
Low Susceptibility of Rubella Virus in First-Trimester Trophoblast Cell Lines

 , , and
, , and {kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Rubella Virus
2.3. Viral Infection
2.4. Immunofluorescence Assay
2.5. FCM Analysis
2.6. Statistical Analysis
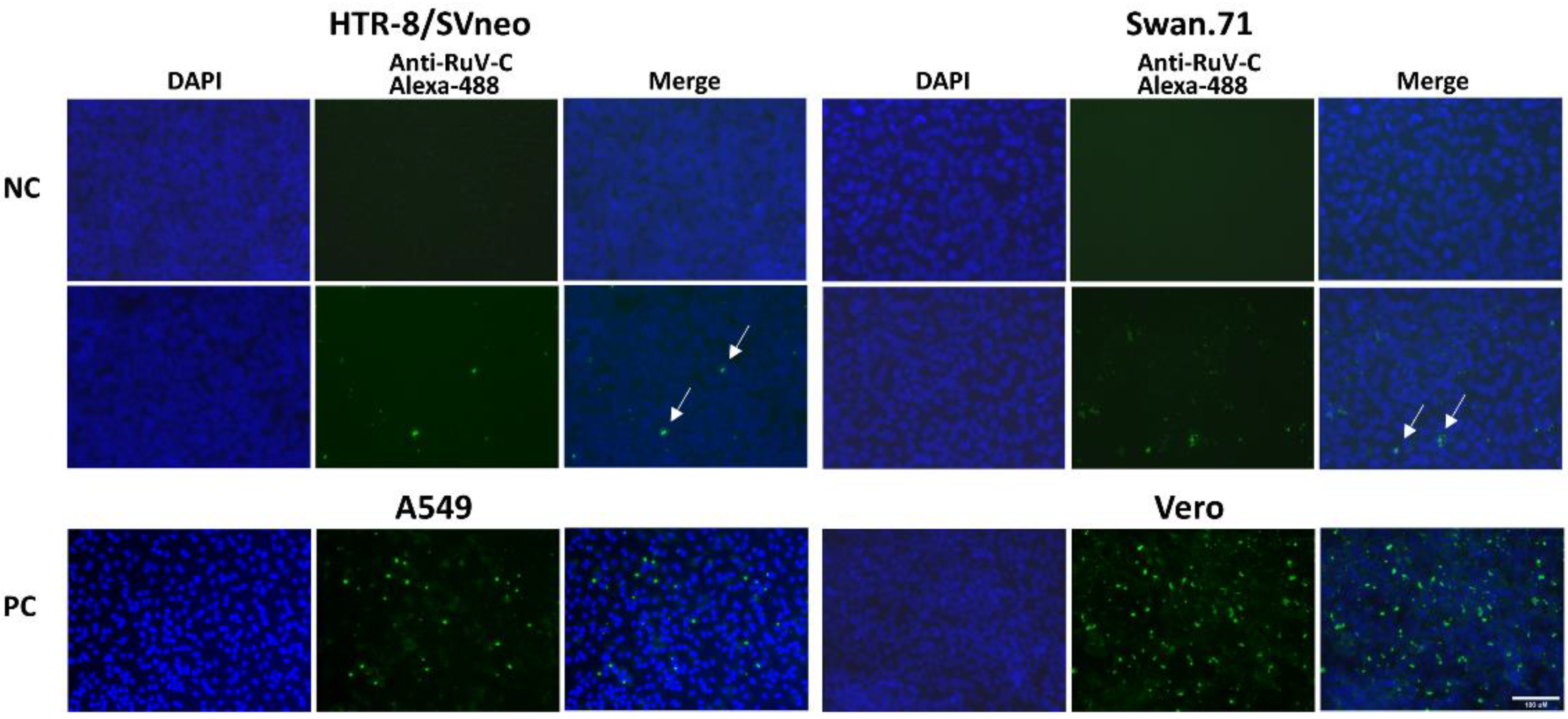
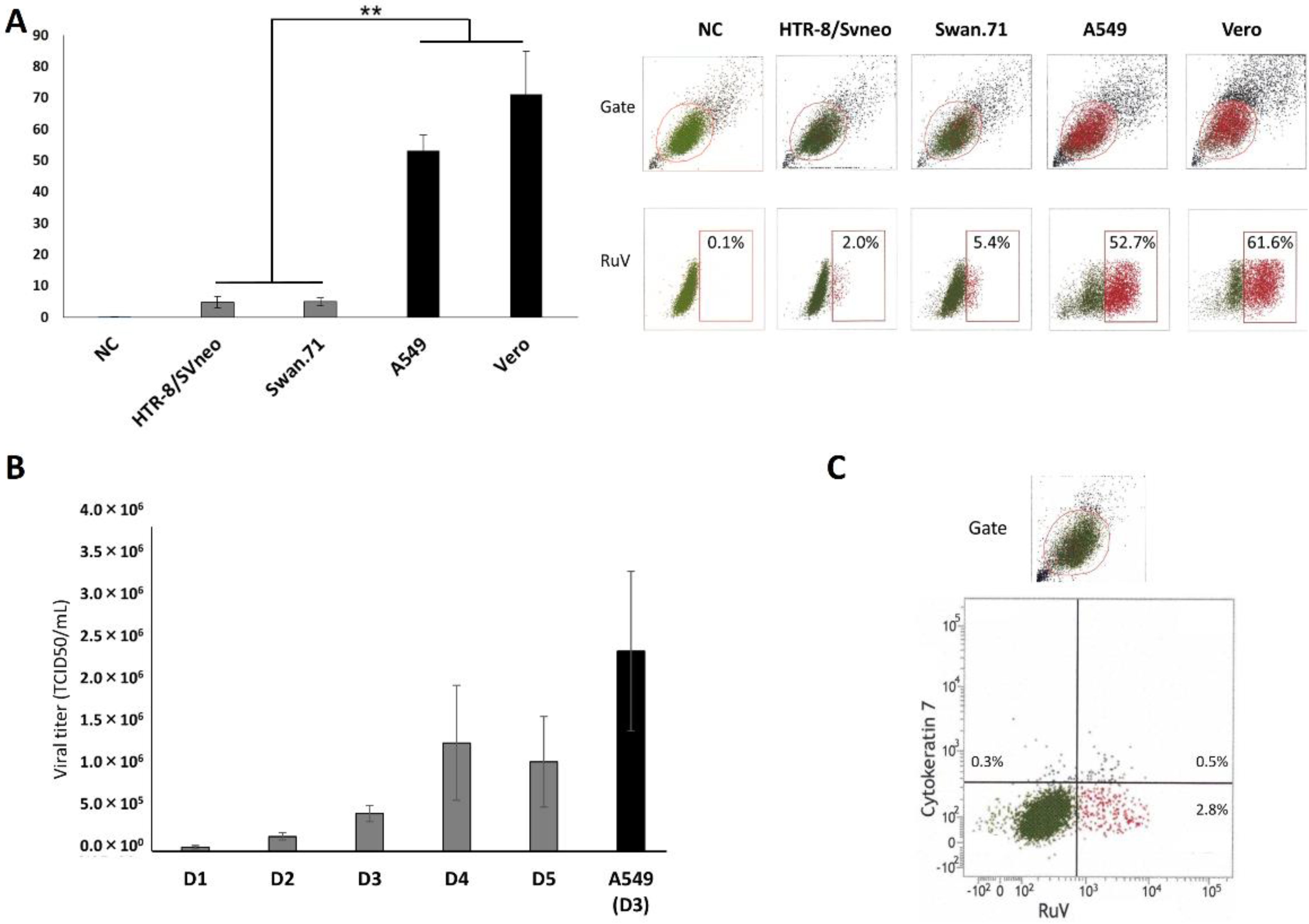
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Das, P.K.; Kielian, M. Molecular and Structural Insights into the Life Cycle of Rubella Virus. J. Virol. 2021, 95, e02349-20. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Mukhopadhyay, S.; Merits, A.; Bolling, B.; Nasar, F.; Coffey, L.; Powers, A.; Weaver, S.; Smith, D.; Simmonds, P.; et al. Create a new family Matonaviridae to include the genus Rubivirus, removed from the family Togaviridae. 2018. Available online: https://talk.ictvonline.org/files/ictv_official_taxonomy_updates_since_the_8th_report/m/animal-ssrna-viruses/8087 (accessed on 16 February 2022).
- Hobman, T.C. Rubella Virus. In Fields Virology, 6th ed.; Knipe, D.M., Howley, P., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2013; Volume 1, pp. 687–711. [Google Scholar]
- Winter, A.K.; Moss, W.J. Rubella. Lancet 2022, 399, 1336–1346. [Google Scholar] [CrossRef]
- Mawson, A.R.; Croft, A.M. Rubella Virus Infection, the Congenital Rubella Syndrome, and the Link to Autism. Int. J. Environ. Res. Public Health 2019, 16, 3543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Motaze, N.V.; Mthombothi, Z.E.; Adetokunboh, O.; Hazelbag, C.M.; Saldarriaga, E.M.; Mbuagbaw, L.; Wiysonge, C.S. The Impact of Rubella Vaccine Introduction on Rubella Infection and Congenital Rubella Syndrome: A Systematic Review of Mathematical Modelling Studies. Vaccines 2021, 9, 84. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Verma, S.; Dash, N.; Goel, M.; Rawat, A.; Singh, M.P.; Rohit, M.K.; Kumar, P. Congenital Rubella: A Salient Cause of Congenital Heart Defects in Infants. J. Trop. Pediatr. 2021, 67, fmab044. [Google Scholar] [CrossRef] [PubMed]
- Yazigi, A.; De Pecoulas, A.E.; Vauloup-Fellous, C.; Grangeot-Keros, L.; Ayoubi, J.M.; Picone, O. Fetal and neonatal abnormalities due to congenital rubella syndrome: A review of literature. J. Matern.-Fetal Neonatal Med. 2017, 30, 274–278. [Google Scholar] [CrossRef] [PubMed]
- Bayer, A.; Delorme-Axford, E.; Sleigher, C.; Frey, T.K.; Trobaugh, D.W.; Klimstra, W.B.; Emert-Sedlak, L.A.; Smithgall, T.E.; Kinchington, P.R.; Vadia, S.; et al. Human trophoblasts confer resistance to viruses implicated in perinatal infection. Am. J. Obstet. Gynecol. 2015, 212, 71.e1–71.e8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trinh, Q.D.; Pham, N.T.K.; Takada, K.; Komine-Aizawa, S.; Hayakawa, S. Myelin Oligodendrocyte Glycoprotein-Independent Rubella Infection of Keratinocytes and Resistance of First-Trimester Trophoblast Cells to Rubella Virus In Vitro. Viruses 2018, 10, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graham, C.H.; Hawley, T.S.; Hawley, R.G.; MacDougall, J.R.; Kerbel, R.S.; Khoo, N.; Lala, P.K. Establishment and characterization of first trimester human trophoblast cells with extended lifespan. Exp. Cell Res. 1993, 206, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Straszewski-Chavez, S.L.; Abrahams, V.M.; Alvero, A.B.; Aldo, P.B.; Ma, Y.; Guller, S.; Romero, R.; Mor, G. The isolation and characterization of a novel telomerase immortalized first trimester trophoblast cell line, Swan 71. Placenta 2009, 30, 939–948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hemphill, M.L.; Forng, R.Y.; Abernathy, E.S.; Frey, T.K. Time course of virus-specific macromolecular synthesis during rubella virus infection in Vero cells. Virology 1988, 162, 65–75. [Google Scholar] [CrossRef]
- Geyer, H.; Bauer, M.; Neumann, J.; Ludde, A.; Rennert, P.; Friedrich, N.; Claus, C.; Perelygina, L.; Mankertz, A. Gene expression profiling of rubella virus infected primary endothelial cells of fetal and adult origin. Virol. J. 2016, 13, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bilz, N.C.; Willscher, E.; Binder, H.; Bohnke, J.; Stanifer, M.L.; Hubner, D.; Boulant, S.; Liebert, U.G.; Claus, C. Teratogenic Rubella Virus Alters the Endodermal Differentiation Capacity of Human Induced Pluripotent Stem Cells. Cells 2019, 8, 870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reed, L.J.; Muench, H. A Simple Method of Estimating Fifty Percent Endpoints. Am. J. Hyg. 1938, 27, 493–497. [Google Scholar]
- Grigorov, B.; Rabilloud, J.; Lawrence, P.; Gerlier, D. Rapid titration of measles and other viruses: Optimization with determination of replication cycle length. PLoS ONE 2011, 6, e24135. [Google Scholar] [CrossRef] [PubMed]
- Pham, N.T.K.; Trinh, Q.D.; Takada, K.; Takano, C.; Sasano, M.; Okitsu, S.; Ushijima, H.; Komine-Aizawa, S.; Hayakawa, S. The Epithelial-to-Mesenchymal Transition-Like Process Induced by TGF-β1 Enhances Rubella Virus Binding and Infection in A549 Cells via the Smad Pathway. Microorganisms 2021, 9, 662. [Google Scholar] [CrossRef] [PubMed]
- Kaur, M.; Esau, L. Two-step protocol for preparing adherent cells for high-throughput flow cytometry. Biotechniques 2015, 59, 119–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perelygina, L.; Zheng, Q.; Metcalfe, M.; Icenogle, J. Persistent infection of human fetal endothelial cells with rubella virus. PLoS ONE 2013, 8, e73014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trinh, Q.D.; Takada, K.; Pham, N.T.K.; Takano, C.; Namiki, T.; Ikuta, R.; Hayashida, S.; Okitsu, S.; Ushijima, H.; Komine-Aizawa, S.; et al. Enhancement of rubella virus infection in immortalized human first-trimester trophoblasts under low glucose stress conditions. Front. Microbiol. 2022. Submitted. [Google Scholar]
- Ding, J.H.; Aldo, P.; Roberts, C.M.; Stabach, P.; Liu, H.; You, Y.; Qiu, X.M.; Jeong, J.; Maxwell, A.; Lindenbach, B.; et al. Placenta-derived interferon-stimulated gene 20 controls ZIKA virus infection. EMBO Rep. 2021, 22, e52450. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pham, N.T.K.; Trinh, Q.D.; Takada, K.; Komine-Aizawa, S.; Hayakawa, S. Low Susceptibility of Rubella Virus in First-Trimester Trophoblast Cell Lines. Viruses 2022, 14, 1169. https://doi.org/10.3390/v14061169
Pham NTK, Trinh QD, Takada K, Komine-Aizawa S, Hayakawa S. Low Susceptibility of Rubella Virus in First-Trimester Trophoblast Cell Lines. Viruses. 2022; 14(6):1169. https://doi.org/10.3390/v14061169
Chicago/Turabian StylePham, Ngan Thi Kim, Quang Duy Trinh, Kazuhide Takada, Shihoko Komine-Aizawa, and Satoshi Hayakawa. 2022. "Low Susceptibility of Rubella Virus in First-Trimester Trophoblast Cell Lines" Viruses 14, no. 6: 1169. https://doi.org/10.3390/v14061169
APA StylePham, N. T. K., Trinh, Q. D., Takada, K., Komine-Aizawa, S., & Hayakawa, S. (2022). Low Susceptibility of Rubella Virus in First-Trimester Trophoblast Cell Lines. Viruses, 14(6), 1169. https://doi.org/10.3390/v14061169