
Detection of Equus Caballus Papillomavirus Type-2 in Asymptomatic Italian Horses

 ,
, 
 , , , , , , , ,
, , , , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Selection of Horses for EcPV2 Detection
2.2. EcPV2 DNA Real-Time PCR
2.3. NGS and Bioinformatics Analysis
2.4. RNA Extraction and EcPV2 Gene Expression
2.5. Host Gene Expression Study
2.6. Statistical Analysis
3. Results
3.1. Sampled Population
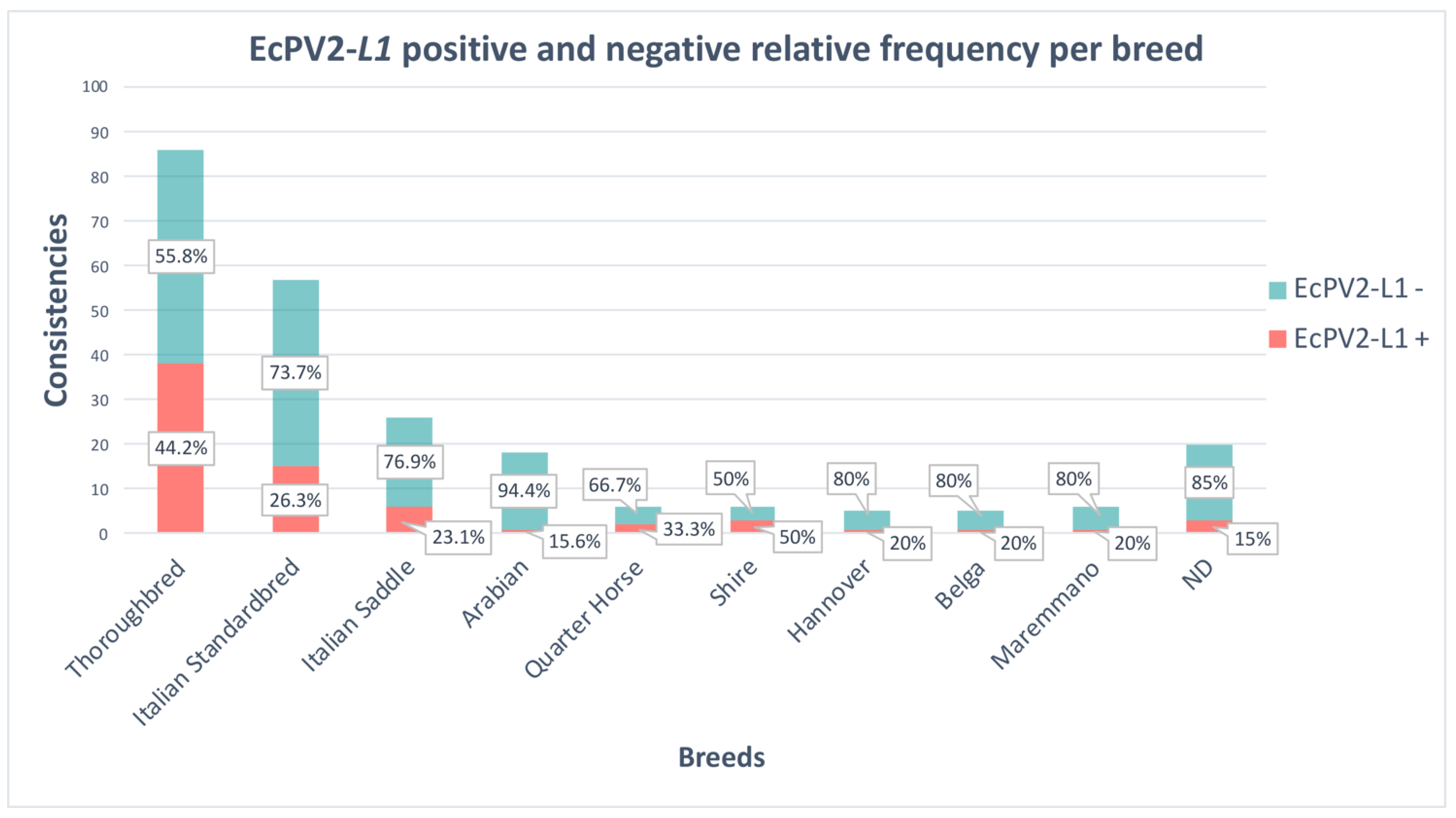
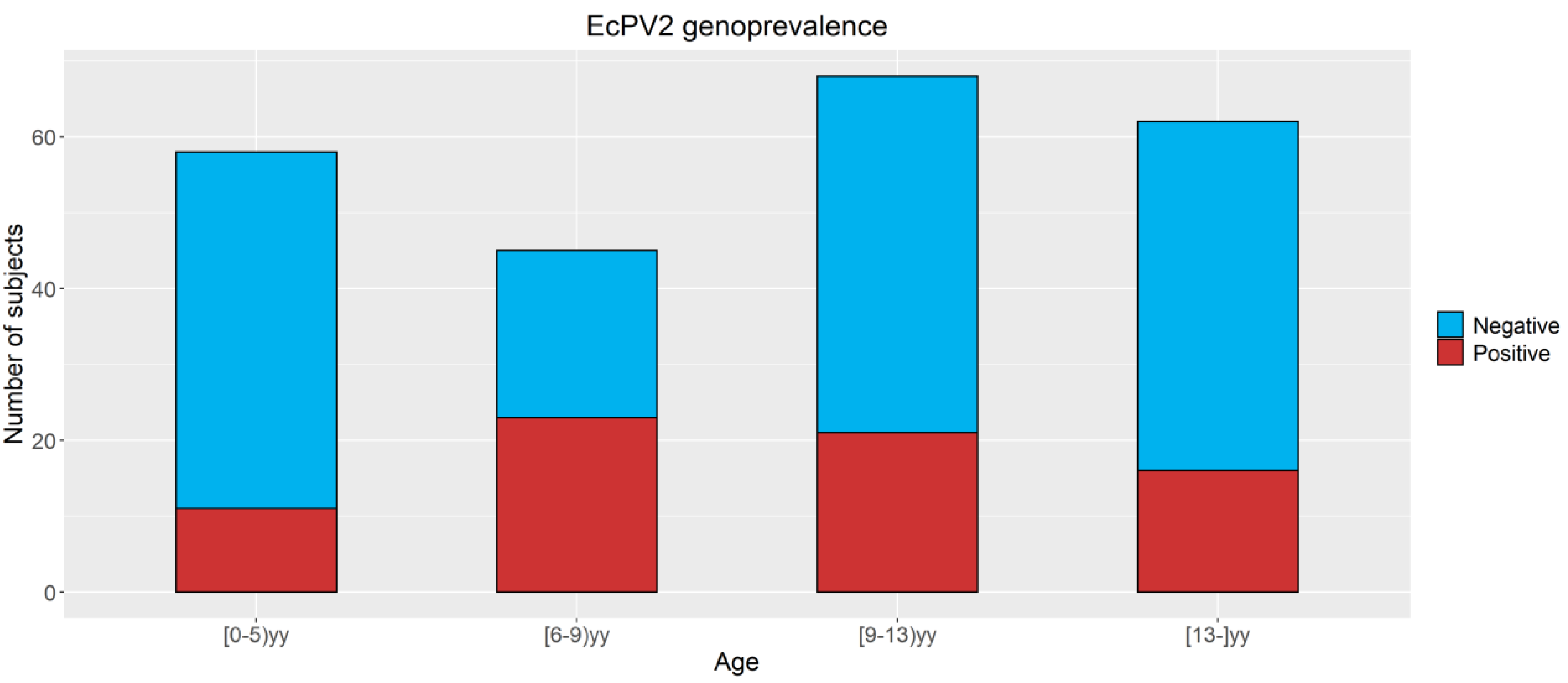
3.2. Detection of EcPV2 DNA
3.3. Detection of EcPV2 DNA in Mares
3.4. NGS and Bioinformatics Analysis
3.5. Detection of Gene Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Doorbar, J. Host Control of Human Papillomavirus Infection and Disease. Best Pract. Res. Clin. Obstet. Gynaecol. 2018, 47, 27–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, J.R.; Fernandez, D.J.; Thornton, S.M.; Skeate, J.G.; Lühen, K.P.; Da Silva, D.M.; Langen, R.; Kast, W.M. Heterotetrameric Annexin A2/S100A10 (A2t) Is Essential for Oncogenic Human Papillomavirus Trafficking and Capsid Disassembly, and Protects Virions from Lysosomal Degradation. Sci. Rep. 2018, 8, 11642. [Google Scholar] [CrossRef]
- Ramsauer, A.S.; Wachoski-Dark, G.L.; Fraefel, C.; Tobler, K.; Brandt, S.; Knight, C.G.; Favrot, C.; Grest, P. Paving the Way for More Precise Diagnosis of EcPV2-Associated Equine Penile Lesions. BMC Vet. Res. 2019, 15, 356. [Google Scholar] [CrossRef] [Green Version]
- Doorbar, J.; Quint, W.; Banks, L.; Bravo, I.G.; Stoler, M.; Broker, T.R.; Stanley, M.A. The Biology and Life-Cycle of Human Papillomaviruses. Vaccine 2012, 30 (Suppl. 5), F55–F70. [Google Scholar] [CrossRef]
- Araldi, R.P.; Assaf, S.M.R.; de Carvalho, R.F.; de Carvalho, M.A.C.R.; de Souza, J.M.; Magnelli, R.F.; Módolo, D.G.; Roperto, F.P.; de Cassia Stocco, Rita; Beçak, W. Papillomaviruses: A Systematic Review. Genet. Mol. Biol. 2017, 40, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Munday, J.S.; Thomson, N.A.; Luff, J.A. Papillomaviruses in Dogs and Cats. Vet. J. 2017, 225, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Sykora, S.; Brandt, S. Papillomavirus Infection and Squamous Cell Carcinoma in Horses. Vet. J. 2017, 223, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Hibi, H.; Hatama, S.; Obata, A.; Shibahara, T.; Kadota, K. Laryngeal Squamous Cell Carcinoma and Papilloma Associated with Equus caballus Papillomavirus 2 in a Horse. J. Vet. Med. Sci. 2019, 81, 1029–1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porcellato, I.; Modesto, P.; Cappelli, K.; Varello, K.; Peletto, S.; Brachelente, C.; Martini, I.; Mechelli, L.; Ferrari, A.; Ghelardi, A.; et al. Equus caballus Papillomavirus Type 2 (EcPV2) in Co-Occurring Vulvar and Gastric Lesions of a Pony. Res. Vet. Sci. 2020, 132, 167–171. [Google Scholar] [CrossRef] [PubMed]
- Mecocci, S.; Porcellato, I.; Armando, F.; Mechelli, L.; Brachelente, C.; Pepe, M.; Gialletti, R.; Passeri, B.; Modesto, P.; Ghelardi, A.; et al. Equine Genital Squamous Cell Carcinoma Associated with EcPV2 Infection: RANKL Pathway Correlated to Inflammation and Wnt Signaling Activation. Biology 2021, 10, 244. [Google Scholar] [CrossRef]
- Sykora, S.; Samek, L.; Schönthaler, K.; Palm, F.; Borzacchiello, G.; Aurich, C.; Brandt, S. EcPV-2 Is Transcriptionally Active in Equine SCC but Only Rarely Detectable in Swabs and Semen from Healthy Horses. Vet. Microbiol. 2012, 158, 194–198. [Google Scholar] [CrossRef]
- Bogaert, L.; Willemsen, A.; Vanderstraeten, E.; Bracho, M.A.; De Baere, C.; Bravo, I.G.; Martens, A. EcPV2 DNA in Equine Genital Squamous Cell Carcinomas and Normal Genital Mucosa. Vet. Microbiol. 2012, 158, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Fischer, N.M.; Favrot, C.; Birkmann, K.; Jackson, M.; Schwarzwald, C.C.; Müller, M.; Tobler, K.; Geisseler, M.; Lange, C.E. Serum Antibodies and DNA Indicate a High Prevalence of Equine Papillomavirus 2 (EcPV2) among Horses in Switzerland. Vet. Dermatol. 2014, 25, 210-e54. [Google Scholar] [CrossRef] [PubMed]
- Knight, C.G.; Munday, J.S.; Peters, J.; Dunowska, M. Equine Penile Squamous Cell Carcinomas Are Associated with the Presence of Equine Papillomavirus Type 2 DNA Sequences. Vet. Pathol. 2011, 48, 1190–1194. [Google Scholar] [CrossRef] [PubMed]
- Knight, C.G.; Dunowska, M.; Munday, J.S.; Peters-Kennedy, J.; Rosa, B.V. Comparison of the Levels of Equus caballus Papillomavirus Type 2 (EcPV-2) DNA in Equine Squamous Cell Carcinomas and Non-Cancerous Tissues Using Quantitative PCR. Vet. Microbiol. 2013, 166, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Omran, O.M.; AlSheeha, M. Human Papilloma Virus Early Proteins E6 (HPV16/18-E6) and the Cell Cycle Marker P16 (INK4a) Are Useful Prognostic Markers in Uterine Cervical Carcinomas in Qassim Region—Saudi Arabia. Pathol. Oncol. Res. POR 2015, 21, 157–166. [Google Scholar] [CrossRef]
- Greenwood, S.; Chow-Lockerbie, B.; Epp, T.; Knight, C.; Wachoski-Dark, G.; MacDonald-Dickinson, V.; Wobeser, B. Prevalence and Prognostic Impact of Equus caballus Papillomavirus Type 2 Infection in Equine Squamous Cell Carcinomas in Western Canadian Horses. Vet. Pathol. 2020, 57, 623–631. [Google Scholar] [CrossRef]
- Armando, F.; Mecocci, S.; Orlandi, V.; Porcellato, I.; Cappelli, K.; Mechelli, L.; Brachelente, C.; Pepe, M.; Gialletti, R.; Ghelardi, A.; et al. Investigation of the Epithelial to Mesenchymal Transition (EMT) Process in Equine Papillomavirus-2 (EcPV-2)-Positive Penile Squamous Cell Carcinomas. Int. J. Mol. Sci. 2021, 22, 10588. [Google Scholar] [CrossRef]
- Porcellato, I.; Mecocci, S.; Brachelente, C.; Cappelli, K.; Armando, F.; Tognoloni, A.; Chiaradia, E.; Stefanetti, V.; Mechelli, L.; Pepe, M.; et al. PD-L1/PD-1 and CTLA-4 Expression in Equine Penile Squamous Cell Carcinomas. Animals 2021, 11, 2121. [Google Scholar] [CrossRef]
- Vichi, G.; Porcellato, I.; Mechelli, L.; Fantauzzo, G.; Razzuoli, E.; Modesto, P.; Mecocci, S.; Brachelente, C. Case Report Co-occurrence of Papillomas Related to Equus caballus Papillomavirus Type 2 and Cutaneous Habronemiasis. Eq. Vet. Educ. 2022, 34, e193–e197. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. 2010. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 26 April 2010).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Thorvaldsdóttir, H.; Robinson, J.T.; Mesirov, J.P. Integrative Genomics Viewer (IGV): High-performance genomics data visualization and exploration. Brief Bioinform. 2013, 14, 178–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tcherepanov, V.; Ehlers, A.; Upton, C. Genome Annotation Transfer Utility (GATU): Rapid annotation of viral genomes using a closely related reference genome. BMC Genom. 2006, 7, 150. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- LeBlanc, S.J. Interactions of Metabolism, Inflammation, and Reproductive Tract Health in thePostpartum Period in Dairy Cattle. Reprod. Dom. Anim. 2012, 47, 18–30. [Google Scholar] [CrossRef]
- Munn, L. Cancer and Inflammation. Wiley Interdiscip. Rev. Syst. Biol. Med. 2017, 9, e1370. [Google Scholar] [CrossRef] [Green Version]
- Zhu, K.W.; Affolter, V.K.; Gaynor, A.M.; Dela Cruz, F.N.; Pesavento, P.A. Equine Genital Squamous Cell Carcinoma: In Situ Hybridization Identifies a Distinct Subset Containing Equus caballus Papillomavirus 2. Vet. Pathol. 2015, 52, 1067–1072. [Google Scholar] [CrossRef] [PubMed]
- Swan, D.C.; Tucker, R.A.; Tortolero-Luna, G.; Mitchell, M.F.; Wideroff, L.; Unger, E.R.; Nisenbaum, R.A.; Reeves, W.C.; Icenogle, J.P. Human Papillomavirus (HPV) DNA Copy Number Is Dependent on Grade of Cervical Disease and HPV Type. J. Clin. Microbiol. 1999, 37, 1030–1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broström, H.; Fahlbrink, E.; Dubath, M.L.; Lazary, S. Association between Equine Leucocyte Antigens (ELA) and Equine Sarcoid Tumors in the Population of Swedish Halfbreds and Some of Their Families. Vet. Immunol. Immunopathol. 1988, 19, 215–223. [Google Scholar] [CrossRef]
- Staiger, E.A.; Tseng, C.T.; Miller, D.; Cassano, J.M.; Nasir, L.; Garrick, D.; Brooks, S.A.; Antczak, D.F. Host Genetic Influence on Papillomavirus-Induced Tumors in the Horse. Int. J. Cancer 2016, 139, 784–792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Araujo Souza, P.S.; Sichero, L.; Maciag, P.C. HPV Variants and HLA Polymorphisms: The Role of Variability on the Risk of Cervical Cancer. Future Oncol. Lond. Engl. 2009, 5, 359–370. [Google Scholar] [CrossRef] [PubMed]
- Porcellato, I.; Mecocci, S.; Mechelli, L.; Cappelli, K.; Brachelente, C.; Pepe, M.; Orlandi, M.; Gialletti, R.; Passeri, B.; Ferrari, A.; et al. Equine Penile Squamous Cell Carcinomas as a Model for Human Disease: A Preliminary Investigation on Tumor Immune Microenvironment. Cells 2020, 9, 2364. [Google Scholar] [CrossRef]
- Ferris, D.G.; Brown, D.R.; Giuliano, A.R.; Myers, E.; Joura, E.A.; Garland, S.M.; Kjaer, S.K.; Perez, G.; Saah, A.; Luxembourg, A.; et al. Prevalence, Incidence, and Natural History of HPV Infection in Adult Women Ages 24 to 45 Participating in a Vaccine Trial. Papillomavirus Res. Amst. Neth. 2020, 10, 100202. [Google Scholar] [CrossRef]
- Skoczyński, M.; Goździcka-Józefiak, A.; Kwaśniewska, A. Risk Factors of the Vertical Transmission of Human Papilloma Virus in Newborns from Singleton Pregnancy—Preliminary Report. J. Matern.-Fetal Neonatal Med. Off. J. Eur. Assoc. Perinat. Med. Fed. Asia Ocean. Perinat. Soc. Int. Soc. Perinat. Obstet. 2014, 27, 239–242. [Google Scholar] [CrossRef]
- Stocco dos Santos, R.C.; Lindsey, C.J.; Ferraz, O.P.; Pinto, J.R.; Mirandola, R.S.; Benesi, F.J.; Birgel, E.H.; Pereira, C.A.; Beçak, W. Bovine Papillomavirus Transmission and Chromosomal Aberrations: An Experimental Model. J. Gen. Virol. 1998, 79 Pt 9, 2127–2135. [Google Scholar] [CrossRef]
- McBride, A.A. Human Papillomaviruses: Diversity, Infection and Host Interactions. Nat. Rev. Microbiol. 2022, 20, 95–108. [Google Scholar] [CrossRef]
- Stoler, M.H.; Wolinsky, S.M.; Whitbeck, A.; Broker, T.R.; Chow, L.T. Differentiation-linked human papillomavirus types 6 and 11 transcription in genital condylomata revealed by in situ hybridization with message-specific RNA probes. Virology 1989, 172, 331–340. [Google Scholar] [CrossRef]
- Finkler, E.B.; Stoler, M.H.; Girardi, F.; Pfister, H. Cell differentiation-related gene expression of human papillomavirus 33. Med. Microbiol. Immunol. 1990, 179, 185–192. [Google Scholar] [CrossRef]
- Middleton, K.; Peh, W.; Southern, S.; Griffin, H.; Sotlar, K.; Nakahara, T.; El-Sherif, A.; Morris, L.; Seth, R.; Hibma, M.; et al. Organization of Human Papillomavirus Productive Cycle during Neoplastic Progression Provides a Basis for Selection of Diagnostic Markers. Am. Soc. Microbiol. J. Virol. 2003, 77, 10186–10201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinarello, C.A. Immunological and inflammatory functions of the interleukin-1 family. Annu. Rev. Immunol. 2009, 27, 519–550. [Google Scholar] [CrossRef]
- Ainouze, M.; Rochefort, P.; Parroche, P.; Roblot, G.; Tout, I.; Briat, F.; Zannetti, C.; Marotel, M.; Goutagny, N.; Auron, P.; et al. Human papillomavirus type 16 antagonizes IRF6 regulation of IL-1beta. PLoS Pathog. 2018, 14, e1007158. [Google Scholar] [CrossRef] [PubMed]
- Scagnolari, C.; Cannella, F.; Pierangeli, A.; Mellinger Pilgrim, R.; Antonelli, G.; Rowley, D.; Wong, M.; Best, S.; Xing, D.; Roden, R.B.S.; et al. Insights into the Role of Innate Immunity in Cervicovaginal Papillomavirus Infection from Studies Using Gene-Deficient Mice. J. Virol. 2020, 94, e00087-20. [Google Scholar] [CrossRef] [PubMed]
- Song, D.; Li, H.; Li, H.; Dai, J. Effect of human papillomavirus infection on the immune system and its role in the course of cervical cancer. Oncol. Lett. 2015, 10, 600–606. [Google Scholar] [CrossRef] [Green Version]
- van Dam, P.A.; Verhoeven, Y.; Trinh, X.B.; Wouters, A.; Lardon, F.; Prenen, H.; Smits, E.; Baldewijns, M.; Lammens, M. RANK/RANKL Signaling Inhibition May Improve the Effectiveness of Checkpoint Blockade in Cancer Treatment. Crit. Rev. Oncol. Hematol. 2019, 133, 85–91. [Google Scholar] [CrossRef]
- Campo, L.; Zhang, C.; Breuer, E.-K. EMT-Inducing Molecular Factors in Gynecological Cancers. BioMed Res. Int. 2015, 2015, 420891. [Google Scholar] [CrossRef] [Green Version]
- Pickup, M.; Novitskiy, S.; Moses, H.L. The Roles of TGFβ in the Tumour Microenvironment. Nat. Rev. Cancer 2013, 13, 788–799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lam, J.O.; Bream, J.H.; Sugar, E.A.; Coles, C.L.; Weber, K.M.; Burk, R.D.; Wiley, D.J.; Cranston, R.D.; Reddy, S.; Margolick, J.B.; et al. Association of Serum Cytokines with Oral HPV Clearance. Cytokine 2016, 83, 85–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moerman-Herzog, A.; Nakagawa, M. Early Defensive Mechanisms against Human Papillomavirus Infection. Clin. Vaccine Immunol. CVI 2015, 22, 850–857. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequences | Amplicon Length | Accession Number |
|---|---|---|---|
| EcPV2-L1 | F-5′-TTGTCCAGGAGAGGGGTTAG-3′ R-5′-TGCCTTCCTTTTCTTGGTGG-3′ | 80 | NC_012123.1:5673-7172 |
| p-EcPV2-L1 | FAM-CGTCCAGCACCTTCGACCACCA-TAMRA | ||
| EcPV2-E6 | F-5′-CGTTGGCCTTCTTTGCATCT-3′ R-5′-AGGTTCAGGTCTGCTGTGTT-3′ | 80 | NC_012123.1:5-622 |
| p-EcPV2-E6 | FAM- CCGTGTGGCTATGCTGATGACATTTGG-TAMRA | ||
| EcPV2-E7 | F-5′-CTCTGAGCAGCATCACCCTT-3′ R-5′- TCTTCCTCGTCTTCTGTGTCC-3′ | 70 | NC_012123.1:624-959 |
| p-EcPV2-E7 | FAM- AGAGCGCTCCCCCTCAGTCA-TAMRA | ||
| EcPV2-E2 | F-5′-AAAAGGGAGGGTACGTTGTC-3′ R-5′-CCTGGTAGTAGACATGCTGC-3′ | 90 | NC_012123.1:2767-4017 |
| p-EcPV2-E2 | FAM- GCCAAGACAGCCACGACGCCAT-TAMRA | ||
| B2M | F-5′-GGCTACTCTCCCTGACTGG-3′ R-5′- TCAATCTCAGGCGGATGGAA-3′ | 135 | NM_001082502.3 |
| p-B2M | FAM-ACTCACGTCACCCAGCAGAGA-TAMRA |
| Gene | Primer Pairs Sequences | Amplicon Length | Accession Number |
|---|---|---|---|
| RANKL | F-5′-AGCCTGACACTCAACCTTTTG-3′ R-5′-CCAGGAAGACAGACTCACTTTG-3′ | 86 | XM_014732051.2 |
| IL12p35 | F-5′-CTGAGGACCGTCAGCAACAC-3′ R-5′-GTTCGGGGCGAGTTCCAG-3′ | 147 | NM_001082511.2 |
| IL12p40 | F-5′-GATCGTGGTGGATGCTGTTC-3′ R-5′-TCCACCTGCCGAGAATTCTT-3′ | 132 | NM_001082516.1 |
| CXCL8 | F-5′-CTGGCTGTGGCTCTCTTG-3′ R-5′-CAGTTTGGGATTGAAAGGTTTG-3′ | 133 | NM_001083951.2 |
| IL1B | F-5′-TGATGCAGCTGTGCATTCAGT-3′ R-5′-GCACAAAGCTCATGCAGAACA-3′ | 146 | NM_001082526.1 |
| TGFB | F-5′-CGGAATGGCTGTCCTTTGATG-3′ F-5′-CCCACGCGGAGTGTGTTAT-3′ | 127 | NM_001081849.1 |
| Italian Standardbred | Thoroughbred | Shire | Italian Saddle | |
|---|---|---|---|---|
| Positive | 14 | 33 | 3 | 0 |
| Negative | 41 | 35 | 2 | 1 |
| Total | 55 | 68 | 5 | 1 |
| Positive | Negative | Total | |
|---|---|---|---|
| Maiden | 4 | 21 | 25 |
| Pluriparous | 47 | 57 | 104 |
| Total | 51 | 78 | 129 |
| p-value | 0.0111 | ||
| No foal | 21 | 18 | 39 |
| Foal | 26 | 52 | 78 |
| Total | 47 | 70 | 117 |
| p-value | 0.0452 | ||
| Natural service | 34 | 31 | 65 |
| Artificial insemination | 16 | 48 | 64 |
| Total | 50 | 79 | 129 |
| p-value | 0.0037 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cappelli, K.; Ciucis, C.G.D.; Mecocci, S.; Nervo, T.; Crescio, M.I.; Pepe, M.; Gialletti, R.; Pietrucci, D.; Migone, L.F.; Turco, S.; et al. Detection of Equus Caballus Papillomavirus Type-2 in Asymptomatic Italian Horses. Viruses 2022, 14, 1696. https://doi.org/10.3390/v14081696
Cappelli K, Ciucis CGD, Mecocci S, Nervo T, Crescio MI, Pepe M, Gialletti R, Pietrucci D, Migone LF, Turco S, et al. Detection of Equus Caballus Papillomavirus Type-2 in Asymptomatic Italian Horses. Viruses. 2022; 14(8):1696. https://doi.org/10.3390/v14081696
Chicago/Turabian StyleCappelli, Katia, Chiara Grazia De Ciucis, Samanta Mecocci, Tiziana Nervo, Maria Ines Crescio, Marco Pepe, Rodolfo Gialletti, Daniele Pietrucci, Laura Federica Migone, Silvia Turco, and et al. 2022. "Detection of Equus Caballus Papillomavirus Type-2 in Asymptomatic Italian Horses" Viruses 14, no. 8: 1696. https://doi.org/10.3390/v14081696
APA StyleCappelli, K., Ciucis, C. G. D., Mecocci, S., Nervo, T., Crescio, M. I., Pepe, M., Gialletti, R., Pietrucci, D., Migone, L. F., Turco, S., Mechelli, L., Passamonti, F., Drago, C., Donato, G. G., Varello, K., Modesto, P., Chillemi, G., Ghelardi, A., & Razzuoli, E. (2022). Detection of Equus Caballus Papillomavirus Type-2 in Asymptomatic Italian Horses. Viruses, 14(8), 1696. https://doi.org/10.3390/v14081696