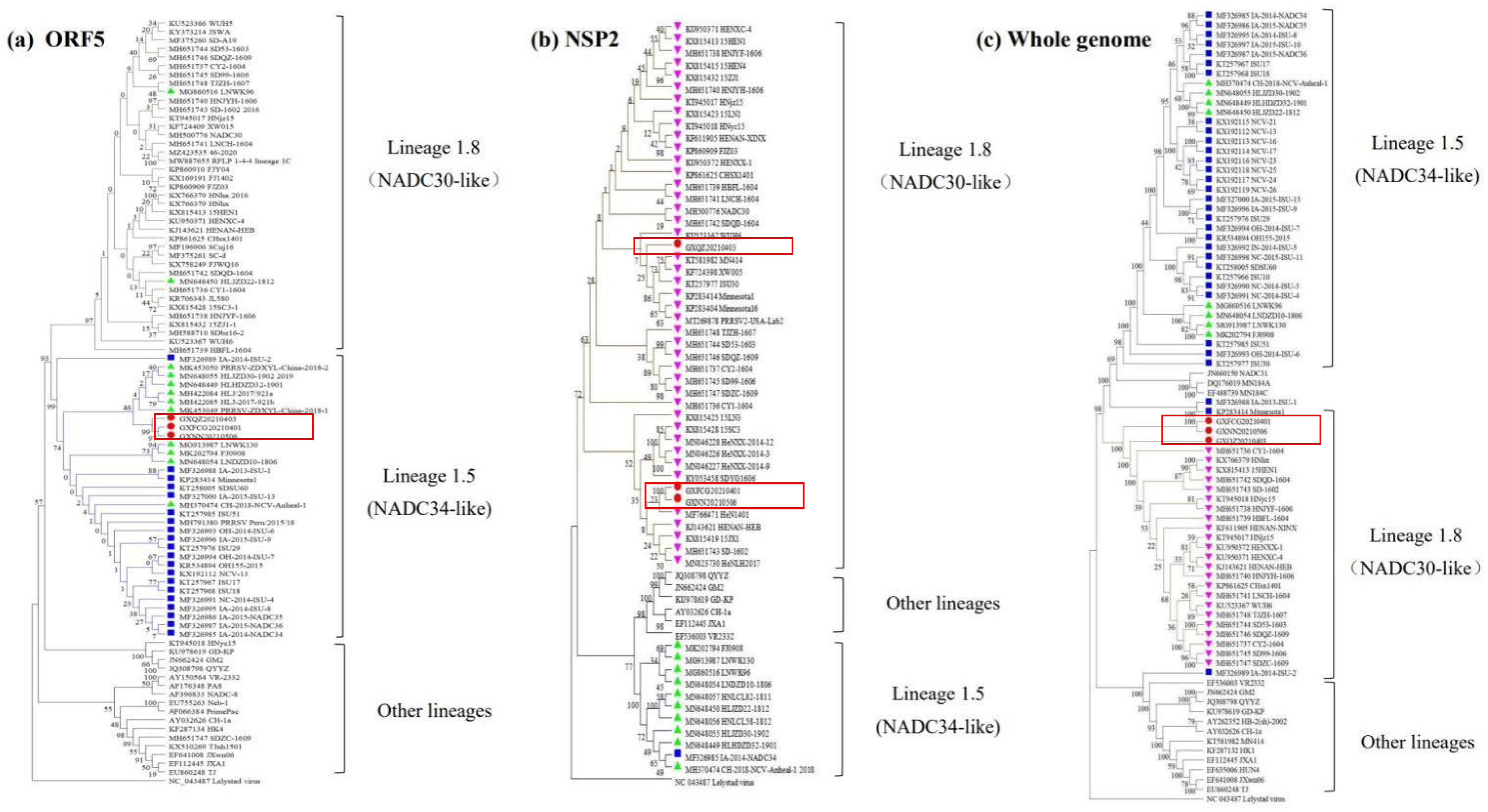
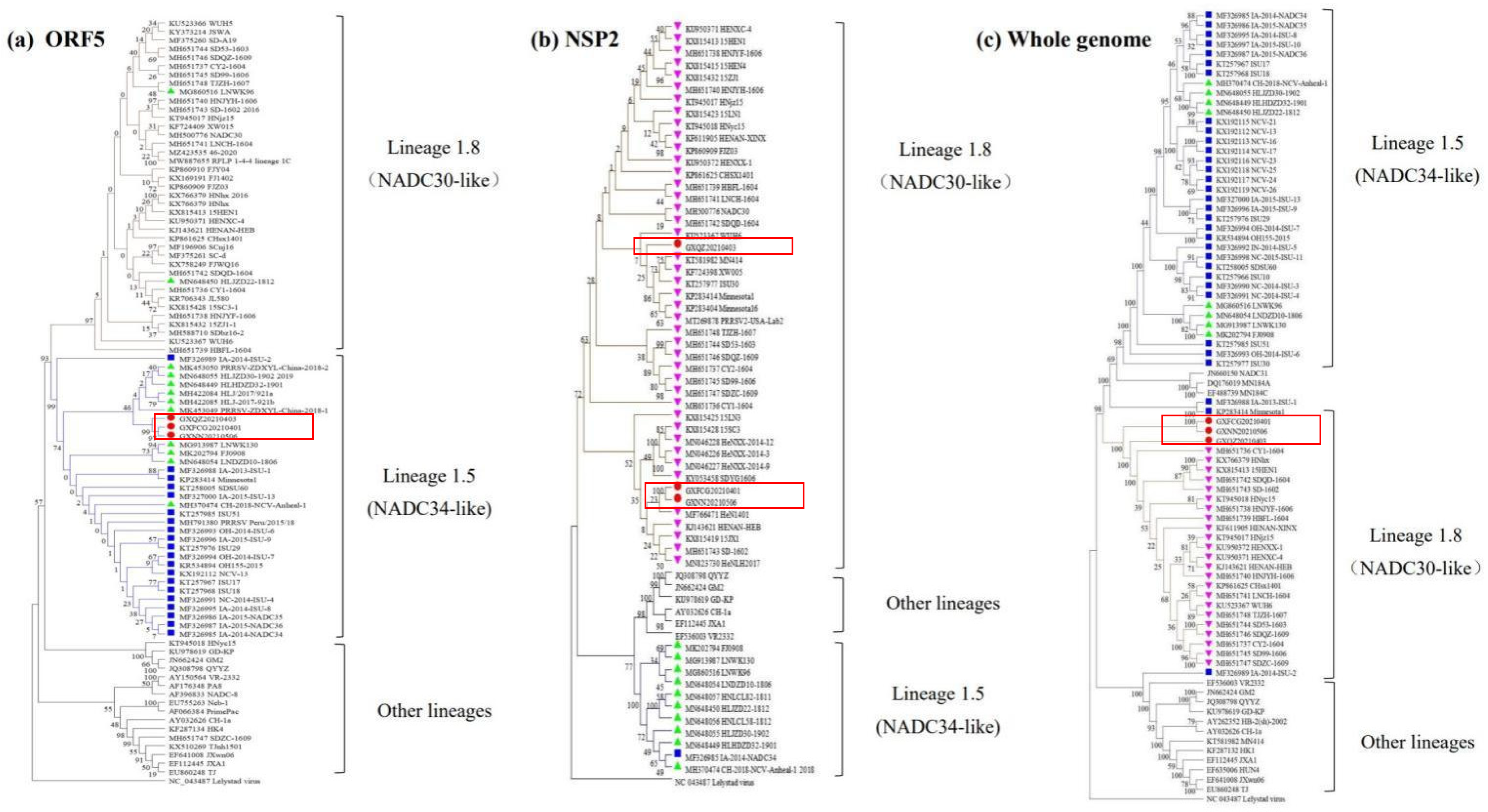
3.3. Phylogenetic Tree
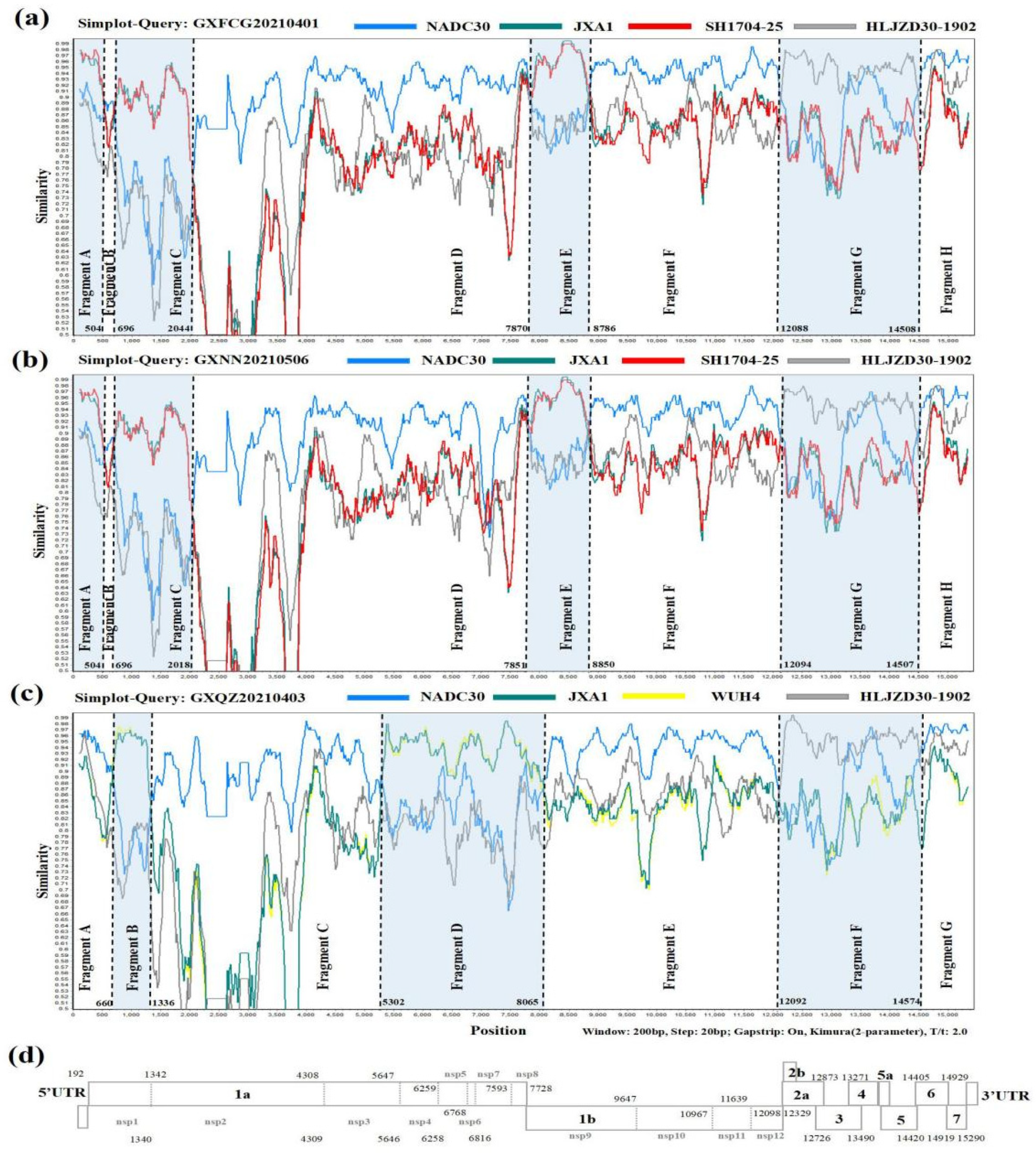
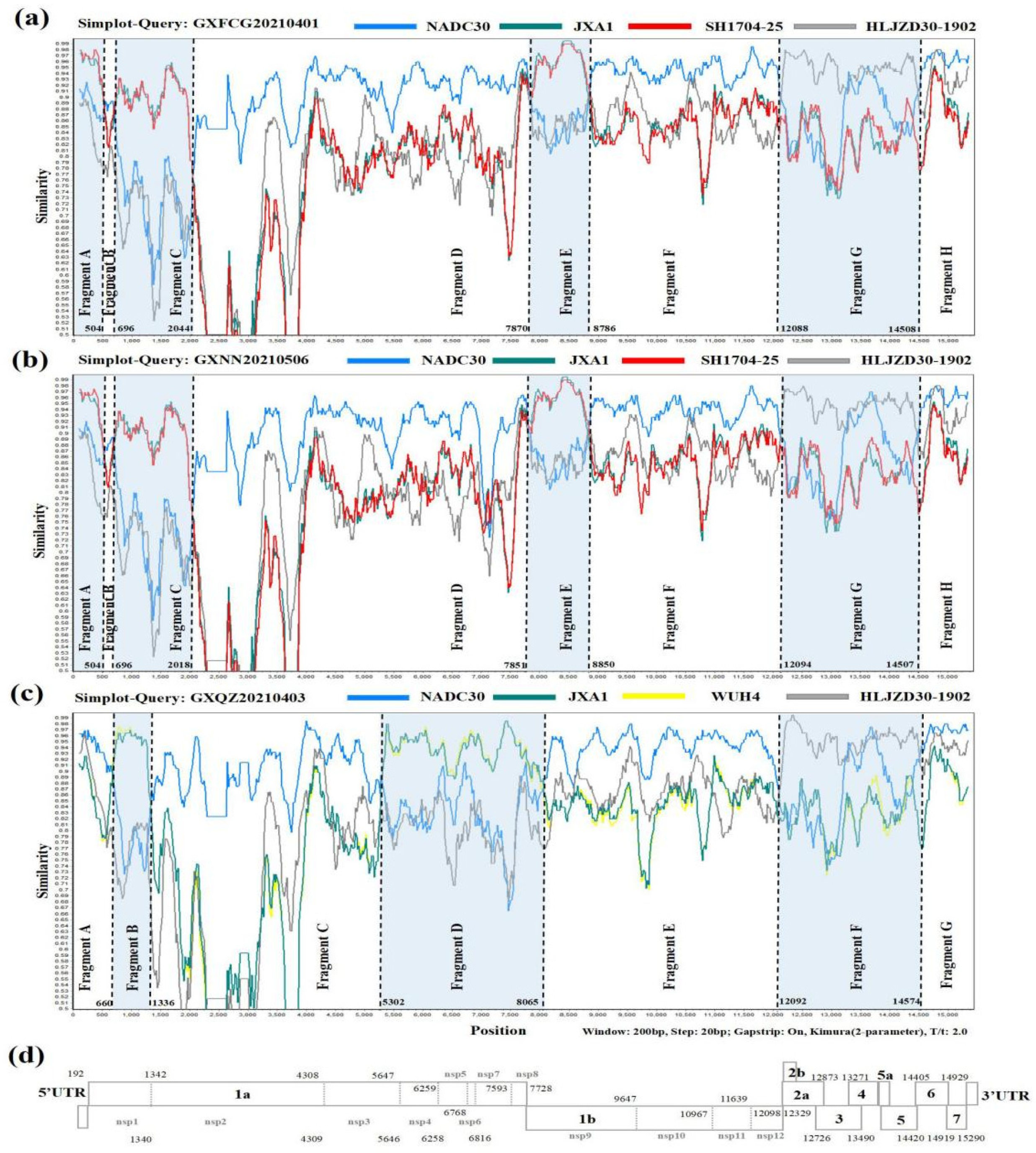
3.4. Recombination Analysis
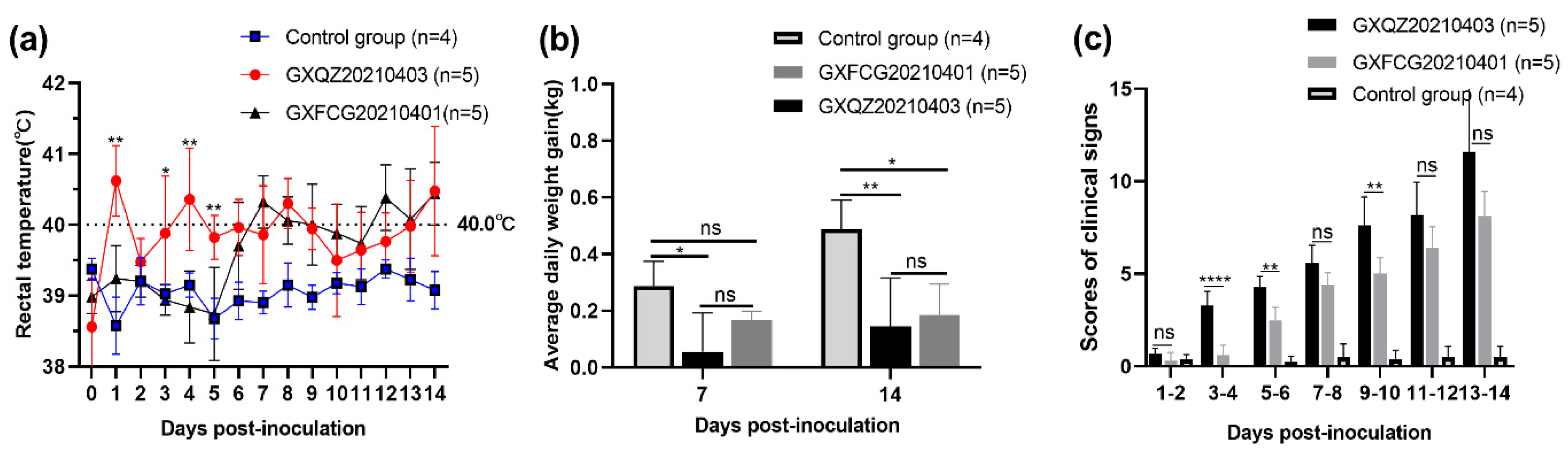
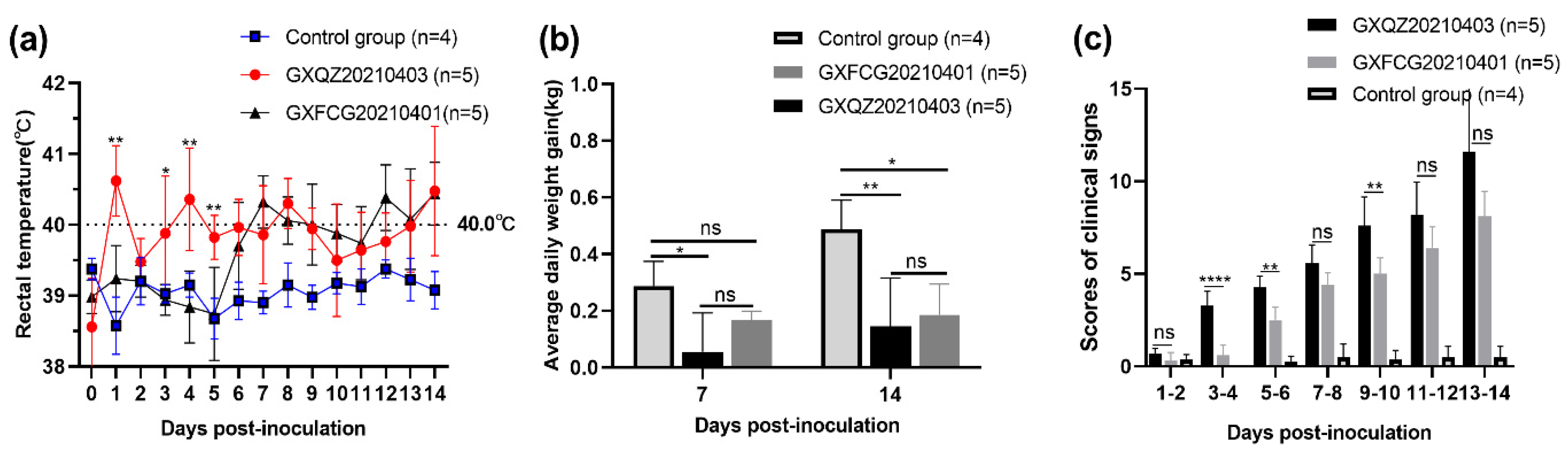
3.5. Observations of Clinical Signs
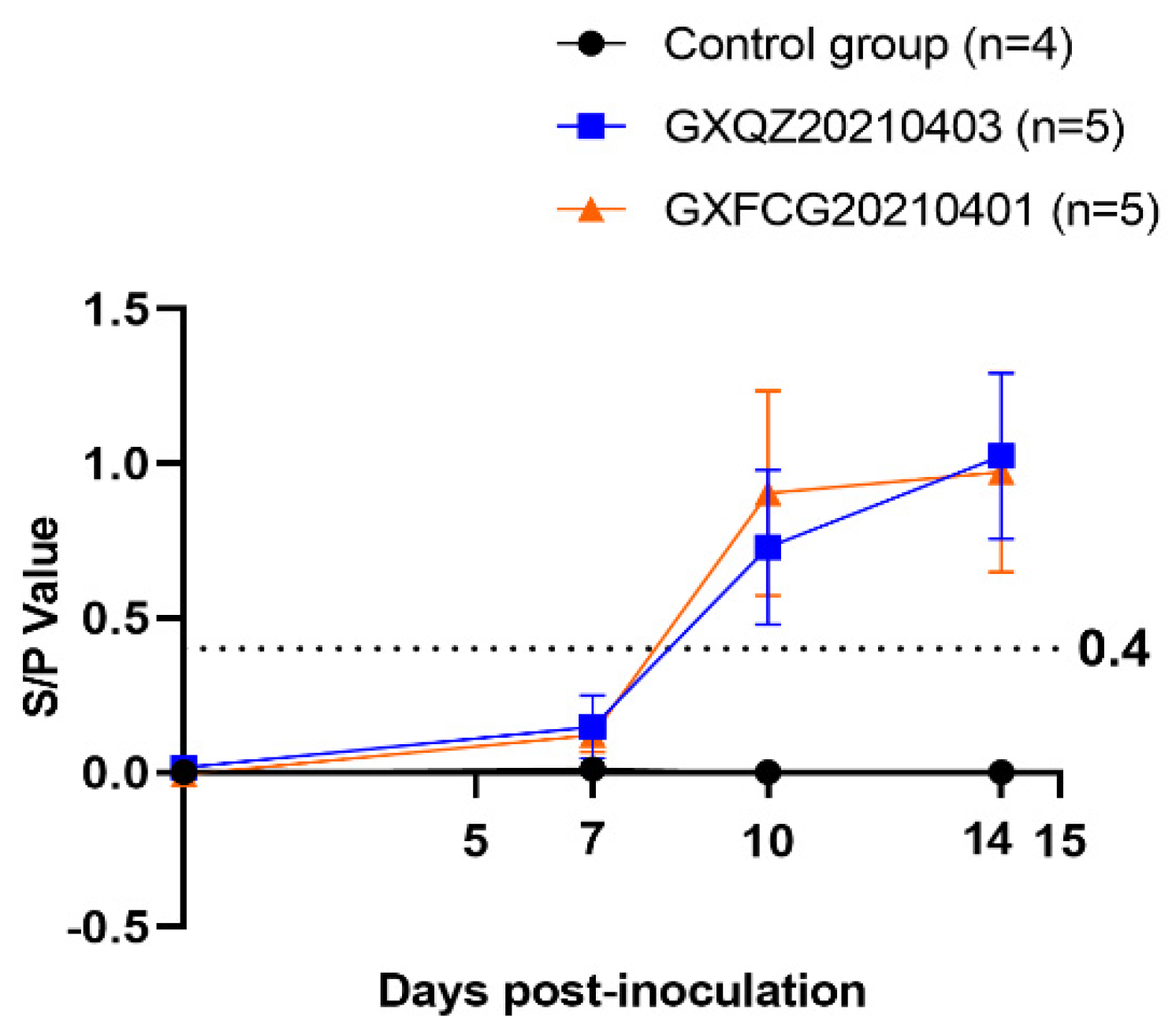
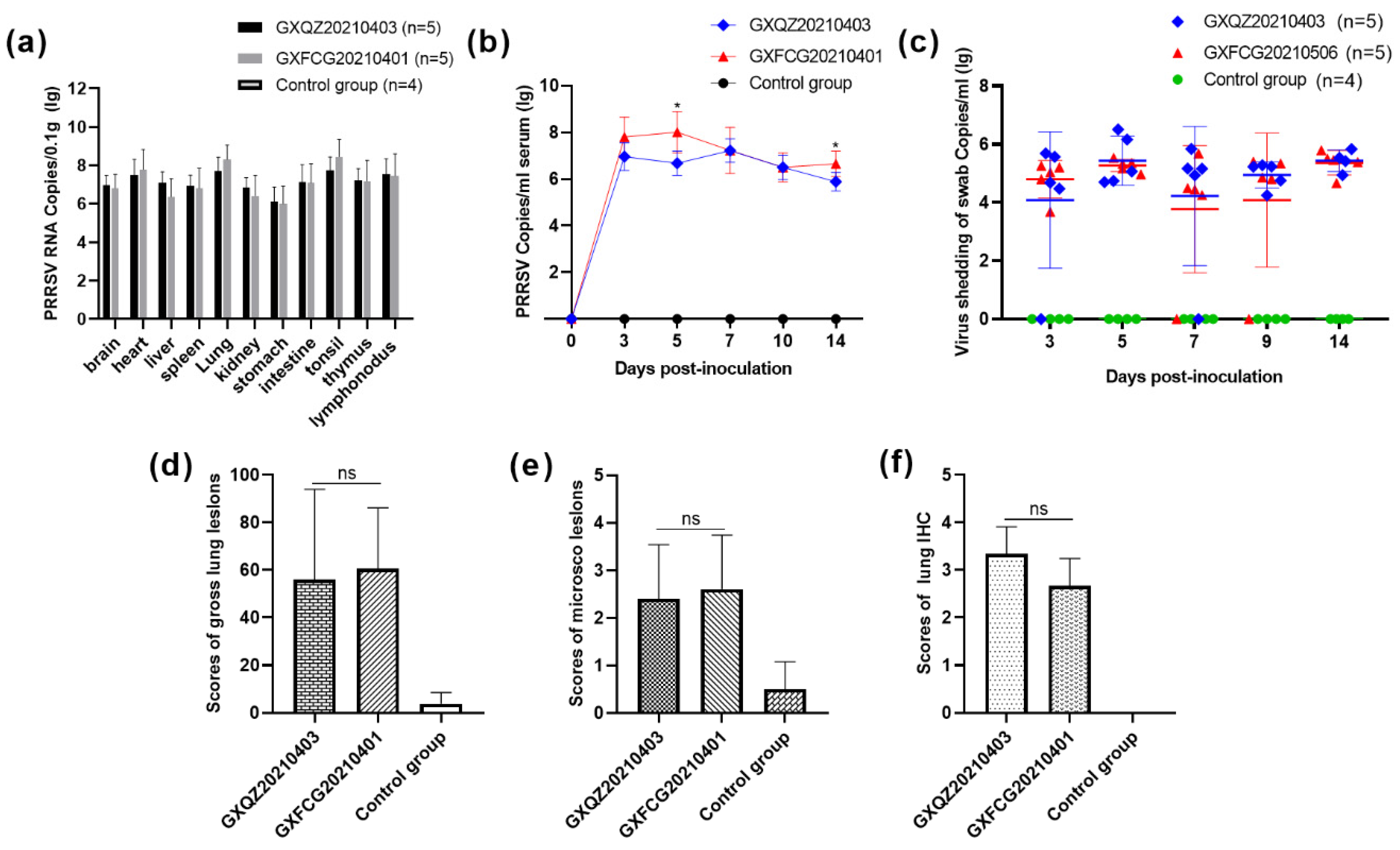
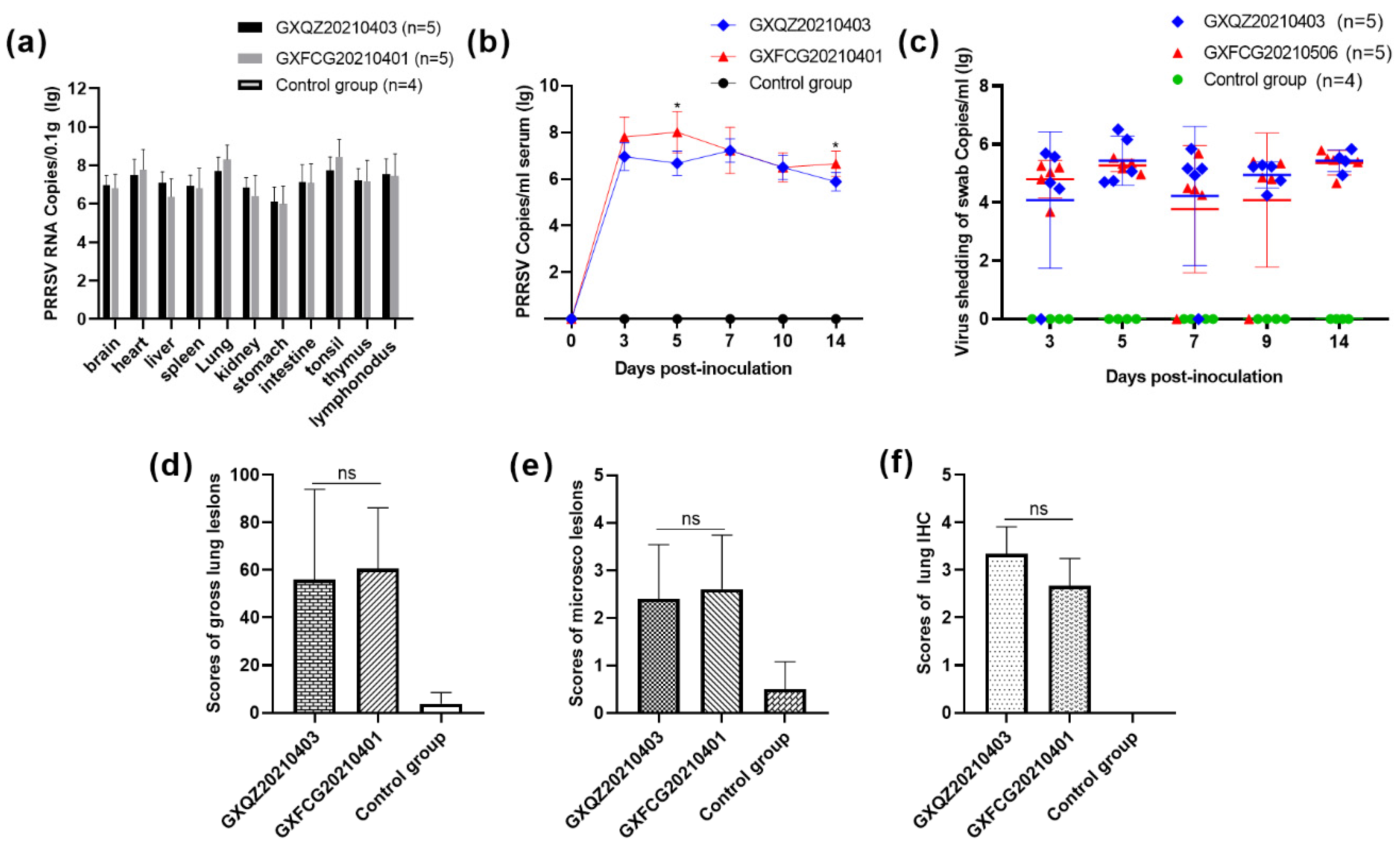
3.6. Antibody Levels, Viremia and Virus Load in Tissues and Swabs
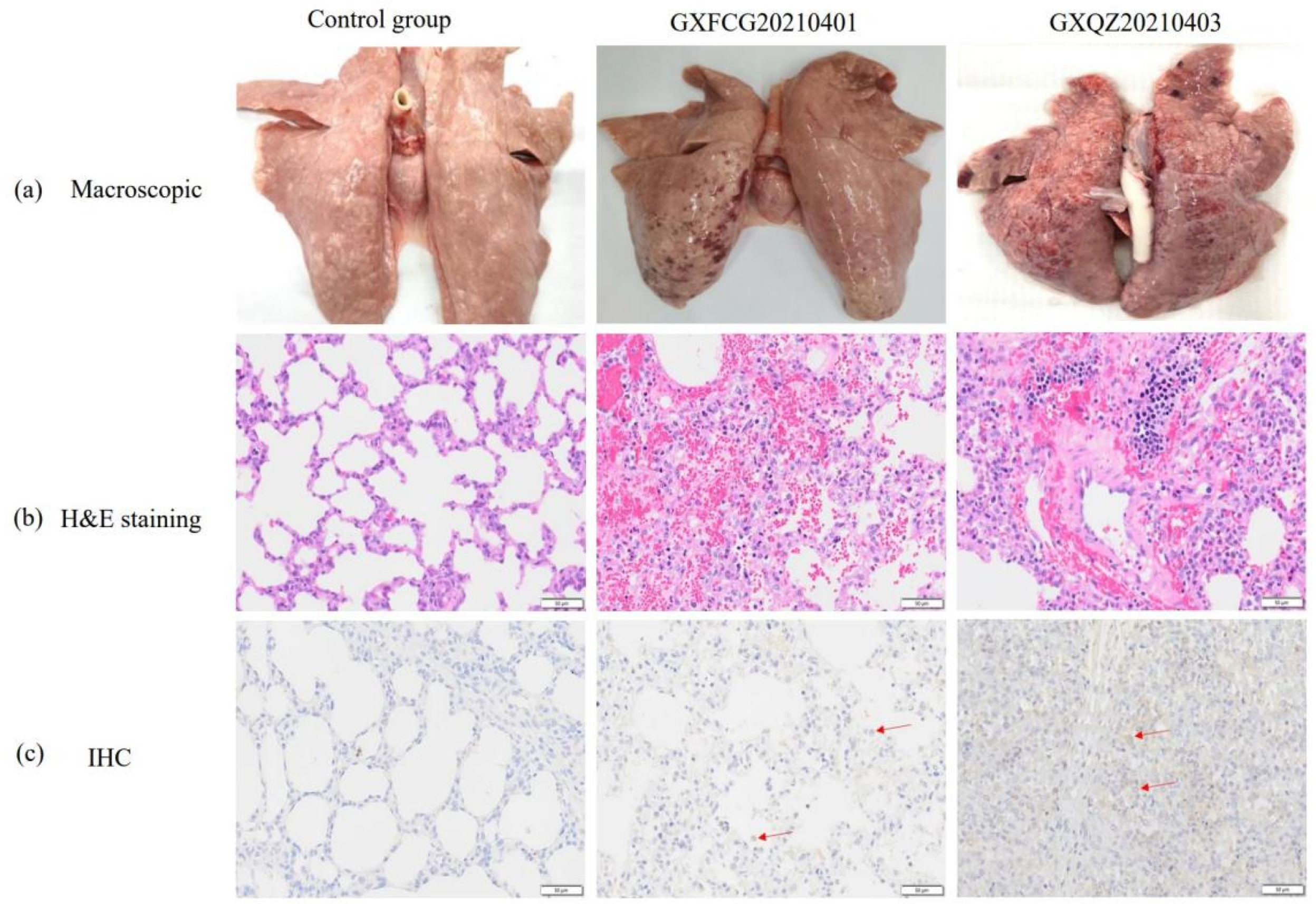
3.7. Macroscopic and Histopathological Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Ethics Approval
References
- Cavanagh, D. Nidovirales: A new order comprising Coronaviridae and Arteriviridae. Arch. Virol. 1997, 142, 629–633. [Google Scholar]
- Meulenberg, J.J. PRRSV, the virus. Vet. Res. 2000, 31, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Dokland, T. The structural biology of PRRSV. Virus Res. 2010, 154, 86–97. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Snijder, E.J. The PRRSV replicase: Exploring the multifunctionality of an intriguing set of nonstructural proteins. Virus Res. 2010, 154, 61–76. [Google Scholar] [CrossRef] [PubMed]
- Firth, A.E.; Zevenhoven-Dobbe, J.C.; Wills, N.M.; Go, Y.Y.; Balasuriya, U.B.R.; Atkins, J.F.; Snijder, E.J.; Posthuma, C.C. Discovery of a small arterivirus gene that overlaps the GP5 coding sequence and is important for virus production. J. Gen. Virol. 2011, 92, 1097–1106. [Google Scholar] [CrossRef]
- Kappes, M.A.; Faaberg, K.S. PRRSV structure, replication and recombination: Origin of phenotype and genotype diversity. Virology 2015, 479, 475–486. [Google Scholar] [CrossRef]
- Nelsen, C.J.; Murtaugh, M.P.; Faaberg, K.S. Porcine reproductive and respiratory syndrome virus comparison: Divergent evolution on two continents. J. Virol. 1999, 73, 270–280. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.H.; Chen, X.X.; Li, R.; Qiao, S.L.; Zhang, G.P. The prevalent status and genetic diversity of porcine reproductive and respiratory syndrome virus in China: A molecular epidemiological perspective. Virol. J. 2018, 15, 2. [Google Scholar] [CrossRef]
- Shi, M.; Lam, T.T.Y.; Hon, C.C.; Hui, R.K.H.; Faaberg, K.S.; Wennblom, T.; Murtaugh, M.P.; Stadejek, T.; Leung, F.C.C. Molecular epidemiology of PRRSV: A phylogenetic perspective. Virus Res. 2010, 154, 7–17. [Google Scholar] [CrossRef]
- Shi, M.; Lam, T.T.Y.; Hon, C.C.; Murtaugh, M.P.; Davies, P.R.; Hui, R.K.H.; Li, J.; Wong, L.T.W.; Yip, C.W.; Jiang, J.W.; et al. Phylogeny-Based Evolutionary, Demographical, and Geographical Dissection of North American Type 2 Porcine Reproductive and Respiratory Syndrome Viruses. J. Virol. 2010, 84, 8700–8711. [Google Scholar] [CrossRef]
- Guo, B. Isolation and Identification of Porcine Reproductory and Respiratory Syndrome (PRRS) Virus from aborted fetuses suspected of PRRS. Chin. J. Anim. Poult. Infect. Dis. 1996, 2, 117–124. [Google Scholar]
- Tian, K.; Yu, X.; Zhao, T.; Feng, Y.; Cao, Z.; Wang, C.; Hu, Y.; Chen, X.; Hu, D.; Tian, X.; et al. Emergence of fatal PRRSV variants: Unparalleled outbreaks of atypical PRRS in China and molecular dissection of the unique hallmark. PLoS ONE 2007, 2, e526. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.J.; Hao, X.F.; Tian, Z.J.; Tong, G.Z.; Yoo, D.; An, T.Q.; Zhou, T.; Li, G.X.; Qiu, H.J.; Wei, T.C.; et al. Highly virulent porcine reproductive and respiratory syndrome virus emerged in China. Transbound. Emerg. Dis. 2008, 55, 152–164. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.X.; Zhu, W.J.; Chen, Y.; Wei, C.Y.; Zhou, P.; Zhang, M.Z.; Huang, Z.; Sun, L.; Su, S.; Zhang, G.H. Molecular epidemiology of PRRSV in South China from 2007 to 2011 based on the genetic analysis of ORF5. Microb. Pathog. 2013, 63, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Ye, C.; Chang, X.B.; Jiang, C.G.; Wang, S.J.; Cai, X.H.; Tong, G.Z.; Tian, Z.J.; Shi, M.; An, T.Q. Importation and Recombination Are Responsible for the Latest Emergence of Highly Pathogenic Porcine Reproductive and Respiratory Syndrome Virus in China. J. Virol. 2015, 89, 10712–10716. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Wang, Z.; Ding, Y.; Ge, X.; Guo, X.; Yang, H. NADC30-like Strain of Porcine Reproductive and Respiratory Syndrome Virus, China. Emerg. Infect. Dis. 2015, 21, 2256–2257. [Google Scholar] [CrossRef]
- Bai, X.; Wang, Y.; Xu, X.; Sun, Z.; Xiao, Y.; Ji, G.; Li, Y.; Tan, F.; Li, X.; Tian, K. Commercial vaccines provide limited protection to NADC30-like PRRSV infection. Vaccine 2016, 34, 5540–5545. [Google Scholar] [CrossRef]
- Zhou, L.; Yang, B.; Xu, L.; Jin, H.; Ge, X.; Guo, X.; Han, J.; Yang, H. Efficacy evaluation of three modified-live virus vaccines against a strain of porcine reproductive and respiratory syndrome virus NADC30-like. Vet. Microbiol. 2017, 207, 108–116. [Google Scholar] [CrossRef]
- Alkhamis, M.A.; Perez, A.M.; Murtaugh, M.P.; Wang, X.; Morrison, R.B. Applications of Bayesian Phylodynamic Methods in a Recent U.S. Porcine Reproductive and Respiratory Syndrome Virus Outbreak. Front. Microbiol. 2016, 7, 67. [Google Scholar] [CrossRef]
- Van Geelen, A.G.M.; Anderson, T.K.; Lager, K.M.; Das, P.B.; Otis, N.J.; Montiel, N.A.; Miller, L.C.; Kulshreshtha, V.; Buckley, A.C.; Brockmeier, S.L.; et al. Porcine reproductive and respiratory disease virus: Evolution and recombination yields distinct ORF5 RFLP 1-7-4 viruses with individual pathogenicity. Virology 2018, 513, 168–179. [Google Scholar] [CrossRef]
- Ramirez, M.; Bauermann, F.V.; Navarro, D.; Rojas, M.; Manchego, A.; Nelson, E.A.; Diel, D.G.; Rivera, H. Detection of porcine reproductive and respiratory syndrome virus (PRRSV) 1-7-4-type strains in Peru. Transbound Emerg. Dis. 2019, 66, 1107–1113. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wei, C.; Lin, Z.; Xia, W.; Ma, Y.; Dai, A.; Yang, X. Full genome sequence analysis of a 1-7-4-like PRRSV strain in Fujian Province, China. PeerJ 2019, 7, e7859. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Xu, H.; Zhao, J.; Leng, C.; Xiang, L.; Li, C.; Fu, J.; Tang, Y.D.; Peng, J.; Wang, Q.; et al. Pathogenicity of NADC34-like PRRSV HLJDZD32-1901 isolated in China. Vet. Microbiol. 2020, 246, 108727. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.; Ha, Z.; Nan, F.; Zhang, Y.; Zhang, H.; Li, J.; Zhang, P.; Han, J.; Zhang, H.; Zhuang, X.; et al. Characterization of porcine reproductive and respiratory syndrome virus (ORF5 RFLP 1-7-4 viruses) in northern China. Microb. Pathog. 2020, 140, 103941. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.L.; Zhang, W.L.; Xiang, L.R.; Leng, C.L.; Tian, Z.J.; Tang, Y.D.; Cai, X.H. Emergence of novel porcine reproductive and respiratory syndrome viruses (ORF5 RFLP 1-7-4 viruses) in China. Vet. Microbiol. 2018, 222, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.Z.; Ha, Z.; Zhang, H.; Zhang, Y.; Xie, Y.B.; Zhang, H.; Nan, F.L.; Wang, Z.; Zhang, P.; Xu, W.; et al. Pathogenicity of porcine reproductive and respiratory syndrome virus (ORF5 RFLP 1-7-4 viruses) in China. Transbound. Emerg. Dis. 2020, 67, 2065–2072. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Jiang, P.; Song, Z.; Lv, L.; Li, L.; Bai, J. Pathogenicity and antigenicity of a novel NADC30-like strain of porcine reproductive and respiratory syndrome virus emerged in China. Vet. Microbiol. 2016, 197, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.F.; Liu, Y.; Yang, J.; Li, W.Z.; Yu, X.X.; Wang, S.Y.; Li, L.A.; Yu, H. Recombination between NADC34-like and QYYZ-like strain of porcine reproductive and respiratory syndrome virus with high pathogenicity for piglets in China. Transbound. Emerg. Dis. 2022. [Google Scholar] [CrossRef]
- Xu, H.; Li, C.; Li, W.S.; Zhao, J.; Gong, B.J.; Sun, Q.; Tang, Y.D.; Xiang, L.R.; Leng, C.L.; Peng, J.M.; et al. Novel characteristics of Chinese NADC34-like PRRSV during 2020–2021. Transbound. Emerg. Dis. 2022. [Google Scholar] [CrossRef]
- Zhang, H.; Leng, C.; Ding, Y.; Zhai, H.; Li, Z.; Xiang, L.; Zhang, W.; Liu, C.; Li, M.; Chen, J.; et al. Characterization of newly emerged NADC30-like strains of porcine reproductive and respiratory syndrome virus in China. Arch. Virol. 2019, 164, 401–411. [Google Scholar] [CrossRef]
- Zhang, X.; Zhu, Z.; Yang, F.; Cao, W.; Tian, H.; Zhang, K.; Zheng, H.; Liu, X. Review of Seneca Valley Virus: A Call for Increased Surveillance and Research. Front. Microbiol. 2018, 9, 940. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Wu, J.; Shang, Y.; Cheng, Y.; Liu, X. The development of a rapid SYBR one step real-time RT-PCR for detection of Porcine Reproductive and Respiratory Syndrome Virus. Virol. J. 2010, 7, 90. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Yan, Y.; Shi, M.; Liu, H.Z.; Zhang, H.L.; Yang, Y.B.; Huang, X.Y.; Gauger, P.C.; Zhang, J.; Zhang, Y.H.; et al. Phylogenetics, Genomic Recombination, and NSP2 Polymorphic Patterns of Porcine Reproductive and Respiratory Syndrome Virus in China and the United States in 2014–2018. J. Virol. 2020, 94, e01813-19. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Han, Q.; Zhang, L.; Zhang, Z.; Wu, Y.; Shen, H.; Jiang, P. Emergence of mosaic recombinant strains potentially associated with vaccine JXA1-R and predominant circulating strains of porcine reproductive and respiratory syndrome virus in different provinces of China. Virol. J. 2017, 14, 67. [Google Scholar] [CrossRef] [PubMed]
- Tian, K. NADC30-Like Porcine Reproductive and Respiratory Syndrome in China. Open Virol. J. 2017, 11, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Ansari, I.H.; Kwon, B.; Osorio, F.A.; Pattnaik, A.K. Influence of N-linked glycosylation of porcine reproductive and respiratory syndrome virus GP5 on virus infectivity, antigenicity, and ability to induce neutralizing antibodies. J. Virol. 2006, 80, 3994–4004. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.Z.; Lin, T.; Sun, L.C.; Li, Y.H.; Wang, X.M.; Gao, F.; Liu, R.X.; Chen, C.Y.; Tong, G.Z.; Yuan, S.S. N-Linked Glycosylation of GP5 of Porcine Reproductive and Respiratory Syndrome Virus Is Critically Important for Virus Replication In Vivo. J. Virol. 2012, 86, 9941–9951. [Google Scholar] [CrossRef]
- Zhu, Z.; Yuan, L.; Hu, D.; Lian, Z.; Yao, X.; Liu, P.; Li, X. Isolation and genomic characterization of a Chinese NADC34-like PRRSV isolated from Jiangsu province. Transbound. Emerg. Dis. 2021, 69, e1015–e1027. [Google Scholar] [CrossRef]
- Popescu, L.N.; Trible, B.R.; Chen, N.H.; Rowland, R.R.R. GP5 of porcine reproductive and respiratory syndrome virus (PRRSV) as a target for homologous and broadly neutralizing antibodies. Vet. Microbiol. 2017, 209, 90–96. [Google Scholar] [CrossRef]
- Xu, H.; Song, S.; Zhao, J.; Leng, C.; Fu, J.; Li, C.; Tang, Y.D.; Xiang, L.; Peng, J.; Wang, Q.; et al. A potential endemic strain in China: NADC34-like porcine reproductive and respiratory syndrome virus. Transbound. Emerg. Dis. 2020, 67, 1730–1738. [Google Scholar] [CrossRef]
- Wei, C.; Dai, A.; Fan, J.; Li, Y.; Chen, A.; Zhou, X.; Luo, M.; Yang, X.; Liu, J. Efficacy of Type 2 PRRSV vaccine against challenge with the Chinese lineage 1 (NADC30-like) PRRSVs in pigs. Sci. Rep. 2019, 9, 10781. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, J.; Yang, J.; Zeng, H.; Guo, L.; Ren, S.; Sun, W.; Chen, Z.; Cong, X.; Shi, J.; et al. Emergence of Different Recombinant Porcine Reproductive and Respiratory Syndrome Viruses, China. Sci. Rep. 2018, 8, 4118. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Kang, R.; Xie, B.; Tian, Y.; Wu, X.; Lv, X.; Yang, X.; Wang, H. Identification of a Novel Recombinant Type 2 Porcine Reproductive and Respiratory Syndrome Virus in China. Viruses 2018, 10, 151. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Predicted N-Glycosylation Sites of the PRRSV GP5 Protein | ||
|---|---|---|
| Strains | N-Glycosylation Sites | Numbers |
| GXFCG20210401 | N32, N33, N44, N51, N57 | 5 |
| GXQZ20210403 | N32, N33, N44, N51, N57 | 5 |
| GXNN20210506 | N32, N33, N44, N51, N57 | 5 |
| IA/2014/NADC34 | N32, N33, N44, N51, N57 | 5 |
| NADC30 | N34, N44, N51 | 3 |
| CHsx1401 | N33, N44, N51 | 3 |
| JXA1 | N30, N34, N35, N44, N51 | 5 |
| QYYZ | N34, N44, N51 | 3 |
| GM-2 | N33, N44, N51 | 3 |
| CH-1a | N34, N44, N51 | 3 |
| VR2332 | N34, N44, N51 | 3 |
| FJ0908 | N32, N33, N44, N51 | 4 |
| LNWK96 | N33, N34, N44, N51 | 4 |
| LNWK130 | N32, N34, N44, N51 | 4 |
| HLHDZD32-1901 | N32, N33, N44, N51 | 4 |
| HLJZD22-1812 | N32, N33, N44, N51 | 4 |
| HLJZD30-1902 | N30, N33, N44, N51 | 4 |
| LNDZD10-1806 | N30, N33, N44, N51 | 4 |
| PRRSV-ZDXYL-China-2018-1 | N32, N33, N44, N51, N57 | 5 |
| PRRSV-ZDXYL-China-2018-2 | N32, N33, N44, N51 | 4 |
| CH-2018-NCV-Anheal-1 | N32, N33, N34, N44, N51, N57 | 6 |
| JS2021NADC34 | N32, N33, N44, N51 | 4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Zhang, K.; Mo, Q.; Chen, G.; Lv, J.; Huang, J.; Pang, Y.; Wang, H.; Liu, W.; Huang, K.; et al. The Emergence and Pathogenesis of Recombinant Viruses Associated with NADC34-like Strains and the Predominant Circulating Strains of Porcine Reproductive and Respiratory Syndrome Virus in Southern China. Viruses 2022, 14, 1695. https://doi.org/10.3390/v14081695
Wang X, Zhang K, Mo Q, Chen G, Lv J, Huang J, Pang Y, Wang H, Liu W, Huang K, et al. The Emergence and Pathogenesis of Recombinant Viruses Associated with NADC34-like Strains and the Predominant Circulating Strains of Porcine Reproductive and Respiratory Syndrome Virus in Southern China. Viruses. 2022; 14(8):1695. https://doi.org/10.3390/v14081695
Chicago/Turabian StyleWang, Xindong, Kang Zhang, Qingrong Mo, Guochang Chen, Jing Lv, Jing Huang, Yanli Pang, Hao Wang, Wenbo Liu, Kai Huang, and et al. 2022. "The Emergence and Pathogenesis of Recombinant Viruses Associated with NADC34-like Strains and the Predominant Circulating Strains of Porcine Reproductive and Respiratory Syndrome Virus in Southern China" Viruses 14, no. 8: 1695. https://doi.org/10.3390/v14081695
APA StyleWang, X., Zhang, K., Mo, Q., Chen, G., Lv, J., Huang, J., Pang, Y., Wang, H., Liu, W., Huang, K., Min, X., Ren, T., Ouyang, K., Chen, Y., Huang, W., & Wei, Z. (2022). The Emergence and Pathogenesis of Recombinant Viruses Associated with NADC34-like Strains and the Predominant Circulating Strains of Porcine Reproductive and Respiratory Syndrome Virus in Southern China. Viruses, 14(8), 1695. https://doi.org/10.3390/v14081695