
Recombinant Muscovy Duck Parvovirus Led to Ileac Damage in Muscovy Ducklings


Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus and Viral DNA Extraction
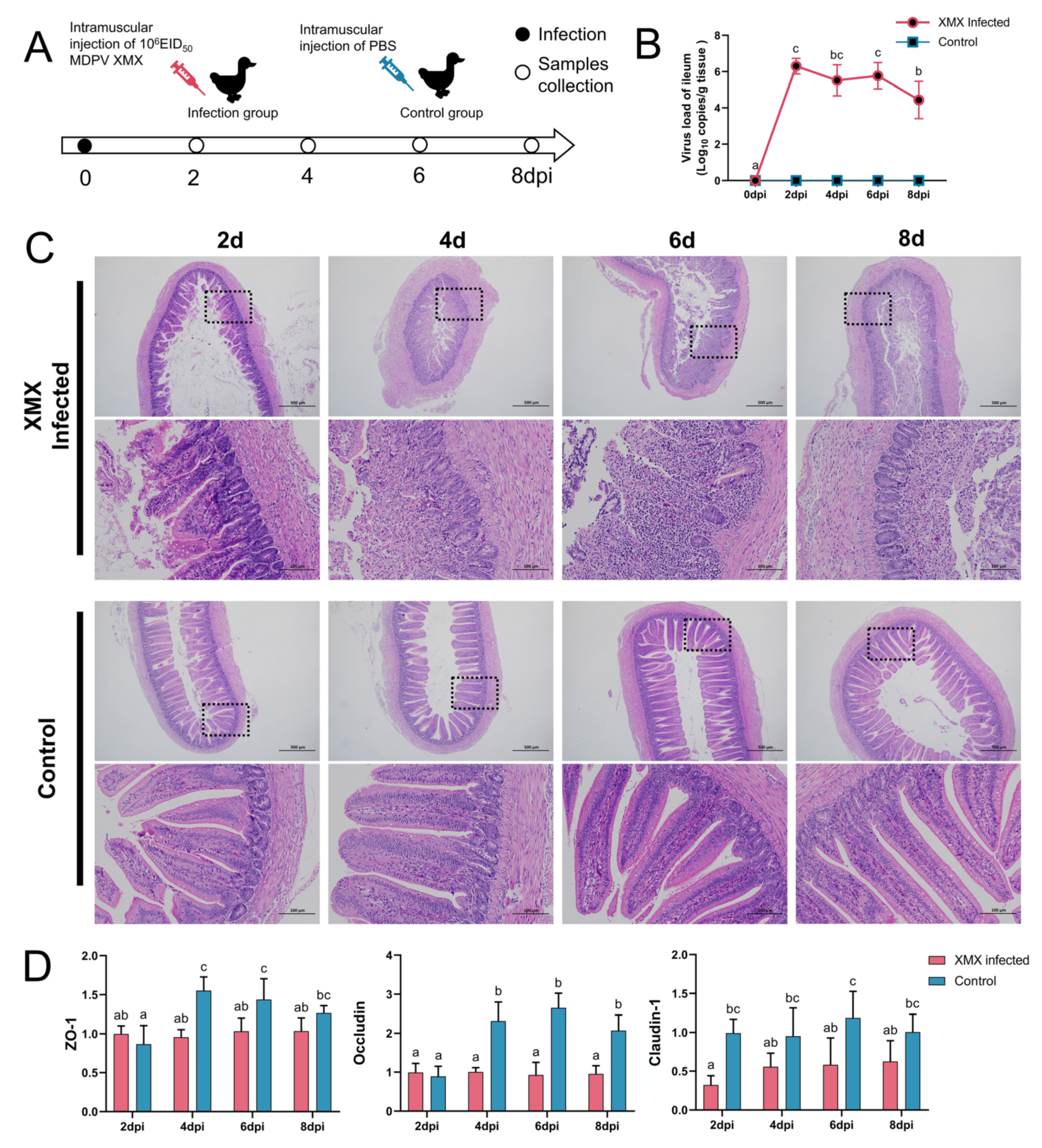
2.2. Animal Experiments
2.3. Sample Collection
2.4. Real-Time Quantitative PCR (qRT-PCR)
2.5. Assay of Intestinal Flora
2.6. Detection of Plasma Antioxidant Capacity
2.7. Genome Sequencing and Analysis
2.8. Statistics
3. Results
3.1. XMX Infection Causes Intestinal Structure Injury
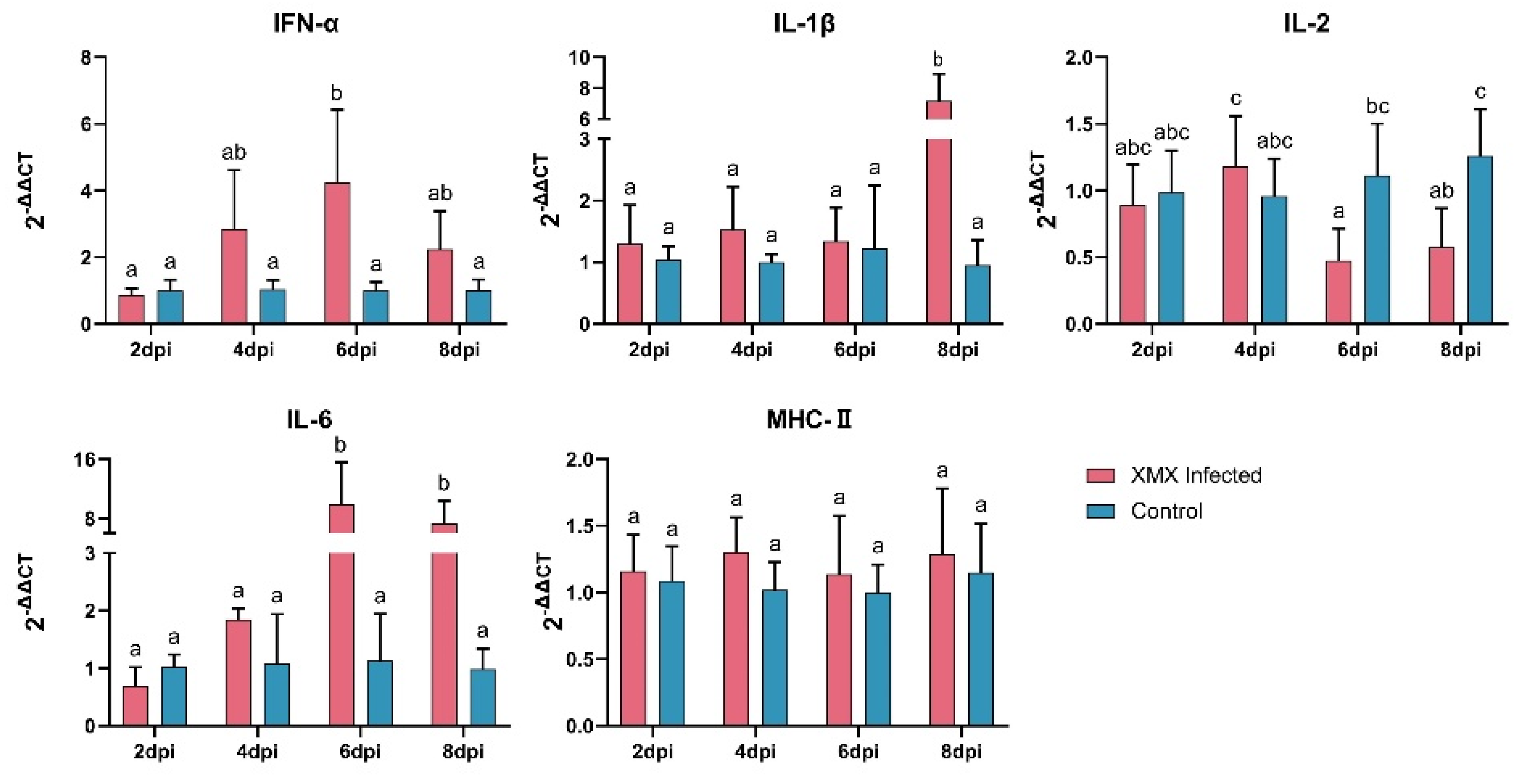
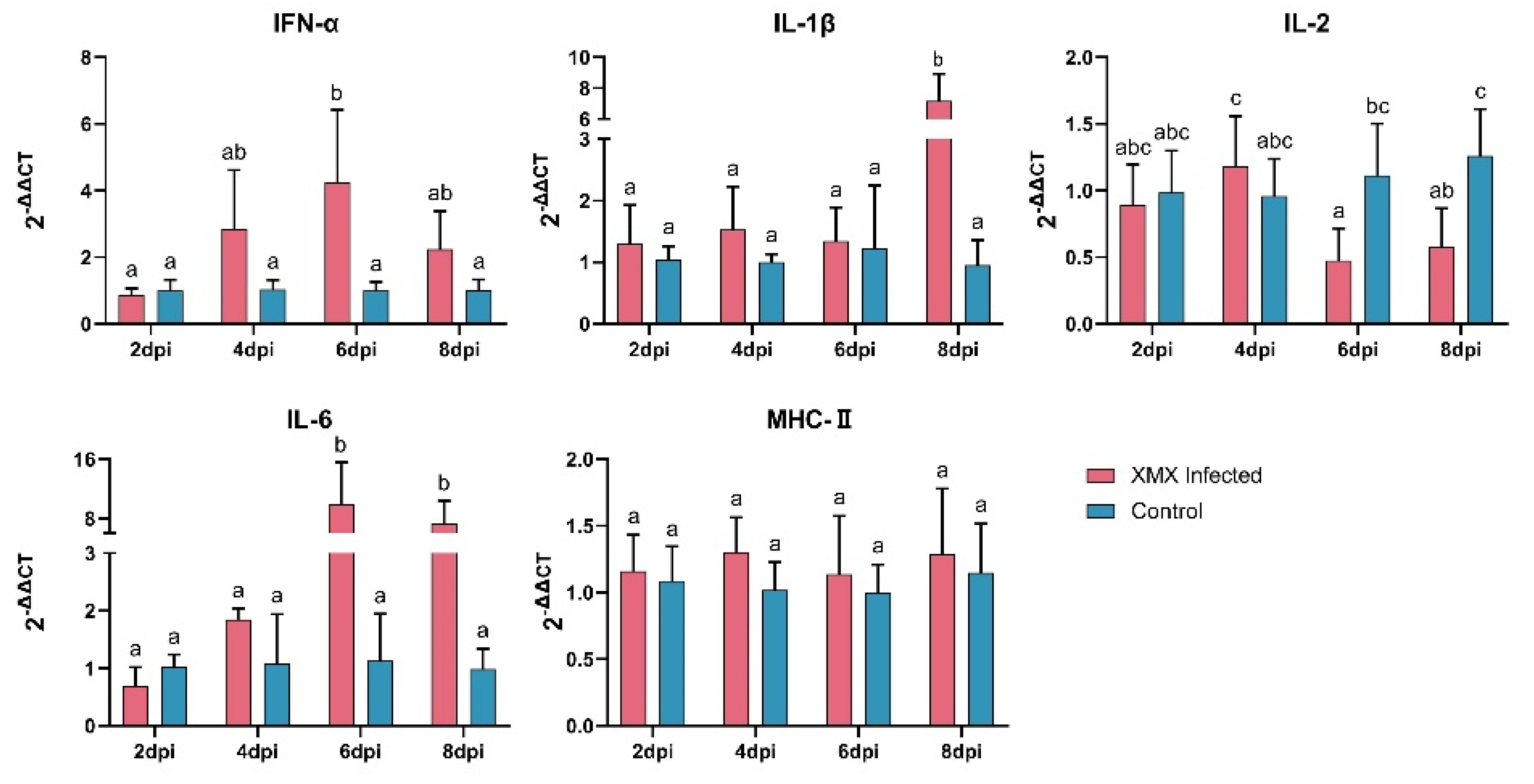
3.2. XMX Infection Affects mRNA Expression of IFN-α, IL-1β, IL-2, and IL-6
3.3. XMX Infection Affects the Diversity of Ileac Microbiota
3.4. XMX Infection Affects the Composition of Ileac Microbiota
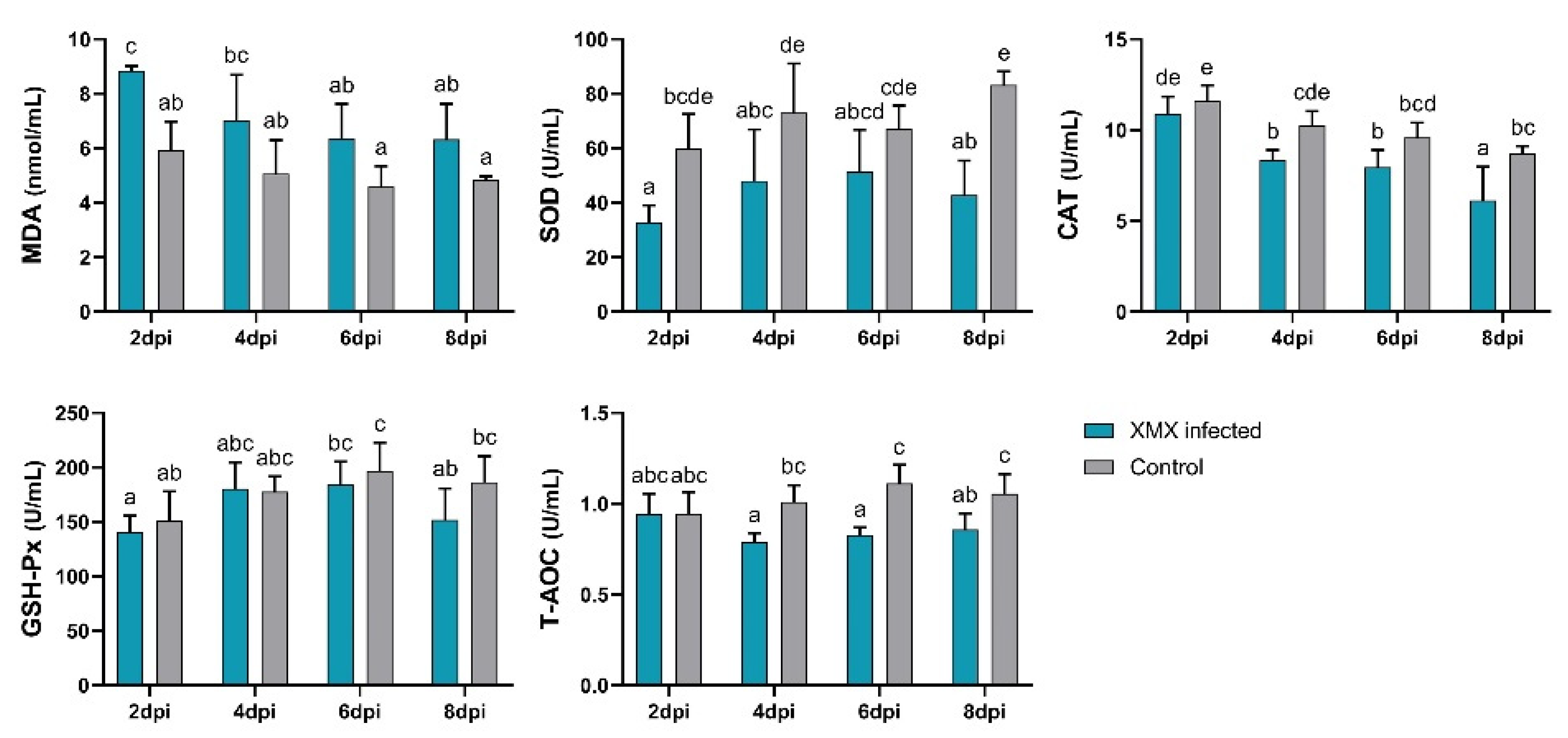
3.5. XMX Infection Reduces Plasma Antioxidant Capacity
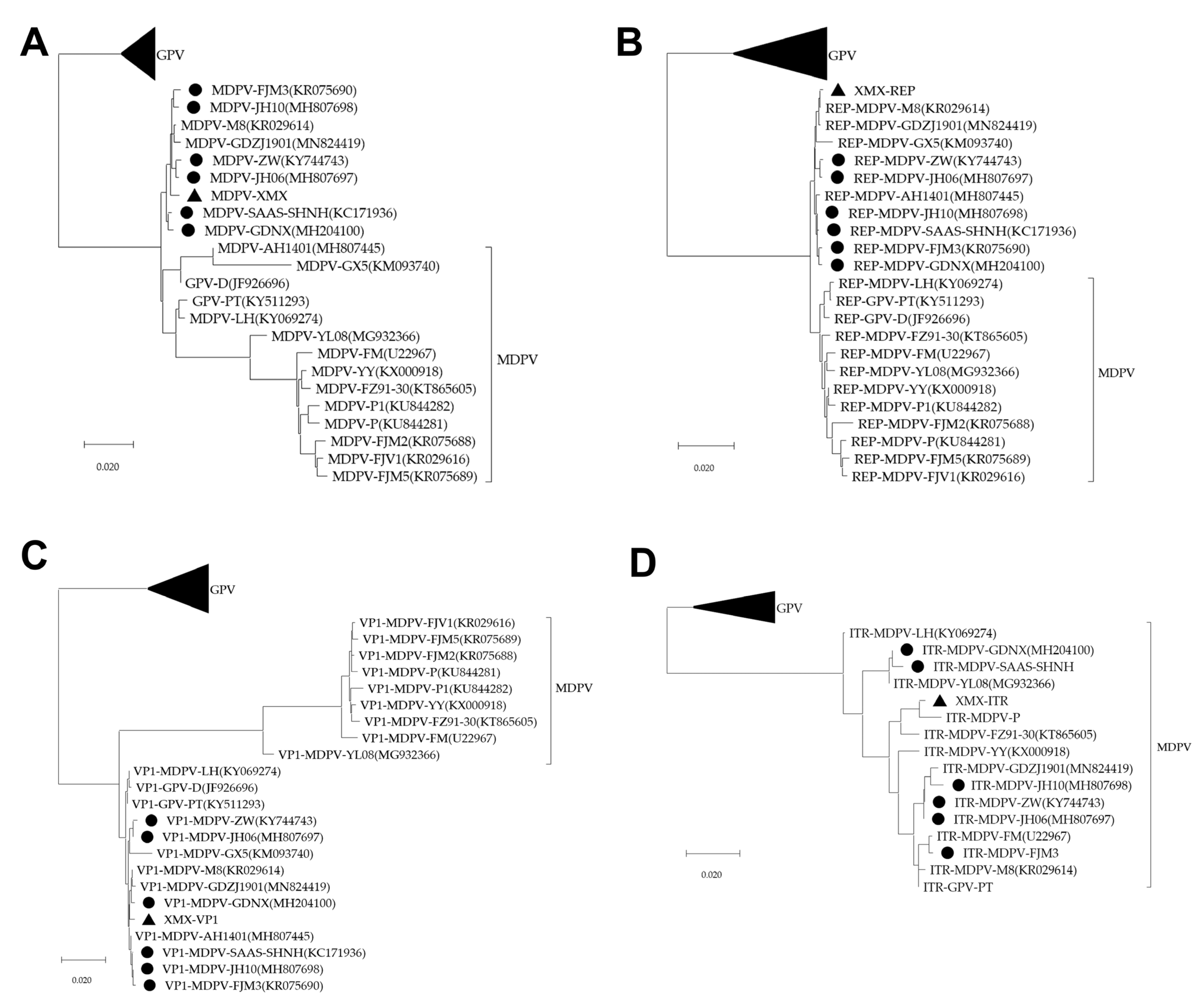
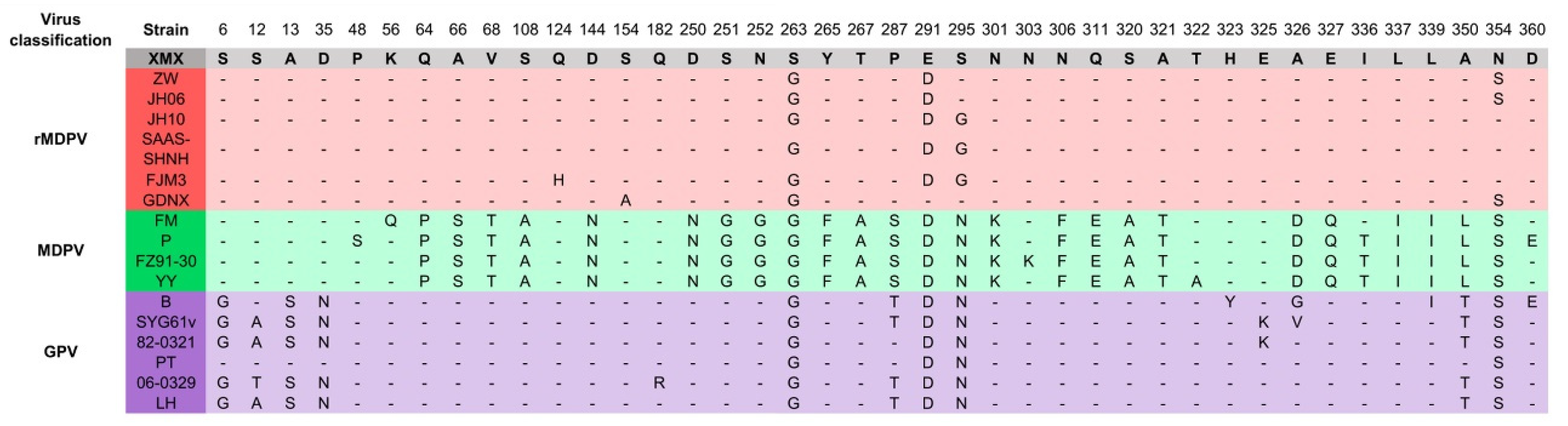
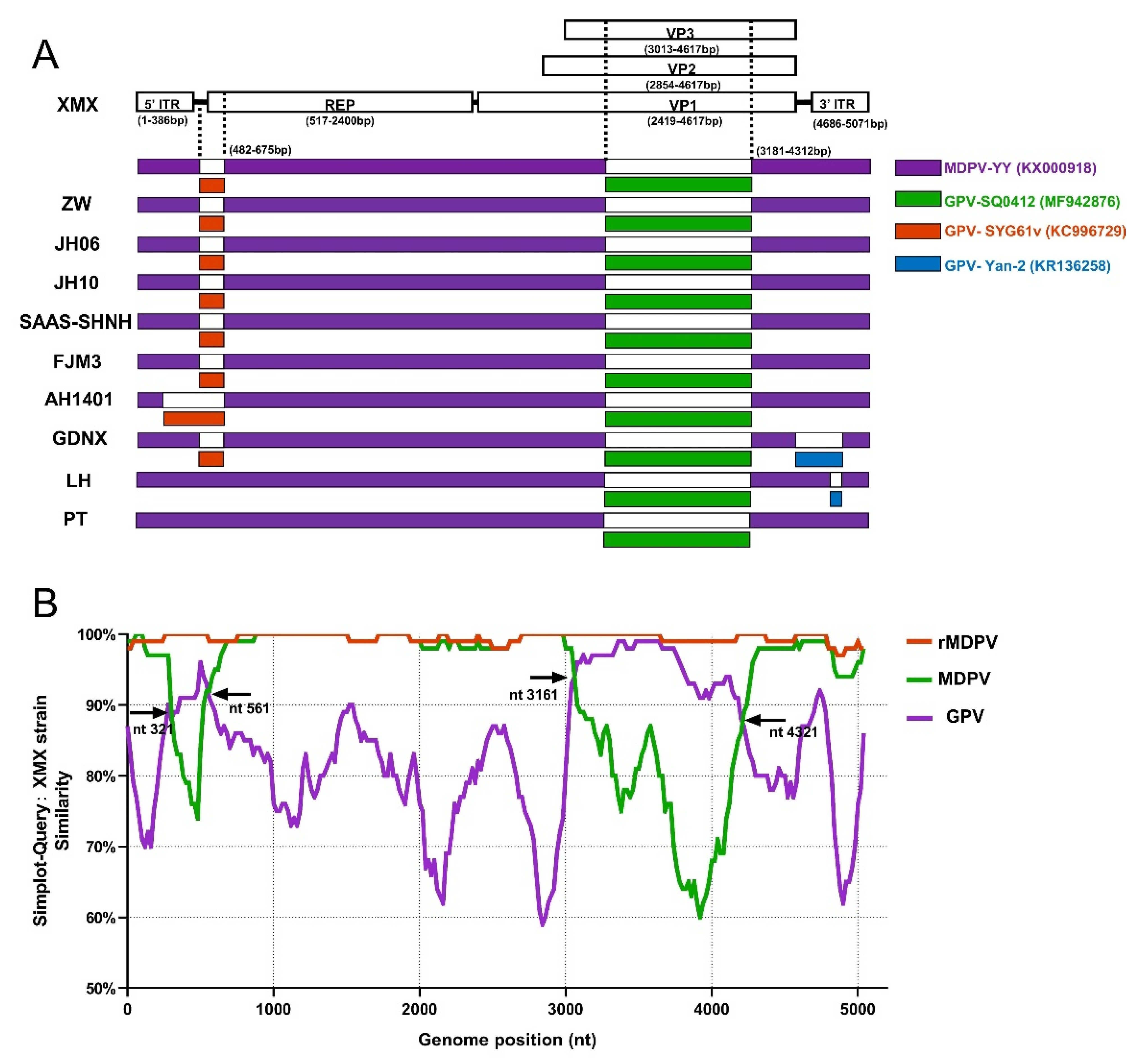
3.6. Genomic Sequence and Analysis
3.7. Recombination Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kailasan, S.; Agbandje-McKenna, M.; Parrish, C.R. Parvovirus Family Conundrum: What Makes a Killer? Annu. Rev. Virol. 2015, 2, 425–450. [Google Scholar] [CrossRef] [PubMed]
- Pénzes, J.J.; de Souza, W.M.; Agbandje-McKenna, M.; Gifford, R.J. An Ancient Lineage of Highly Divergent Parvoviruses Infects both Vertebrate and Invertebrate Hosts. Viruses 2019, 11, 525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, S.T.; Yu, X.L.; Chen, B.D.; Chen, B.L. Diagnosis of a new viral infectious disease in Muscovy ducklings. Chin. J. Prev. Vet. Med. 1991, 2, 27–28. [Google Scholar]
- Woolcock, P.R.; Jestin, V.; Shivaprasad, H.L.; Zwingelstein, F.; Arnauld, C.; McFarland, M.D.; Pedersen, J.C.; Senne, D.A. Evidence of Muscovy duck parvovirus in Muscovy ducklings in California. Vet. Rec. 2000, 146, 68–72. [Google Scholar] [CrossRef]
- Poonia, B.; Dunn, P.A.; Lu, H.; Jarosinski, K.W.; Schat, K.A. Isolation and molecular characterization of a new Muscovy duck parvovirus from Muscovy ducks in the USA. Avian Pathol. 2006, 35, 435–441. [Google Scholar] [CrossRef]
- Palya, V.J. Viral Infections of Waterfowl: Parvovirus Infections of Waterfowl. In Diseases of Poultry; Wiley-Blackwell: Ames, IO, USA, 2013. [Google Scholar]
- Fu, Q.; Huang, Y.; Wan, C.; Fu, G.; Qi, B.; Cheng, L.; Shi, S.; Chen, H.; Liu, R.; Chen, Z. Genomic and pathogenic analysis of a Muscovy duck parvovirus strain causing short beak and dwarfism syndrome without tongue protrusion. Res. Vet. Sci. 2017, 115, 393–400. [Google Scholar] [CrossRef]
- Yang, J.L.; Cheng, A.C.; Wang, M.S.; Pan, K.C.; Li, M.; Guo, Y.F.; Li, C.F.; Zhu, D.K.; Chen, X.Y. Development of a fluorescent quantitative real-time polymerase chain reaction assay for the detection of Goose parvovirus in vivo. Virol. J. 2009, 6, 142. [Google Scholar] [CrossRef] [Green Version]
- Woźniakowski, G.; Samorek-Salamonowicz, E.; Kozdruń, W. Quantitative analysis of waterfowl parvoviruses in geese and Muscovy ducks by real-time polymerase chain reaction: Correlation between age, clinical symptoms and DNA copy number of waterfowl parvoviruses. BMC Vet. Res. 2012, 8, 29. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Tang, Y.; Dou, Y.; Zheng, X.; Diao, Y. Evidence for Vertical Transmission of Novel Duck-Origin Goose Parvovirus-Related Parvovirus. Transbound. Emerg. Dis. 2016, 63, 243–247. [Google Scholar] [CrossRef]
- Wang, J.; Mi, Q.; Wang, Z.; Jia, J.; Li, Y.; Zhu, G. Sole recombinant Muscovy duck parvovirus infection in Muscovy ducklings can form characteristic intestinal embolism. Vet. Microbiol. 2020, 242, 108590. [Google Scholar] [CrossRef]
- Li, L.; Qiu, J.; Pintel, D.J. The choice of translation initiation site of the rep proteins from goose parvovirus P9-generated mRNA is governed by splicing and the nature of the excised intron. J. Virol. 2009, 83, 10264–10268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Li, Y.; Liu, M.; Zhang, D.; Guo, D.; Liu, C.; Zhi, H.; Wang, X.; Li, G.; Li, N.; et al. Development and evaluation of a VP3-ELISA for the detection of goose and Muscovy duck parvovirus antibodies. J. Virol. Methods 2010, 163, 405–409. [Google Scholar] [CrossRef] [PubMed]
- Le Gall-Reculé, G.; Jestin, V. Biochemical and genomic characterization of muscovy duck parvovirus. Arch. Virol. 1994, 139, 121–131. [Google Scholar] [CrossRef]
- Zádori, Z.; Stefancsik, R.; Rauch, T.; Kisary, J. Analysis of the complete nucleotide sequences of goose and muscovy duck parvoviruses indicates common ancestral origin with adeno-associated virus 2. Virology 1995, 212, 562–573. [Google Scholar] [CrossRef] [Green Version]
- Le Gall-Reculé, G.; Jestin, V.; Chagnaud, P.; Blanchard, P.; Jestin, A. Expression of muscovy duck parvovirus capsid proteins (VP2 and VP3) in a baculovirus expression system and demonstration of immunity induced by the recombinant proteins. J. Gen. Virol. 1996, 77, 2159–2163. [Google Scholar] [CrossRef]
- Wang, J.; Huang, Y.; Zhou, M.; Hardwidge, P.R.; Zhu, G. Construction and sequencing of an infectious clone of the goose embryo-adapted Muscovy duck parvovirus vaccine strain FZ91-30. Virol. J. 2016, 13, 104. [Google Scholar] [CrossRef] [Green Version]
- Cotmore, S.F.; Agbandje-McKenna, M.; Chiorini, J.A.; Mukha, D.V.; Pintel, D.J.; Qiu, J.; Soderlund-Venermo, M.; Tattersall, P.; Tijssen, P.; Gatherer, D.; et al. The family Parvoviridae. Arch. Virol. 2014, 159, 1239–1247. [Google Scholar] [CrossRef] [PubMed]
- Kapgate, S.S.; Kumanan, K.; Vijayarani, K.; Barbuddhe, S.B. Avian parvovirus: Classification, phylogeny, pathogenesis and diagnosis. Avian Pathol. 2018, 47, 536–545. [Google Scholar] [CrossRef]
- Fan, W.; Sun, Z.; Shen, T.; Xu, D.; Huang, K.; Zhou, J.; Song, S.; Yan, L. Analysis of Evolutionary Processes of Species Jump in Waterfowl Parvovirus. Front. Microbiol. 2017, 8, 421. [Google Scholar] [CrossRef] [Green Version]
- Glávits, R.; Zolnai, A.; Szabó, E.; Ivanics, E.; Zarka, P.; Mató, T.; Palya, V. Comparative pathological studies on domestic geese (Anser anser domestica) and Muscovy ducks (Cairina moschata) experimentally infected with parvovirus strains of goose and Muscovy duck origin. Acta Vet. Hung. 2005, 53, 73–89. [Google Scholar] [CrossRef]
- Dong, J.; Bingga, G.; Sun, M.; Li, L.; Liu, Z.; Zhang, C.; Guo, P.; Huang, Y.; Zhang, J. Application of high-resolution melting curve analysis for identification of Muscovy duck parvovirus and goose parvovirus. J. Virol. Methods 2019, 266, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Wan, C.H.; Chen, H.M.; Qiu-Ling, F.U.; Guang-Hua, F.U.; Cheng, L.F.; Shi, S.H.; Huang, Y.; Kai-Hui, H.U. Genomic Characterization of Goose Parvovirus and Muscovy Duck Parvovirus Co-infection in Fujian, China. Kafkas Univ. Vet. Fak. Derg. 2015, 21, 923–928. [Google Scholar]
- Wang, J.; Ling, J.; Wang, Z.; Huang, Y.; Zhu, J.; Zhu, G. Molecular characterization of a novel Muscovy duck parvovirus isolate: Evidence of recombination between classical MDPV and goose parvovirus strains. BMC Vet. Res. 2017, 13, 327. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Zhou, Z.; Huang, Y.; Yu, R.; Dong, S.; Li, Z.; Zhang, Y. Identification of a recombinant Muscovy Duck parvovirus (MDPV) in Shanghai, China. Vet. Microbiol. 2014, 174, 560–564. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Huang, J.; Yan, Z.; Yin, L.; Li, Q.; Zhou, Q.; Chen, F. Isolation and characterization of a recombinant Muscovy duck parvovirus circulating in Muscovy ducks in South China. Arch. Virol. 2020, 165, 2931–2936. [Google Scholar] [CrossRef]
- Liu, R.; Chen, C.; Huang, Y.; Cheng, L.; Lu, R.; Fu, G.; Shi, S.; Chen, H.; Wan, C.; Fu, Q.; et al. Microbiological identification and analysis of waterfowl livers collected from backyard farms in southern China. J. Vet. Med. Sci. 2018, 80, 667–671. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Wang, Z.; Jia, J.; Ling, J.; Mi, Q.; Zhu, G. Retrospective investigation and molecular characteristics of the recombinant Muscovy duck parvovirus circulating in Muscovy duck flocks in China. Avian Pathol. 2019, 48, 343–351. [Google Scholar] [CrossRef]
- Wang, S.; Cheng, X.X.; Chen, S.Y.; Zhu, X.L.; Chen, S.L.; Lin, F.Q.; Li, Z.L. Genetic characterization of a potentially novel goose parvovirus circulating in Muscovy duck flocks in Fujian Province, China. J. Vet. Med. Sci. 2013, 75, 1127–1130. [Google Scholar] [CrossRef] [Green Version]
- Day, J.M.; Zsak, L. Recent progress in the characterization of avian enteric viruses. Avian. Dis. 2013, 57, 573–580. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Diao, Y.; Wang, D.; Chen, H.; Tang, Y.; Diao, Y. Duck viral infection escalated the incidence of avian pathogenic Escherichia coli in China. Transbound. Emerg. Dis. 2019, 66, 929–938. [Google Scholar] [CrossRef]
- Luo, Q.; Lei, X.; Xu, J.; Jahangir, A.; He, J.; Huang, C.; Liu, W.; Cheng, A.; Tang, L.; Geng, Y.; et al. An altered gut microbiota in duck-origin parvovirus infection on cherry valley ducklings is associated with mucosal barrier dysfunction. Poult. Sci. 2021, 100, 101021. [Google Scholar] [CrossRef] [PubMed]
- Pereira, G.Q.; Gomes, L.A.; Santos, I.S.; Alfieri, A.F.; Weese, J.S.; Costa, M.C. Fecal microbiota transplantation in puppies with canine parvovirus infection. J. Vet. Intern. Med. 2018, 32, 707–711. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Wang, X.L. Species diversity of fecal microbial flora in Canis lupus familiaris infected with canine parvovirus. Vet. Microbiol. 2019, 237, 108390. [Google Scholar] [CrossRef] [PubMed]
- Hooper, L.V.; Gordon, J.I. Commensal host-bacterial relationships in the gut. Science 2001, 292, 1115–1118. [Google Scholar] [CrossRef]
- Broom, L.J.; Kogut, M.H. The role of the gut microbiome in shaping the immune system of chickens. Vet. Immunol. Immunopathol. 2018, 204, 44–51. [Google Scholar] [CrossRef]
- Garrett, W.S.; Lord, G.M.; Punit, S.; Lugo-Villarino, G.; Mazmanian, S.K.; Ito, S.; Glickman, J.N.; Glimcher, L.H. Communicable ulcerative colitis induced by T-bet deficiency in the innate immune system. Cell 2007, 131, 33–45. [Google Scholar] [CrossRef] [Green Version]
- Barreau, F.; Hugot, J.P. Intestinal barrier dysfunction triggered by invasive bacteria. Curr. Opin. Microbiol. 2014, 17, 91–98. [Google Scholar] [CrossRef]
- Chen, Q.; Xu, X.; Yu, Z.; Sui, C.; Zuo, K.; Zhi, G.; Ji, J.; Yao, L.; Kan, Y.; Bi, Y.; et al. Characterization and genomic analysis of emerging astroviruses causing fatal gout in goslings. Transbound. Emerg. Dis. 2020, 67, 865–876. [Google Scholar] [CrossRef]
- Lin, W.; Chen, T.; Liao, L.; Wang, Z.; Xiao, J.; Lu, J.; Song, C.; Qin, J.; Chen, F.; Chang, Y.F.; et al. A parrot-type Chlamydia psittaci strain is in association with egg production drop in laying ducks. Transbound. Emerg. Dis. 2019, 66, 2002–2010. [Google Scholar] [CrossRef]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Haas, B.J.; Gevers, D.; Earl, A.M.; Feldgarden, M.; Ward, D.V.; Giannoukos, G.; Ciulla, D.; Tabbaa, D.; Highlander, S.K.; Sodergren, E.; et al. Chimeric 16S rRNA sequence formation and detection in Sanger and 454-pyrosequenced PCR amplicons. Genome Res. 2011, 21, 494–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Shao, D.; Zhou, J.; Gu, J.; Qin, J.; Chen, W.; Wei, W. Signatures within esophageal microbiota with progression of esophageal squamous cell carcinoma. Chin. J. Cancer Res. 2020, 32, 755–767. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Martin, D.P.; Murrell, B.; Golden, M.; Khoosal, A.; Muhire, B. RDP4: Detection and analysis of recombination patterns in virus genomes. Virus Evol. 2015, 1, vev003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lole, K.S.; Bollinger, R.C.; Paranjape, R.S.; Gadkari, D.; Kulkarni, S.S.; Novak, N.G.; Ingersoll, R.; Sheppard, H.W.; Ray, S.C. Full-length human immunodeficiency virus type 1 genomes from subtype C-infected seroconverters in India, with evidence of intersubtype recombination. J. Virol. 1999, 73, 152–160. [Google Scholar] [CrossRef] [Green Version]
- Wan, C.H.; Qiu, F.U.; Chen, C.T.; Guang, F.U.; Chen, H.M.; Cheng, L.F.; Shi, S.H.; Huang, Y. Molecular characterization of the genome for recombinant Muscovy duck parvovirus strain FJM3. Chin. J. Vet. Sci. 2016, 36, 1836–1841. [Google Scholar]
- Zadori, Z.; Erdei, J.; Nagy, J.; Kisary, J. Characteristics of the genome of goose parvovirus. Avian Pathol. 1994, 23, 359–364. [Google Scholar] [CrossRef]
- Wang, J.; Huang, Y.; Zhou, M.; Zhu, G. Analysis of the genome sequence of the pathogenic Muscovy duck parvovirus strain YY reveals a 14-nucleotide-pair deletion in the inverted terminal repeats. Arch. Virol. 2016, 161, 2589–2594. [Google Scholar] [CrossRef]
- Kisary, J. Some growth characteristics of goose parvovirus strain “B”. Acta. Vet. Acad. Sci. Hung 1974, 24, 329–333. [Google Scholar]
- Wang, J.; Duan, J.; Meng, X.; Gong, J.; Jiang, Z.; Zhu, G. Cloning of the genome of a goose parvovirus vaccine strain SYG61v and rescue of infectious virions from recombinant plasmid in embryonated goose eggs. J. Virol. Methods 2014, 200, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Shien, J.H.; Wang, Y.S.; Chen, C.H.; Shieh, H.K.; Hu, C.C.; Chang, P.C. Identification of sequence changes in live attenuated goose parvovirus vaccine strains developed in Asia and Europe. Avian Pathol. 2008, 37, 499–505. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.Y.; Lin, H.Q.; Cheng, X.X.; Ying, L.Y.; Cheng, Y.Q.; Lin, T.L. Study on Two-combinative Attenuated Vaccine of Goose and Muscovy duckling Parvovirus. Chin. J. Vet. Sci. 2003, 23, 226–228. [Google Scholar]
- Wang, J.; Duan, J.; Zhu, L.; Jiang, Z.; Zhu, G. Sequencing and generation of an infectious clone of the pathogenic goose parvovirus strain LH. Arch. Virol. 2015, 160, 711–718. [Google Scholar] [CrossRef]
- Moon, H.W. Comparative histopathology of intestinal infections. Adv. Exp. Med. Biol. 1997, 412, 1–19. [Google Scholar] [CrossRef]
- Stuetzer, B.; Hartmann, K. Feline parvovirus infection and associated diseases. Vet. J. 2014, 201, 150–155. [Google Scholar] [CrossRef]
- Domanska-Blicharz, K.; Jacukowicz, A.; Lisowska, A.; Minta, Z. Genetic characterization of parvoviruses circulating in turkey and chicken flocks in Poland. Arch. Virol. 2012, 157, 2425–2430. [Google Scholar] [CrossRef] [Green Version]
- LF, N.N.; Santander-Parra, S.H.; De la Torre, D.I.; Sá, L.R.M.; Buim, M.R.; Astolfi-Ferreira, C.S.; Piantino Ferreira, A.J. Molecular Characterization and Pathogenicity of Chicken Parvovirus (ChPV) in Specific Pathogen-Free Chicks Infected Experimentally. Pathogens 2020, 9, 606. [Google Scholar] [CrossRef]
- Finkler, F.; Lima, D.A.; Cerva, C.; Moraes, L.B.; Cibulski, S.P.; Teixeira, T.F.; Santos, H.F.; Almeida, L.L.; Roehe, P.M.; Franco, A.C. Chicken parvovirus and its associations with malabsorption syndrome. Res. Vet. Sci. 2016, 107, 178–181. [Google Scholar] [CrossRef]
- Rodgers, L.S.; Beam, M.T.; Anderson, J.M.; Fanning, A.S. Epithelial barrier assembly requires coordinated activity of multiple domains of the tight junction protein ZO-1. J. Cell Sci. 2013, 126, 1565–1575. [Google Scholar] [CrossRef] [Green Version]
- Günzel, D.; Fromm, M. Claudins and other tight junction proteins. Compr. Physiol. 2012, 2, 1819–1852. [Google Scholar] [CrossRef] [PubMed]
- Inai, T.; Kobayashi, J.; Shibata, Y. Claudin-1 contributes to the epithelial barrier function in MDCK cells. Eur. J. Cell Biol. 1999, 78, 849–855. [Google Scholar] [CrossRef]
- Zhao, J.; Wan, S.; Sun, N.; Sun, P.; Sun, Y.; Khan, A.; Guo, J.; Zheng, X.; Fan, K.; Yin, W.; et al. Damage to intestinal barrier integrity in piglets caused by porcine reproductive and respiratory syndrome virus infection. Vet. Res. 2021, 52, 93. [Google Scholar] [CrossRef] [PubMed]
- Giron, L.B.; Dweep, H.; Yin, X.; Wang, H.; Damra, M.; Goldman, A.R.; Gorman, N.; Palmer, C.S.; Tang, H.Y.; Shaikh, M.W.; et al. Plasma Markers of Disrupted Gut Permeability in Severe COVID-19 Patients. Front. Immunol. 2021, 12, 686240. [Google Scholar] [CrossRef] [PubMed]
- Zong, Q.F.; Huang, Y.J.; Wu, L.S.; Wu, Z.C.; Wu, S.L.; Bao, W.B. Effects of porcine epidemic diarrhea virus infection on tight junction protein gene expression and morphology of the intestinal mucosa in pigs. Pol. J. Vet. Sci. 2019, 22, 345–353. [Google Scholar] [CrossRef]
- Chen, X.; Liu, R.; Liu, X.; Xu, C.; Wang, X. Protective Role of Coxsackie-Adenovirus Receptor in the Pathogenesis of Inflammatory Bowel Diseases. BioMed Res. Int. 2018, 2018, 7207268. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Liu, X.; Chen, F.; Zuo, K.; Wu, C.; Yan, Y.; Chen, W.; Lin, W.; Xie, Q. Avian Influenza Virus Subtype H9N2 Affects Intestinal Microbiota, Barrier Structure Injury, and Inflammatory Intestinal Disease in the Chicken Ileum. Viruses 2018, 10, 270. [Google Scholar] [CrossRef] [Green Version]
- Witkowski, P.T.; Perley, C.C.; Brocato, R.L.; Hooper, J.W.; Jürgensen, C.; Schulzke, J.D.; Krüger, D.H.; Bücker, R. Gastrointestinal Tract As Entry Route for Hantavirus Infection. Front. Microbiol. 2017, 8, 1721. [Google Scholar] [CrossRef] [Green Version]
- Feldman, G.J.; Mullin, J.M.; Ryan, M.P. Occludin: Structure, function and regulation. Adv. Drug Deliv. Rev. 2005, 57, 883–917. [Google Scholar] [CrossRef]
- Reinecker, H.C.; Steffen, M.; Doehn, C.; Petersen, J.; Pflüger, I.; Voss, A.; Raedler, A. Proinflammatory cytokines in intestinal mucosa. Immunol. Res. 1991, 10, 247–248. [Google Scholar] [CrossRef]
- Monteleone, G.; Pender, S.L.; Alstead, E.; Hauer, A.C.; Lionetti, P.; McKenzie, C.; MacDonald, T.T. Role of interferon alpha in promoting T helper cell type 1 responses in the small intestine in coeliac disease. Gut 2001, 48, 425–429. [Google Scholar] [CrossRef] [Green Version]
- Saxena, K.; Simon, L.M.; Zeng, X.L.; Blutt, S.E.; Crawford, S.E.; Sastri, N.P.; Karandikar, U.C.; Ajami, N.J.; Zachos, N.C.; Kovbasnjuk, O.; et al. A paradox of transcriptional and functional innate interferon responses of human intestinal enteroids to enteric virus infection. Proc. Natl. Acad. Sci. USA 2017, 114, E570–E579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, T.; Yoshinaga, N.; Tanabe, S. Interleukin-6 (IL-6) regulates claudin-2 expression and tight junction permeability in intestinal epithelium. J. Biol. Chem. 2011, 286, 31263–31271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bulek, K.; Zhao, J.; Liao, Y.; Rana, N.; Corridoni, D.; Antanaviciute, A.; Chen, X.; Wang, H.; Qian, W.; Miller-Little, W.A.; et al. Epithelial-derived gasdermin D mediates nonlytic IL-1β release during experimental colitis. J. Clin. Investig. 2020, 130, 4218–4234. [Google Scholar] [CrossRef] [PubMed]
- Noval Rivas, M.; Wakita, D.; Franklin, M.K.; Carvalho, T.T.; Abolhesn, A.; Gomez, A.C.; Fishbein, M.C.; Chen, S.; Lehman, T.J.; Sato, K.; et al. Intestinal Permeability and IgA Provoke Immune Vasculitis Linked to Cardiovascular Inflammation. Immunity 2019, 51, 508–521. [Google Scholar] [CrossRef]
- Jeffery, V.; Goldson, A.J.; Dainty, J.R.; Chieppa, M.; Sobolewski, A. IL-6 Signaling Regulates Small Intestinal Crypt Homeostasis. J. Immunol. 2017, 199, 304–311. [Google Scholar] [CrossRef]
- Luo, Q.; Xu, J.; Huang, C.; Lei, X.; Cheng, D.; Liu, W.; Cheng, A.; Tang, L.; Fang, J.; Ou, Y.; et al. Impacts of Duck-Origin Parvovirus Infection on Cherry Valley Ducklings From the Perspective of Gut Microbiota. Front. Microbiol. 2019, 10, 624. [Google Scholar] [CrossRef]
- Zhou, Y.; Jin, X.H.; Jing, Y.X.; Song, Y.; He, X.X.; Zheng, L.L.; Wang, Y.B.; Wei, Z.Y.; Zhang, G.P. Porcine parvovirus infection activates inflammatory cytokine production through Toll-like receptor 9 and NF-κB signaling pathways in porcine kidney cells. Vet. Microbiol. 2017, 207, 56–62. [Google Scholar] [CrossRef]
- Nigro, G.; Bastianon, V.; Colloridi, V.; Ventriglia, F.; Gallo, P.; D’Amati, G.; Koch, W.C.; Adler, S.P. Human parvovirus B19 infection in infancy associated with acute and chronic lymphocytic myocarditis and high cytokine levels: Report of 3 cases and review. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2000, 31, 65–69. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Chu, C.; Teng, F.; Bessman, N.J.; Goc, J.; Santosa, E.K.; Putzel, G.G.; Kabata, H.; Kelsen, J.R.; Baldassano, R.N.; et al. Innate lymphoid cells support regulatory T cells in the intestine through interleukin-2. Nature 2019, 568, 405–409. [Google Scholar] [CrossRef]
- Adams, S.C.; Xing, Z.; Li, J.; Cardona, C.J. Immune-related gene expression in response to H11N9 low pathogenic avian influenza virus infection in chicken and Pekin duck peripheral blood mononuclear cells. Mol. Immunol. 2009, 46, 1744–1749. [Google Scholar] [CrossRef] [PubMed]
- Liang, Q.L.; Luo, J.; Zhou, K.; Dong, J.X.; He, H.X. Immune-related gene expression in response to H5N1 avian influenza virus infection in chicken and duck embryonic fibroblasts. Mol. Immunol. 2011, 48, 924–930. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Cao, C.; Qu, Z.; Zhang, W.; Liu, Y.; Qi, H.; Hao, C.; Zhang, W.; Gao, M.; Wang, J.; et al. Pathogenicity of duck hepatitis A virus type 3 and innate immune responses of the ducklings to virulent DHAV-3. Mol. Immunol. 2018, 95, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Su, S.; Chen, X.; Zhao, L.; Chen, H. Biological characteristic and cytokines response of passages duck plague virus in ducks. Virus Res. 2021, 295, 198320. [Google Scholar] [CrossRef]
- Shin, Y.; Yoon, C.H.; Lim, H.; Park, J.; Roh, T.Y.; Kang, C.; Choi, B.S. Impaired IL-2 expression in latent HIV-1 infection. Biochem. Biophys. Res. Commun. 2015, 463, 1237–1242. [Google Scholar] [CrossRef]
- Fan, J.; Bass, H.Z.; Fahey, J.L. Elevated IFN-gamma and decreased IL-2 gene expression are associated with HIV infection. J. Immunol. 1993, 151, 5031–5040. [Google Scholar]
- Méndez-Lagares, G.; Jaramillo-Ruiz, D.; Pion, M.; Leal, M.; Muñoz-Fernández, M.A.; Pacheco, Y.M.; Correa-Rocha, R. HIV infection deregulates the balance between regulatory T cells and IL-2-producing CD4 T cells by decreasing the expression of the IL-2 receptor in Treg. J. Acquir. Immune Defic. Syndr. 2014, 65, 278–282. [Google Scholar] [CrossRef]
- Biton, M.; Haber, A.L.; Rogel, N.; Burgin, G.; Beyaz, S.; Schnell, A.; Ashenberg, O.; Su, C.W.; Smillie, C.; Shekhar, K.; et al. T Helper Cell Cytokines Modulate Intestinal Stem Cell Renewal and Differentiation. Cell 2018, 175, 1307–1320. [Google Scholar] [CrossRef] [Green Version]
- Zijlstra, R.T.; McCracken, B.A.; Odle, J.; Donovan, S.M.; Gelberg, H.B.; Petschow, B.W.; Zuckermann, F.A.; Gaskins, H.R. Malnutrition modifies pig small intestinal inflammatory responses to rotavirus. J. Nutr. 1999, 129, 838–843. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Zheng, M.; Huang, M.; Xiao, S.; Lin, F.; Chen, S.; Chen, S. Muscovy Duck Reovirus Infection Disrupts the Composition of Intestinal Microbiota in Muscovy Ducklings. Curr. Microbiol. 2020, 77, 769–778. [Google Scholar] [CrossRef]
- Barko, P.C.; McMichael, M.A.; Swanson, K.S.; Williams, D.A. The Gastrointestinal Microbiome: A Review. J. Vet. Intern. Med. 2018, 32, 9–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pammi, M.; Cope, J.; Tarr, P.I.; Warner, B.B.; Morrow, A.L.; Mai, V.; Gregory, K.E.; Kroll, J.S.; McMurtry, V.; Ferris, M.J.; et al. Intestinal dysbiosis in preterm infants preceding necrotizing enterocolitis: A systematic review and meta-analysis. Microbiome 2017, 5, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arias, C.A.; Murray, B.E. The rise of the Enterococcus: Beyond vancomycin resistance. Nat. Rev. Microbiol. 2012, 10, 266–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castañeda, C.D.; Dittoe, D.K.; Wamsley, K.G.S.; McDaniel, C.D.; Blanch, A.; Sandvang, D.; Kiess, A.S. In ovo inoculation of an Enterococcus faecium-based product to enhance broiler hatchability, live performance, and intestinal morphology. Poult. Sci. 2020, 99, 6163–6172. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Zheng, A.; Meng, K.; Chang, W.; Bai, Y.; Li, K.; Cai, H.; Liu, G.; Yao, B. Proteome changes in the intestinal mucosa of broiler (Gallus gallus) activated by probiotic Enterococcus faecium. J. Proteom. 2013, 91, 226–241. [Google Scholar] [CrossRef]
- Wang, S.; Ng, L.H.; Chow, W.L.; Lee, Y.K. Infant intestinal Enterococcus faecalis down-regulates inflammatory responses in human intestinal cell lines. World J. Gastroenterol. 2008, 14, 1067–1076. [Google Scholar] [CrossRef]
- Wu, M.; Yi, D.; Zhang, Q.; Wu, T.; Yu, K.; Peng, M.; Wang, L.; Zhao, D.; Hou, Y.; Wu, G. Puerarin enhances intestinal function in piglets infected with porcine epidemic diarrhea virus. Sci. Rep. 2021, 11, 6552. [Google Scholar] [CrossRef]
- Medina Fernández, S.; Cretenet, M.; Bernardeau, M. In vitro inhibition of avian pathogenic Enterococcus cecorum isolates by probiotic Bacillus strains. Poult. Sci. 2019, 98, 2338–2346. [Google Scholar] [CrossRef]
- Jung, A.; Rautenschlein, S. Development of an in-house ELISA for detection of antibodies against Enterococcus cecorum in Pekin ducks. Avian Pathol. 2020, 49, 355–360. [Google Scholar] [CrossRef]
- Ryan, M.P.; Pembroke, J.T. The Genus Ochrobactrum as Major Opportunistic Pathogens. Microorganisms 2020, 8, 1797. [Google Scholar] [CrossRef]
- Walujkar, S.A.; Kumbhare, S.V.; Marathe, N.P.; Patangia, D.V.; Lawate, P.S.; Bharadwaj, R.S.; Shouche, Y.S. Molecular profiling of mucosal tissue associated microbiota in patients manifesting acute exacerbations and remission stage of ulcerative colitis. World J. Microbiol. Biotechnol. 2018, 34, 76. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Li, S.; Jiang, Y.; Hu, X.; Wu, H. Necrotizing Enterocolitis and Intestinal Microbiota: The Timing of Disease and Combined Effects of Multiple Species. Front. Pediatr. 2021, 9, 657349. [Google Scholar] [CrossRef]
- Zuo, T.; Zhang, F.; Lui, G.C.Y.; Yeoh, Y.K.; Li, A.Y.L.; Zhan, H.; Wan, Y.; Chung, A.C.K.; Cheung, C.P.; Chen, N.; et al. Alterations in Gut Microbiota of Patients With COVID-19 During Time of Hospitalization. Gastroenterology 2020, 159, 944–955. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Kukreti, R.; Saso, L.; Kukreti, S. Oxidative Stress: A Key Modulator in Neurodegenerative Diseases. Molecules 2019, 24, 1583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isaguliants, M.; Smirnova, O.; Ivanov, A.V.; Kilpelainen, A.; Kuzmenko, Y.; Petkov, S.; Latanova, A.; Krotova, O.; Engström, G.; Karpov, V.; et al. Oxidative stress induced by HIV-1 reverse transcriptase modulates the enzyme’s performance in gene immunization. Hum. Vaccines Immunother. 2013, 9, 2111–2119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khomich, O.A.; Kochetkov, S.N.; Bartosch, B.; Ivanov, A.V. Redox Biology of Respiratory Viral Infections. Viruses 2018, 10, 392. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Rong, L.; Li, Y.P. Flaviviridae Viruses and Oxidative Stress: Implications for Viral Pathogenesis. Oxidative Med. Cell. Longev. 2019, 2019, 1409582. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Chen, F.; Liu, T.; Chen, F.; Liu, S.; Yang, J. The role of oxidative stress in influenza virus infection. Microbes Infect. 2017, 19, 580–586. [Google Scholar] [CrossRef]
- McCord, J.M.; Edeas, M.A. SOD, oxidative stress and human pathologies: A brief history and a future vision. Biomed. Pharmacother. 2005, 59, 139–142. [Google Scholar] [CrossRef]
- Gargouri, B.; Mseddi, M.; Mnif, F.; Abid, M.; Attia, H.; Lassoued, S. Oxidative stress enhances the immune response to oxidatively modified catalase enzyme in patients with Graves’ disease. J. Clin. Lab. Anal. 2020, 34, e23051. [Google Scholar] [CrossRef]
- Rehman, Z.U.; Meng, C.; Sun, Y.; Safdar, A.; Pasha, R.H.; Munir, M.; Ding, C. Oxidative Stress in Poultry: Lessons from the Viral Infections. Oxidative Med. Cell. Longev. 2018, 2018, 5123147. [Google Scholar] [CrossRef] [PubMed]
- Del Rio, D.; Stewart, A.J.; Pellegrini, N. A review of recent studies on malondialdehyde as toxic molecule and biological marker of oxidative stress. Nutr. Metab. Cardiovasc. Dis. 2005, 15, 316–328. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, K.B. Oxidative stress during viral infection: A review. Free Radic. Biol. Med. 1996, 21, 641–649. [Google Scholar] [CrossRef]
- Fang, F.C. Antimicrobial actions of reactive oxygen species. mBio 2011, 2, e00141-11. [Google Scholar] [CrossRef] [Green Version]
- Paiva, C.N.; Bozza, M.T. Are reactive oxygen species always detrimental to pathogens? Antioxid. Redox Signal. 2014, 20, 1000–1037. [Google Scholar] [CrossRef] [Green Version]
- Domingo, E. Rapid evolution of viral RNA genomes. J. Nutr. 1997, 127, 958s–961s. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.Q.; Lin, T.L.; Hu, L.Q.; Li, Y.Y.; Zhou, W.M.; Wu, Z.Y. Virus isolation and identification of Muscovy ducklings parvovirus disease. Chin. J. Virol. 1993, 9, 228–235. [Google Scholar]
- Xiao, S.; Chen, S.; Cheng, X.; Lin, F.; Wang, S.; Zhu, X.; Yu, B.; Huang, M.; Wang, J.; Wu, N.; et al. The newly emerging duck-origin goose parvovirus in China exhibits a wide range of pathogenicity to main domesticated waterfowl. Vet. Microbiol. 2017, 203, 252–256. [Google Scholar] [CrossRef]
- Fang, D.Y.; Wang, Y.K.; Zhen, Y.M.; Zhou, Y.S.; Jiang, M.J.; Dong, G.X. Study on the pathogen of gosling plague and specific control. Sci. Agric. Sin. 1981, 4, 1–8, 97. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′–3′) | Reverse Primer (5′–3′) | Accession |
|---|---|---|---|
| ZO-1 | TTCGGGAAGCTGGGTTTCTC | CCAGCGTCTCTTGGTTCACT | 101791291 |
| Occludin | GGGACTACGGCTACGGTTTG | CACCAGCAGCCCCAACTG | XM 013109403 |
| Claudin-1 | GATACTCCTGGGCCTCGTTG | CGAGCCACTCTGTTGCCATA | 101797741 |
| IFN-α | CACCTCTTCGACACCCTCAG | AGGTGGTGGATGTGGTGC | JF894229.1 |
| IL-1β | TCACAGTCCTTCGACATCTTCG | CCTCACTTTCTGGCTGGATGAG | DQ393268 |
| IL-2 | GATGAGAACGTATCTAGTGTTCGG | CAGCTCTCGGCGAAATTCAG | AY193713.1 |
| IL-6 | CCAAGGTGACGGAGGAAGAC | GTTGCCAGATGCTTTGTGCT | 101798321 |
| MHC-II | GAGACCAAGGGGTTCTTCCA | TGCCGGTTGTAGATGTCTCTC | DQ490138.1 |
| GAPDH | GCCACACAGAAGACAGTGGA | GTCAGGTCCACGACAGAGAC | GU564233.1 |
| Primer Names | Primer Sequences (5′–3′) | Primer Positions | Fragment Sizes (nt) |
|---|---|---|---|
| ITR-1 | CTCATTGGAGGGTTCGTTCG | 1–20; 5052–5071 | 187 |
| ITR-2 | GCCCGATCAGCCTTGACAAC | 168–189; 4883–4904 | |
| ITR-3 | GCGCATGCGCCCGATCTGCCATGA | 186–209; 4863–4886 | 532 |
| ITR-4 | GATTTTGTCTGCCAGAGTAACC | 696–717 | |
| 1-F | CCCCATGGTTACTCTGGCAGACAAAA | 690–715 | 927 |
| 1-R | GGGAAGTTCTCATTAGTCCAGT | 1595–1616 | |
| 2-F | CCACCGGAAAGACCAACAT | 1532–1550 | 935 |
| 2-R | GGCTGCAGTCTCATACCAGTCTT | 2444–2466 | |
| 3-F | GCCTGGAGTGTGAAAGAGCTAATT | 2237–2260 | 867 |
| 3-R | GGGAATCGCAATGCCAATT | 3085–3103 | |
| 4-F | GAACCTGTGGCAGCACCTAACAT | 2992–3014 | 1670 |
| 4-R | GCGCGCCAGGAAATGGTTTAT | 4642–4662 | |
| ITR-5 | CCAAACCTGGGAGGTTTTGG | 4303–4322 | 584 |
| ITR-3 | GCGCATGCGCCCGATCTGCCATGA | 186–209; 4863–4886 |
| Virus Classification | Strain | Collection Year | GenBank Accession ID | Reference |
|---|---|---|---|---|
| rMDPV | ZW | 2006 | KY744743 | [24] |
| JH06 | 2006 | MH807697 | [28] | |
| JH10 | 2010 | MH807698 | [28] | |
| SAAS-SHNH | 2012 | KC171936 | [25] | |
| FJM3 | 2013 | KR075690 | [48] | |
| GDNX | 2016 | MH204100 | [22] | |
| MDPV | FM | Unpublished | U22967 | [49] |
| P | 1988 | JF926697 | - | |
| FZ91-30 | 1991 | KT865605 | [17] | |
| YY | 2000 | KX000918 | [50] | |
| GPV | B | Unpublished | U25749 | [51] |
| SYG61v | 1961 | KC996729 | [52] | |
| 82-0321 | 1982 | EU583390 | [53] | |
| PT | 1997 | JF926695 | [54] | |
| 06-0329 | 2006 | EU583391 | [53] | |
| LH | 2012 | KM272560 | [55] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, J.; Zhang, Y.; Hu, Z.; Zhang, L.; Shao, G.; Xie, Z.; Nie, Y.; Li, W.; Li, Y.; Chen, L.; et al. Recombinant Muscovy Duck Parvovirus Led to Ileac Damage in Muscovy Ducklings. Viruses 2022, 14, 1471. https://doi.org/10.3390/v14071471
He J, Zhang Y, Hu Z, Zhang L, Shao G, Xie Z, Nie Y, Li W, Li Y, Chen L, et al. Recombinant Muscovy Duck Parvovirus Led to Ileac Damage in Muscovy Ducklings. Viruses. 2022; 14(7):1471. https://doi.org/10.3390/v14071471
Chicago/Turabian StyleHe, Jiahui, Yukun Zhang, Zezhong Hu, Luxuan Zhang, Guanming Shao, Zi Xie, Yu Nie, Wenxue Li, Yajuan Li, Liyi Chen, and et al. 2022. "Recombinant Muscovy Duck Parvovirus Led to Ileac Damage in Muscovy Ducklings" Viruses 14, no. 7: 1471. https://doi.org/10.3390/v14071471
APA StyleHe, J., Zhang, Y., Hu, Z., Zhang, L., Shao, G., Xie, Z., Nie, Y., Li, W., Li, Y., Chen, L., Huang, B., Chu, F., Feng, K., Lin, W., Li, H., Chen, W., Zhang, X., & Xie, Q. (2022). Recombinant Muscovy Duck Parvovirus Led to Ileac Damage in Muscovy Ducklings. Viruses, 14(7), 1471. https://doi.org/10.3390/v14071471