
Omicron Sub-Lineages (BA.1.1.529 + BA.*) Current Status in Ecuador

 ,
,  , ,
, ,  , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sequence Production and Data Collection
2.2. Lineage Assignment and Phylogenetics
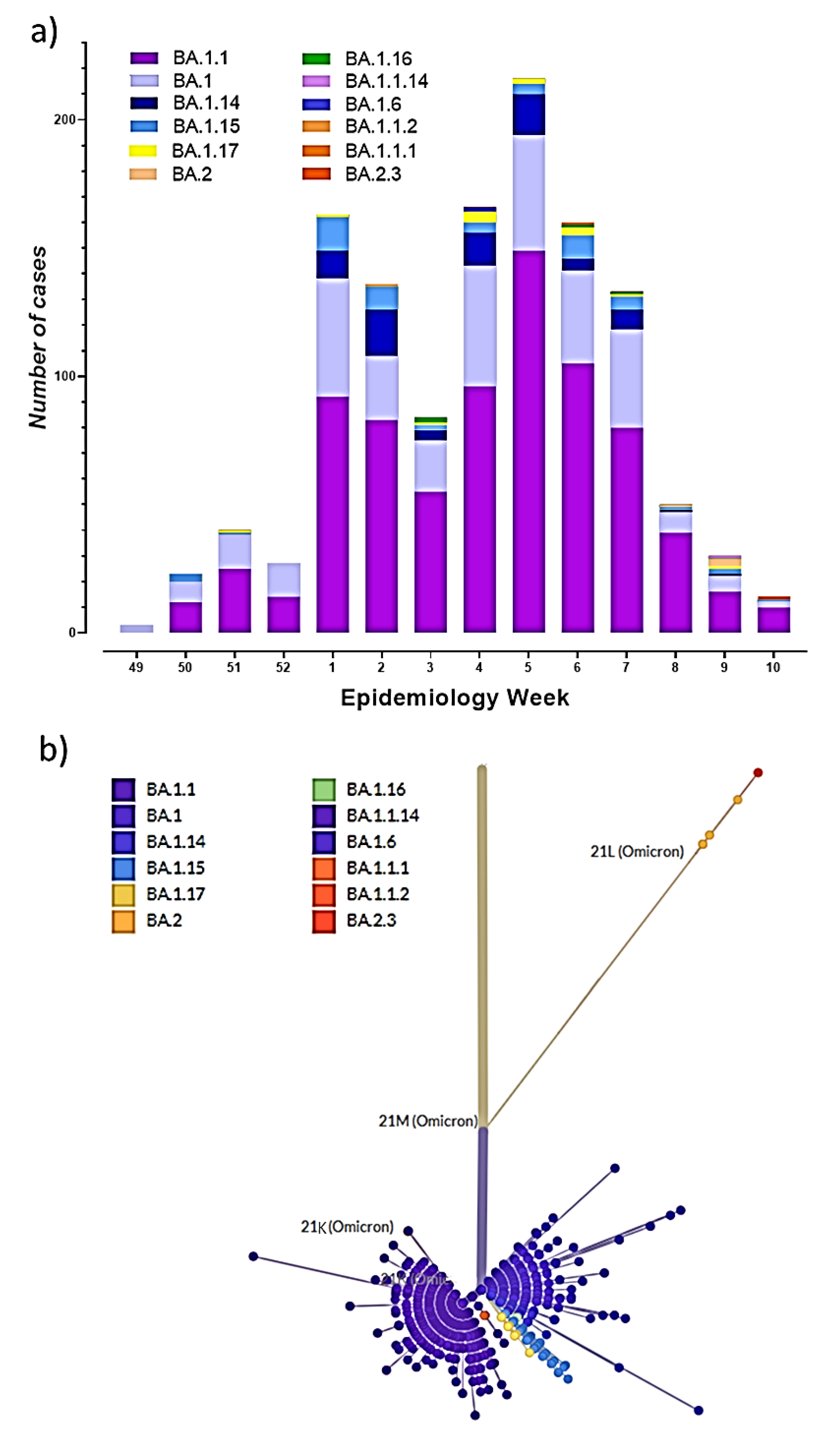
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wu, F.; Zhao, S.; Yu, B.; Chen, Y.; Wang, W.; Song, Z.; Hu, Y.; Tao, Z.; Tian, J.; Pei, Y.; et al. A new coronavirus associated with human respiratory disease in China. Nature 2020, 579, 265–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. WHO Coronavirus (COVID-19) Dashboard. Available online: https://covid19.who.int/ (accessed on 7 April 2022).
- Hu, B.; Guo, H.; Zhou, P.; Shi, Z. Characteristics of SARS-CoV-2 and COVID-19. Nat. Rev. Microbiol. 2021, 19, 141–154. [Google Scholar] [CrossRef] [PubMed]
- Khandia, R.; Singhal, S.; Alqahtani, T.; Kamal, M.A.; El-Shall, N.; Nainu, F.; Desingu, P.; Dhama, K. Emergence of SARS-CoV-2 Omicron (B.1.1.529) variant, salient features, high global health concerns and strategies to counter it amid ongoing COVID-19 pandemic. Environ. Res. 2022, 209, 112816. [Google Scholar] [CrossRef] [PubMed]
- Ito, K.; Piantham, C.; Nishiura, H. Relative instantaneous reproduction number of Omicron SARS-CoV-2 variant with respect to the Delta variant in Denmark. J. Med. Virol. 2022, 94, 2265–2268. [Google Scholar] [CrossRef]
- Nishiura, H.; Ito, K.; Anzai, A.; Kobayashi, T.; Piantham, C.; Rodríguez-Morales, A.J. Relative reproduction number of SARS-CoV-2 Omicron (B.1.1.529) compared with Delta variant in South Africa. J. Clin. Med. 2021, 11, 30. [Google Scholar] [CrossRef]
- Jung, C.; Kmiec, D.; Koepke, L.; Zech, F.; Jacob, T.; Sparrer, K.; Kirchhoff, F. Omicron: What makes the latest SARS-CoV-2 variant of concern so concerning? J. Virol. 2022, 96, e02077-21. [Google Scholar] [CrossRef]
- Lupala, C.; Ye, Y.; Chen, H.; Su, X.D.; Liu, H. Mutations on RBD of SARS-CoV-2 Omicron variant result in stronger binding to human ACE2 receptor. Biochem. Biophys. Res. Commun. 2022, 590, 34–41. [Google Scholar] [CrossRef]
- Emergence of Sub-Lineage and Recombination Events: Genetic Evolution of SARS-CoV-2. Available online: https://www.paho.org/en/documents/emergence-sub-lineage-and-recombination-events-genetic-evolution-sars-cov-2-13-april-2022 (accessed on 13 April 2022).
- Carrazco, A.; Armendáriz, I.; Tello, C.; Morales, D.; Armas, R.; Guizado, D.; León, A.; Ramos, D.; Fuentes, B.; USFQ-Consortium; et al. First detection of SARS-CoV-2 variant B.1.1.529 (Omicron) in Ecuador. New Microbes New Infect. 2021, 45, 100951. [Google Scholar] [CrossRef]
- Ministerio de Salud Pública. Available online: https://www.salud.gob.ec/informes-de-situacion-sitrep-e-infografias-covid-19-desde-26-07-2021/ (accessed on 7 April 2022).
- Khare, S.; Gurry, C.; Freitas, L.; Schultz, M.; Bach, G.; Diallo, A.; Akite, N.; Ho, J.; Lee, R.; Yeo, W.; et al. GISAID’s Role in Pandemic Response. China CDC Wkly. 2021, 3, 1049–1051. [Google Scholar] [CrossRef]
- Resende, P.; Delatorre, E.; Gräf, T.; Mir, D.; Couto, F.; Reis, L.; Dias, A.; Da Fonseca, A.; Ogrzewalska, M.; Caetano, B.; et al. Evolutionary Dynamics and Dissemination Pattern of the the Early Pandemic Phase in Brazil. Front. Microbiol. 2021, 11, 615280. [Google Scholar] [CrossRef]
- Aksamentov, I.; Roemer, C.; Hodcroft, E.; Neher, R. Nextclade: Clade assignment, mutation calling and quality control for viral genomes. J. Open Source Softw. 2021, 6, 3773. [Google Scholar] [CrossRef]
- Rambaut, A.; Holmes, E.; O’Toole, Á.; Verity, H.; McCrone, J.; Ruis, C.; du Plessis, L.; Pybus, O. A dynamic nomenclature proposal for SARS-CoV-2 lineages to assist genomic epidemiology. Nat. Microbiol. 2020, 5, 1403–1407. [Google Scholar] [CrossRef] [PubMed]
- González, F.; Shaw, M.; Phan, T.; Kulkarni, U.; Paraskevis, D.; Luciani, F.; Kimura, H.; Sironi, M. One year into the pandemic: Short-term evolution of SARS-CoV-2 and emergence of new lineages. Infect. Genet. Evol. 2021, 92, 104869. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 14 April 2022).
- GraphPad Software. Available online: www.graphpad.com (accessed on 13 April 2022).
- The Maravi Post. Expansion of L452R-Positive SARS-CoV-2 Omicron Variant, Northern Lombardy, Italy. Available online: https://www.maravipost.com/expansion-of-l452r-positive-sars-cov-2-omicron-variant-northern-lombardy-italy/ (accessed on 14 April 2022).
- Mallapaty, S. Where did Omicron come from? Three key theories. Nature 2022, 602, 26–28. [Google Scholar] [CrossRef]
- Thomas, L. Omicron BA.1.1 and BA.2 Subvariants with New Mutations in New Zealand and Hong Kong. 2022. Available online: https://www.news-medical.net/news/20220320/Omicron-BA11-and-BA2-subvariants-with-new-mutations-in-New-Zealand-and-Hong-Kong.aspx (accessed on 13 May 2022).
- Genomic Overview of SARS-CoV-2 in Denmark. Available online: https://www.covid19genomics.dk/statistics (accessed on 8 April 2022).
- Chen, L.; Abdullah, M.; Chan, W.; Chan, B.; Jonathan, D.; Chu, W.; Lu, L.; Zhang, X.; Zhao, Y.; Chuang, W.; et al. Contribution of low population immunity to the severe Omicron BA.2 outbreak in Hong Kong. 2022; manuscript in preparation. Available online: https://www.researchsquare.com/article/rs-1512533/v1(accessed on 18 April 2022).
- UNAM. Programa de Vigilancia Genómica de SARS-CoV-2 Realizado por el CoViGen-Mex. Available online: http://mexcov2.ibt.unam.mx:8080/COVID-TRACKER/open/reports/23%20de%20Marzo%20del%202022.pdf (accessed on 13 May 2022).
- Rapid Establishment of Omicron Variant in Rio Grande do Sul State, Brazil. Available online: https://virological.org/t/rapid-establishment-of-omicron-variant-in-rio-grande-do-sul-state-brazil/783 (accessed on 13 April 2022).
- Government of Chile. Epidemiology Department. Available online: https://www.minsal.cl/wp-content/uploads/2022/04/Informe_Variantes-N%C2%B028.pdf (accessed on 8 April 2022).
- Wu, C.R.; Yin, W.C.; Jiang, Y.; Xu, H.E. Structure genomics of SARS-CoV-2 and its Omicron variant: Drug design templates for COVID-19. Acta Pharmacol. Sin. 2022. [Google Scholar] [CrossRef]
- Wu, H.; Xing, N.; Meng, K.; Fu, B.; Xue, W.; Dong, P.; Tang, W.; Xiao, Y.; Liu, G.; Luo, H.; et al. Nucleocapsid mutations R203K/G204R increase the infectivity, fitness, and virulence of SARS-CoV-2. Cell Host Microbe 2021, 29, 1788–1801.e6. [Google Scholar] [CrossRef]
- UK Health Security Agency. SARS-CoV-2 Variants of Concern and Variants under Investigation in England. Technical Briefing 39. Available online: https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/1063424/Tech-Briefing-39-25March2022_FINAL.pdf (accessed on 13 May 2022).
- Callaway, F. Why does the Omicron sub-variant spread faster than the original? Nature 2022, 602, 556–557. [Google Scholar] [CrossRef]
- Colson, P.; Delerce, J.; Beye, M.; Levasseur, A.; Boschi, C.; Houhamdi, L.; Fournier, P.E. First cases of infection with the 21L/BA. 2 Omicron variant in Marseille, France. J. Med. Virol. 2022, 94, 3421–3430. [Google Scholar] [CrossRef]
- Ministerio de Salud Pública. Datos Epidemiológicos COVID-19. Available online: https://app.powerbi.com/view?r=eyJrIjoiNjkzNjUyNGMtZjc1NC00OWU0LWEzMWQtOTFkY2ZkYjZkMWIwIiwidCI6IjcwNjIyMGRiLTliMjktNGU5MS1hODI1LTI1NmIwNmQyNjlmMyJ9&pageName=ReportSection33e2bb803a8b183d9100 (accessed on 17 May 2022).
- Trilla, A. One world, one health: The novel coronavirus COVID-19 epidemic. Med. Clin. (Engl. Ed.) 2020, 154, 175–177. [Google Scholar] [CrossRef]
- Singh, J.; Pandit, P.; McArthur, A.G.; Banerjee, A.; Mossman, K. Evolutionary trajectory of SARS-CoV-2 and emerging variants. Virol. J. 2021, 18, 166. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Sub-Lineage | Clade Names | Origin | Genes | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| ORF1a | ORF1b | S | ORF3a | E | M | ORF6 | ORF8 | N | |||
| BA.1 | 21K (Omicron) | South Africa | 8 | 2 | 33 | 0 | 1 | 3 | 0 | 1 | 4 |
| BA.1.1 | 21K (Omicron) | South Africa | 8 | 2 | 32 | 0 | 1 | 3 | 0 | 1 | 4 |
| BA.1.1.1 | 21K (Omicron) | Europe | 8 | 2 | 31 | 0 | 1 | 3 | 0 | 1 | 4 |
| BA.1.1.14 | 21K (Omicron) | Europe | 8 | 2 | 30 | 0 | 1 | 3 | 0 | 1 | 4 |
| BA.1.1.2 | 21K (Omicron) | Japan | 9 | 2 | 34 | 0 | 1 | 3 | 0 | 1 | 4 |
| BA.1.14 | 21K (Omicron) | Brazil | 8 | 2 | 21 | 0 | 1 | 3 | 0 | 1 | 4 |
| BA.1.15 | 21K (Omicron) | USA | 8 | 2 | 30 | 1 | 1 | 3 | 0 | 1 | 5 |
| BA.1.16 | 21K (Omicron) | UK | 8 | 2 | 29 | 0 | 1 | 3 | 0 | 1 | 4 |
| BA.1.17 | 21K (Omicron) | Europe | 9 | 2 | 30 | 0 | 1 | 3 | 0 | 1 | 4 |
| BA.1.6 | 21K (Omicron) | Canada and Sint Maarten | 8 | 3 | 30 | 0 | 1 | 3 | 0 | 1 | 4 |
| BA.2 | 21L (Omicron) | India and South Africa | 9 | 4 | 29 | 1 | 1 | 2 | 1 | 1 | 5 |
| BA.2.3 | 21L (Omicron) | Philippines | 10 | 4 | 29 | 2 | 1 | 2 | 1 | 1 | 5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carrazco-Montalvo, A.; Herrera-Yela, A.; Alarcón-Vallejo, D.; Gutiérrez-Pallo, D.; Armendáriz-Castillo, I.; Andrade-Molina, D.; Muñoz-Mawyin, K.; Fernández-Cadena, J.C.; Morey-León, G.; USFQ-COVID-19 Consortium; et al. Omicron Sub-Lineages (BA.1.1.529 + BA.*) Current Status in Ecuador. Viruses 2022, 14, 1177. https://doi.org/10.3390/v14061177
Carrazco-Montalvo A, Herrera-Yela A, Alarcón-Vallejo D, Gutiérrez-Pallo D, Armendáriz-Castillo I, Andrade-Molina D, Muñoz-Mawyin K, Fernández-Cadena JC, Morey-León G, USFQ-COVID-19 Consortium, et al. Omicron Sub-Lineages (BA.1.1.529 + BA.*) Current Status in Ecuador. Viruses. 2022; 14(6):1177. https://doi.org/10.3390/v14061177
Chicago/Turabian StyleCarrazco-Montalvo, Andrés, Andrés Herrera-Yela, Damaris Alarcón-Vallejo, Diana Gutiérrez-Pallo, Isaac Armendáriz-Castillo, Derly Andrade-Molina, Karen Muñoz-Mawyin, Juan Carlos Fernández-Cadena, Gabriel Morey-León, USFQ-COVID-19 Consortium, and et al. 2022. "Omicron Sub-Lineages (BA.1.1.529 + BA.*) Current Status in Ecuador" Viruses 14, no. 6: 1177. https://doi.org/10.3390/v14061177
APA StyleCarrazco-Montalvo, A., Herrera-Yela, A., Alarcón-Vallejo, D., Gutiérrez-Pallo, D., Armendáriz-Castillo, I., Andrade-Molina, D., Muñoz-Mawyin, K., Fernández-Cadena, J. C., Morey-León, G., USFQ-COVID-19 Consortium, CRN Influenza y OVR—INSPI, & Patiño, L. (2022). Omicron Sub-Lineages (BA.1.1.529 + BA.*) Current Status in Ecuador. Viruses, 14(6), 1177. https://doi.org/10.3390/v14061177