
Characterisation of a Hepatitis C Virus Subtype 2a Cluster in Scottish PWID with a Suboptimal Response to Glecaprevir/Pibrentasvir Treatment

 , , , and
, , , and 
Abstract
:1. Introduction
2. Methods
2.1. Clinical Cohort and Samples
2.2. HCV RNA Quantification
2.3. Library Preparation and Next Generation Sequencing
2.4. Bioinformatic Analyses
2.5. Sub-Genomic Replicon (SGR) Constructs
2.6. RNA Transcription and Electroporation
2.7. Replication Capacity of SGR Constructs
2.8. Sub-Genomic Replication Inhibition Assay
3. Results
3.1. Individual Characteristics
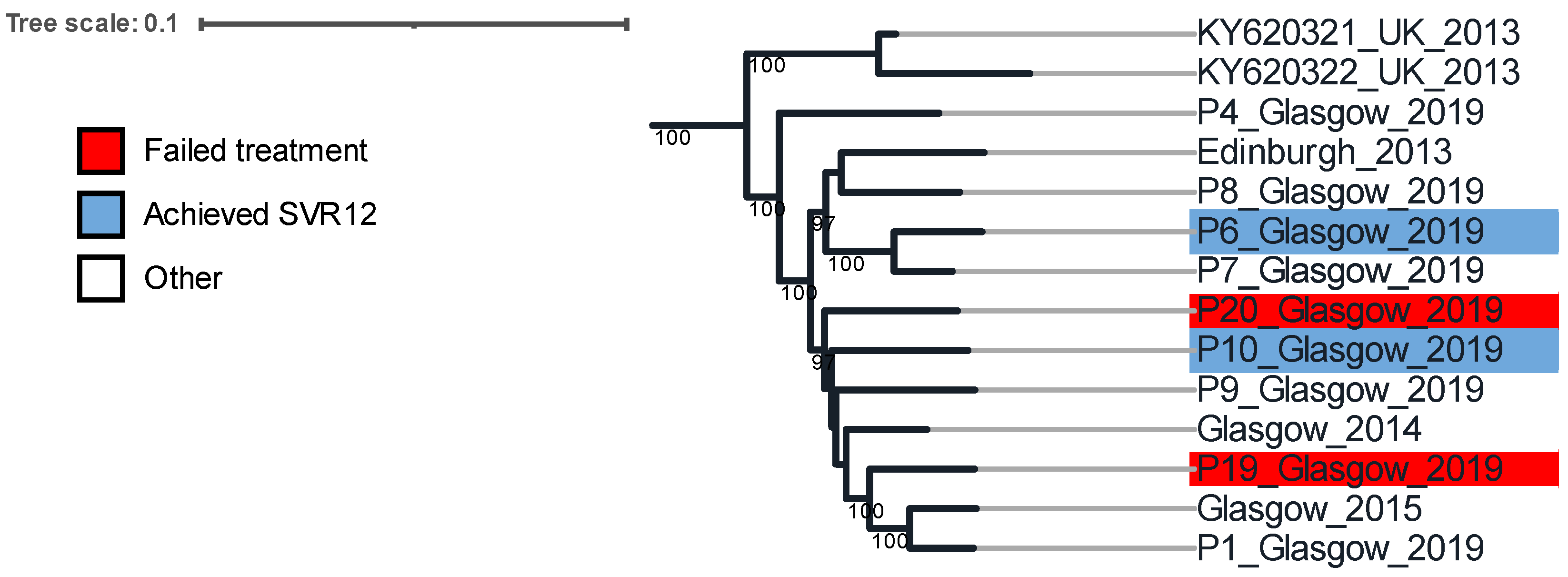
3.2. Phylogenetic Analysis
3.3. Resistance-Associated Substitutions
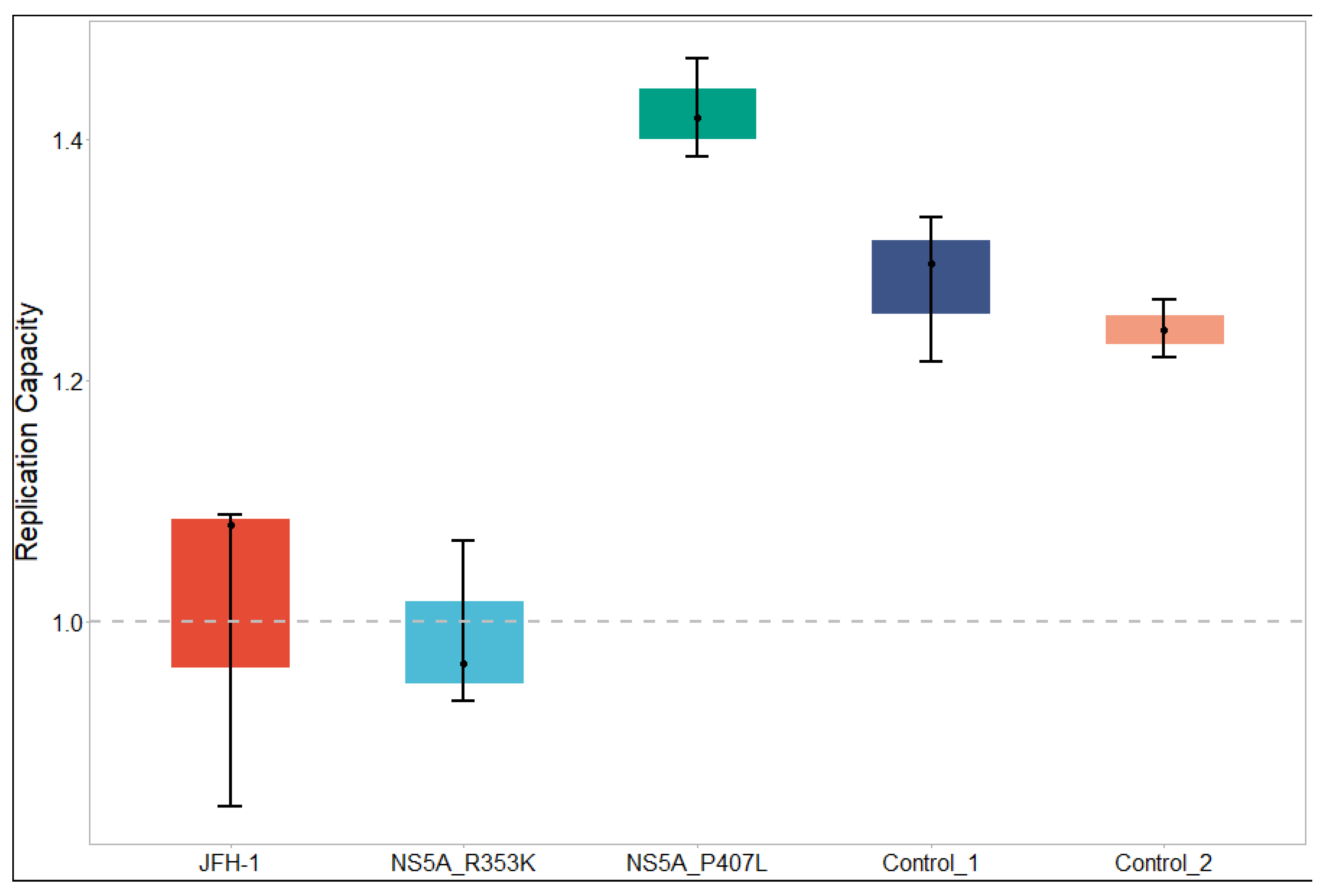
3.4. Replication Capacities of Sub-Genomic Replicon (SGR) Constructs
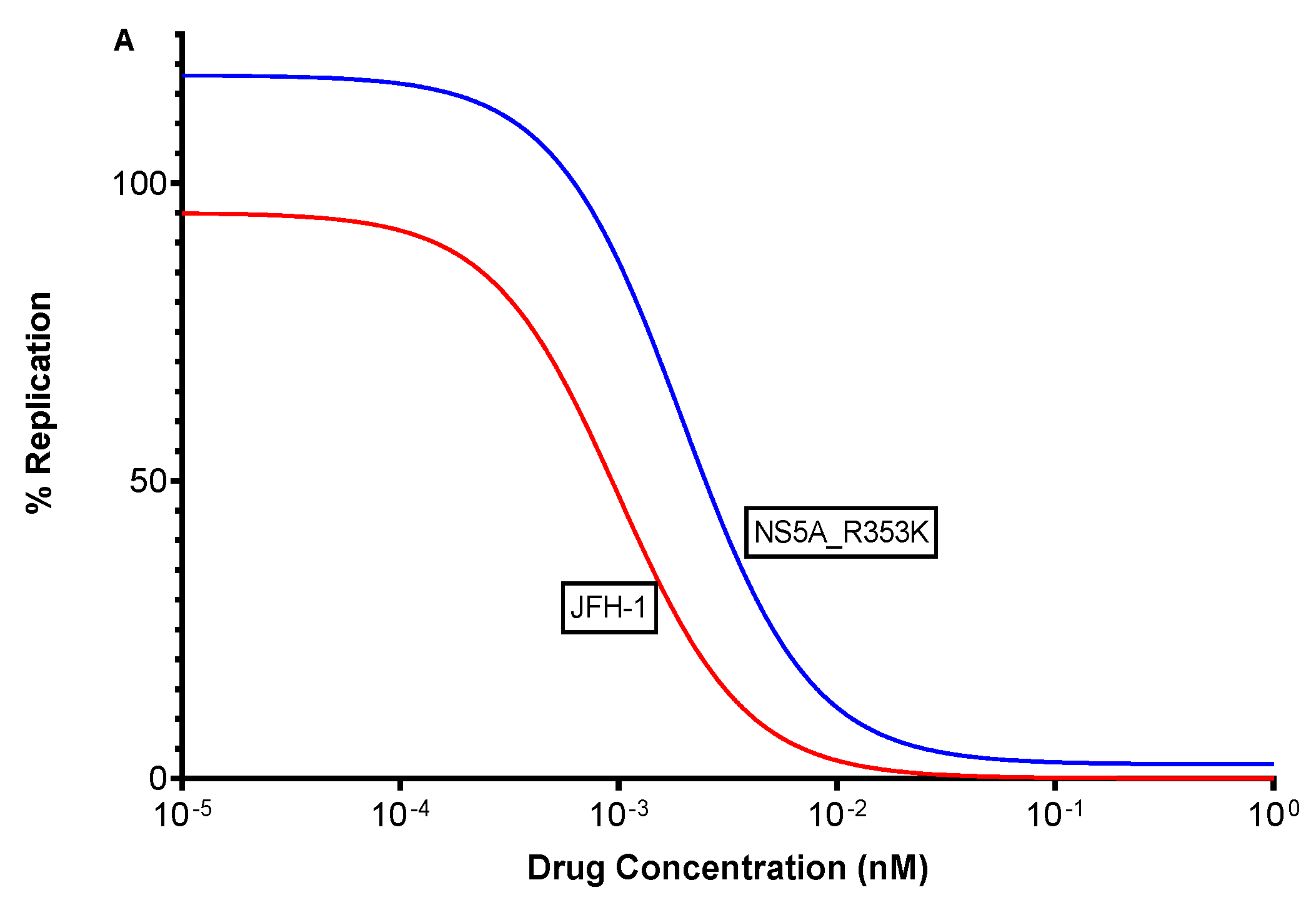
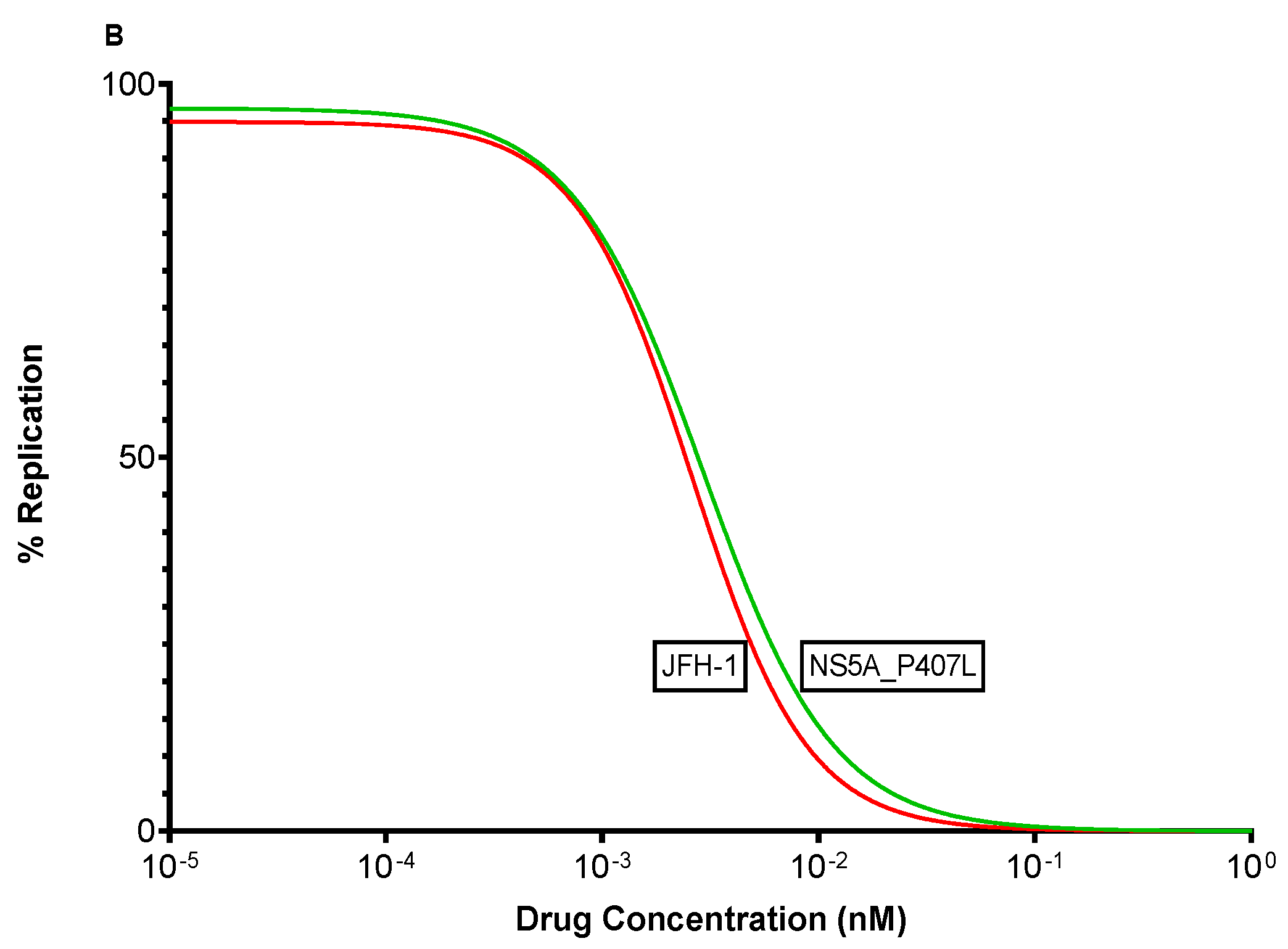
3.5. In Vitro Efficacy of Pibrentasvir
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

Appendix B

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sequence Identifier | Accession Number |
|---|---|
| P1 | OP022902 |
| P2 | OP022903 |
| P3 | OP022904 |
| P4 | OP022905 |
| P5 | OP022906 |
| P6 | OP022907 |
| P7 | OP022908 |
| P8 | OP022909 |
| P9 | OP022910 |
| P10 | OP022911 |
| P11 | OP022912 |
| P12 | OP022913 |
| P13 | OP022914 |
| P14 | OP022915 |
| P15 | OP022916 |
| P16 | OP022917 |
| P17 | OP022918 |
| P18 | OP022919 |
| P19 | OP022920 |
| P20 | OP022921 |
References
- WHO. Global Hepatitis Report; WHO: Geneva, Switzerland, 2017. [Google Scholar]
- WHO. Global Health Sector Strategy on Viral Hepatitis 2016–2021 towards Ending Viral Hepatitis; World Health Organisation: Geneva, Switzerland, 2016. [Google Scholar]
- Pawlotsky, J.M. New Hepatitis C Therapies: The Toolbox, Strategies, and Challenges. Gastroenterology 2014, 146, 1176–1192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pawlotsky, J.-M. Hepatitis C Virus Resistance to Direct-Acting Antiviral Drugs in Interferon-Free Regimens. Gastroenterology 2016, 151, 70–86. [Google Scholar] [CrossRef] [Green Version]
- Lontok, E.; Harrington, P.R.; Howe, A.; Kieffer, T.L.; Lennerstrand, J.; Lenz, O.; McPhee, F.; Mo, H.; Parkin, N.T.; Pilot-Matias, T.; et al. Hepatitis C virus drug resistance-associated substitutions: State of the art summary. Hepatology 2015, 62, 1623–1632. [Google Scholar] [CrossRef] [PubMed]
- European Association for the Study of the Liver. EASL Recommendations on Treatment of Hepatitis C 2018. J. Hepatol. 2018, 69, 461–511. [Google Scholar] [CrossRef] [Green Version]
- AASLD-IDSA HCV Guidance Panel. Hepatitis C Guidance 2018 Update: AASLD-IDSA Recommendations for Testing, Managing, and Treating Hepatitis C Virus Infection. Clin. Infect. Dis. 2018, 67, 1477–1492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. Guidelines for the Care and Treatment of Persons Diagnosed with Chronic Hepatitis C Virus Infection; World Health Organisation: Geneva, Switzerland, 2018. [Google Scholar]
- Esposito, I.; Trinks, J.; Soriano, V. Hepatitis C virus resistance to the new direct-acting antivirals. Expert Opin. Drug Metab. Toxicol. 2016, 12, 1197–1209. [Google Scholar] [CrossRef] [PubMed]
- Thomson, E.; Ip, C.L.C.; Badhan, A.; Christiansen, M.T.; Adamson, W.; Ansari, M.A.; Bibby, D.; Breuer, J.; Brown, A.; Bowden, R.; et al. Comparison of Next-Generation Sequencing Technologies for Comprehensive Assessment of Full-Length Hepatitis C Viral Genomes. J. Clin. Microbiol. 2016, 54, 2470–2484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kazutaka, K.; Misakwa, K.; Kei-ichi, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Singer, J.B.; Thomson, E.C.; McLauchlan, J.; Hughes, J.; Gifford, R.J. GLUE: A flexible software system for virus sequence data. BMC Bioinform. 2018, 19, 532. [Google Scholar] [CrossRef] [Green Version]
- Pilon, R.; Leonard, L.; Kim, J.; Vallée, D.; De Rubeis, E.; Jolly, A.M.; Wylie, J.; Pelude, L.; Sandstrom, P. Transmission Patterns of HIV and Hepatitis C Virus among Networks of People Who Inject Drugs. PLoS ONE 2011, 6, e22245. [Google Scholar] [CrossRef]
- Bamford, C.G.G.; Aranday-Cortes, E.; Filipe, I.C.; Sukumar, S.; Mair, D.; Filipe, A.D.S.; Mendoza, J.L.; Garcia, K.C.; Fan, S.; Tishkoff, S.A.; et al. A polymorphic residue that attenuates the antiviral potential of interferon lambda 4 in hominid lineages. PLoS Pathog. 2018, 14, e1007307. [Google Scholar] [CrossRef] [Green Version]
- Kato, T.; Furusaka, A.; Miyamoto, M.; Date, T.; Yasui, K.; Hiramoto, J.; Nagayama, K.; Tanaka, T.; Wakita, T. Sequence analysis of hepatitis C virus isolated from a fulminant hepatitis patient. J. Med. Virol. 2001, 64, 334–339. [Google Scholar] [CrossRef]
- NCBI Resource Coordinators. Database resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2016, 44, D7–D19. [Google Scholar] [CrossRef] [Green Version]
- Ng, T.I.; Tripathi, R.; Reisch, T.; Lu, L.; Middleton, T.; Hopkins, T.A.; Pithawalla, R.; Irvin, M.; Dekhtyar, T.; Krishnan, P.; et al. In Vitro Antiviral Activity and Resistance Profile of the Next-Generation Hepatitis C Virus NS3/4A Protease Inhibitor Glecaprevir. Antimicrob. Agents Chemother. 2018, 62, e01620-17. [Google Scholar] [CrossRef] [Green Version]
- Pawlotsky, J.M.; Negro, F.; Aghemo, A.; Berenguer, M.; Dalgard, O.; Dusheiko, G.; Marra, F.; Puoti, M.; Wedemeyer, H.; European Association for the Study of the Liver. EASL recommendations on treatment of hepatitis C: Final update of the series. J. Hepatol. 2020, 73, 1170–1218. [Google Scholar] [CrossRef]
- Brown, R.S.; Buti, M.; Rodrigues, L.; Chulanov, V.; Chuang, W.-L.; Aguilar, H.; Horváth, G.; Zuckerman, E.; Carrion, B.R.; Rodriguez-Perez, F.; et al. Glecaprevir/pibrentasvir for 8 weeks in treatment-naïve patients with chronic HCV genotypes 1–6 and compensated cirrhosis: The EXPEDITION-8 trial. J. Hepatol. 2019, 72, 441–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asselah, T.; Kowdley, K.V.; Zadeikis, N.; Wang, S.; Hassanein, T.; Horsmans, Y.; Colombo, M.; Calinas, F.; Aguilar, H.; de Ledinghen, V.; et al. Efficacy of Glecaprevir/Pibrentasvir for 8 or 12 Weeks in Patients with Hepatitis C Virus Genotype 2, 4, 5, or 6 Infection Without Cirrhosis. Clin. Gastroenterol. Hepatol. 2018, 16, 417–426. [Google Scholar] [CrossRef]
- Kwo, P.Y.; Poordad, F.; Asatryan, A.; Wang, S.; Wyles, D.L.; Hassanein, T.; Felizarta, F.; Sulkowski, M.S.; Gane, E.; Maliakkal, B.; et al. Glecaprevir and pibrentasvir yield high response rates in patients with HCV genotype 1–6 without cirrhosis. J. Hepatol. 2017, 67, 263–271. [Google Scholar] [CrossRef]
- Zhou, N.; Han, Z.; Hartman-Neumann, S.; DeGray, B.; Ueland, J.; Vellucci, V.; Hernandez, D.; McPhee, F. Characterization of NS5A polymorphisms and their impact on response rates in patients with HCV genotype 2 treated with daclatasvir-based regimens. J. Antimicrob. Chemother. 2016, 71, 3495–3505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bourlière, M.; Gordon, S.C.; Flamm, S.L.; Cooper, C.L.; Ramji, A.; Tong, M.; Ravendhran, N.; Vierling, J.M.; Tran, T.T.; Pianko, S.; et al. Sofosbuvir, Velpatasvir, and Voxilaprevir for Previously Treated HCV Infection. N. Engl. J. Med. 2017, 376, 2134–2146. [Google Scholar] [CrossRef] [PubMed]
- Gottwein, J.M.; Pham, L.V.; Mikkelsen, L.S.; Ghanem, L.; Ramirez, S.; Scheel, T.K.; Carlsen, T.H.; Bukh, J. Efficacy of NS5A Inhibitors Against Hepatitis C Virus Genotypes 1–7 and Escape Variants. Gastroenterology 2018, 154, 1435–1448. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, P.; Beyer, J.; Mistry, N.; Koev, G.; Reisch, T.; DeGoey, D.; Kati, W.; Campbell, A.; Williams, L.; Xie, W.; et al. In Vitro and In Vivo Antiviral Activity and Resistance Profile of Ombitasvir, an Inhibitor of Hepatitis C Virus NS5A. Antimicrob. Agents Chemother. 2015, 59, 979–987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atsukawa, M.; Tsubota, A.; Toyoda, H.; Takaguchi, K.; Nakamuta, M.; Watanabe, T.; Tada, T.; Tsutsui, A.; Ikeda, H.; Abe, H.; et al. Efficacy and safety of ombitasvir/paritaprevir/ritonavir and ribavirin for chronic hepatitis patients infected with genotype 2a in Japan. Hepatol. Res. 2018, 49, 369–376. [Google Scholar] [CrossRef] [PubMed]
- Lambert, S.; Langley, D.R.; Garnett, J.A.; Angell, R.; Hedgethorne, K.; Meanwell, N.; Matthews, S.J. The crystal structure of NS5A domain 1 from genotype 1a reveals new clues to the mechanism of action for dimeric HCV inhibitors. Protein Sci. 2014, 23, 723–734. [Google Scholar] [CrossRef] [Green Version]
- Appel, N.; Zayas, M.; Miller, S.; Krijnse-Locker, J.; Schaller, T.; Friebe, P.; Kallis, S.; Engel, U.; Bartenschlager, R. Essential Role of Domain III of Nonstructural Protein 5A for Hepatitis C Virus Infectious Particle Assembly. PLoS Pathog. 2008, 4, e1000035. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Welsch, C.; Yi, M.; Lemon, S.M. Regulation of the Production of Infectious Genotype 1a Hepatitis C Virus by NS5A Domain III. J. Virol. 2011, 85, 6645–6656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foster, T.L.; Belyaeva, T.; Stonehouse, N.J.; Pearson, A.R.; Harris, M. All Three Domains of the Hepatitis C Virus Nonstructural NS5A Protein Contribute to RNA Binding. J. Virol. 2010, 84, 9267–9277. [Google Scholar] [CrossRef] [Green Version]




| Treatment Outcome (N = 123) | p Value | ||
|---|---|---|---|
| SVR * (N = 112) | Failure (N = 11) | ||
| Gender | |||
| Male | 80 (71%) | 11 (100%) | |
| Female | 32(29%) | 0 (0%) | 0.0648 † |
| Mean Age (standard deviation) | 48.8 (±10) | 47.5 (±10) | 0.8731 †† |
| Mean HCV Viral Load (IU/mL) | 5.88 (±0.9) | 5.98 (±0.7) | 0.8593 †† |
| HIV Status | |||
| HIV negative | 107 (96%) | 11 (100%) | 1 † |
| HIV positive | 5 (4%) | 0 (0%) | |
| Liver Status | |||
| Non-Cirrhotic | 96 (86%) | 11 (100%) | 0.356 † |
| Cirrhosis (>12KPa on transient elastography) | 16 (14%) | 0 (0%) | |
| Transmission | |||
| IDU ** | 78 (69%) | 8 (73%) | 0.6724 † |
| Other *** | 10 (9%) | 0 (0%) | |
| Unknown | 24 (22%) | 3 (27%) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shah, R.; Barclay, S.T.; Peters, E.S.; Fox, R.; Gunson, R.; Bradley-Stewart, A.; Shepherd, S.J.; MacLean, A.; Tong, L.; van Vliet, V.J.E.; et al. Characterisation of a Hepatitis C Virus Subtype 2a Cluster in Scottish PWID with a Suboptimal Response to Glecaprevir/Pibrentasvir Treatment. Viruses 2022, 14, 1678. https://doi.org/10.3390/v14081678
Shah R, Barclay ST, Peters ES, Fox R, Gunson R, Bradley-Stewart A, Shepherd SJ, MacLean A, Tong L, van Vliet VJE, et al. Characterisation of a Hepatitis C Virus Subtype 2a Cluster in Scottish PWID with a Suboptimal Response to Glecaprevir/Pibrentasvir Treatment. Viruses. 2022; 14(8):1678. https://doi.org/10.3390/v14081678
Chicago/Turabian StyleShah, Rajiv, Stephen T. Barclay, Erica S. Peters, Ray Fox, Rory Gunson, Amanda Bradley-Stewart, Samantha J. Shepherd, Alasdair MacLean, Lily Tong, Vera Jannie Elisabeth van Vliet, and et al. 2022. "Characterisation of a Hepatitis C Virus Subtype 2a Cluster in Scottish PWID with a Suboptimal Response to Glecaprevir/Pibrentasvir Treatment" Viruses 14, no. 8: 1678. https://doi.org/10.3390/v14081678
APA StyleShah, R., Barclay, S. T., Peters, E. S., Fox, R., Gunson, R., Bradley-Stewart, A., Shepherd, S. J., MacLean, A., Tong, L., van Vliet, V. J. E., Ngan Chiu Bong, M., Filipe, A., Thomson, E. C., & Davis, C. (2022). Characterisation of a Hepatitis C Virus Subtype 2a Cluster in Scottish PWID with a Suboptimal Response to Glecaprevir/Pibrentasvir Treatment. Viruses, 14(8), 1678. https://doi.org/10.3390/v14081678