
A Novel Sandwich ELASA Based on Aptamer for Detection of Largemouth Bass Virus (LMBV)




Abstract
:1. Introduction
2. Methods and Materials
2.1. Cultured Cells, Fish and Viruses
2.2. Aptamers
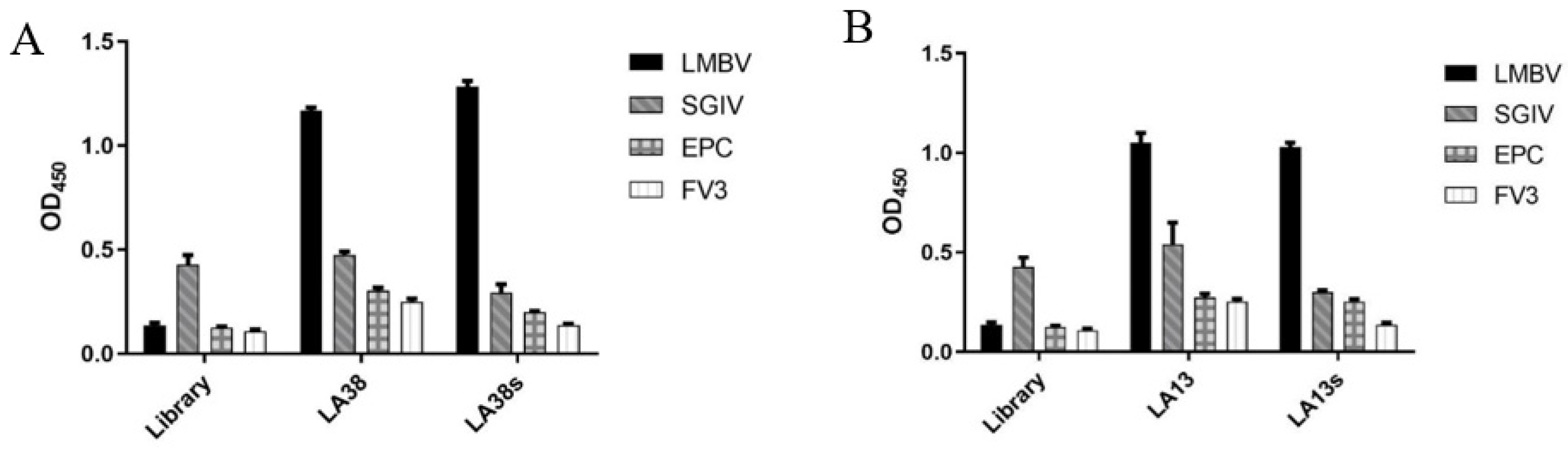
2.3. Specificity Analysis of LA38s and LA13s
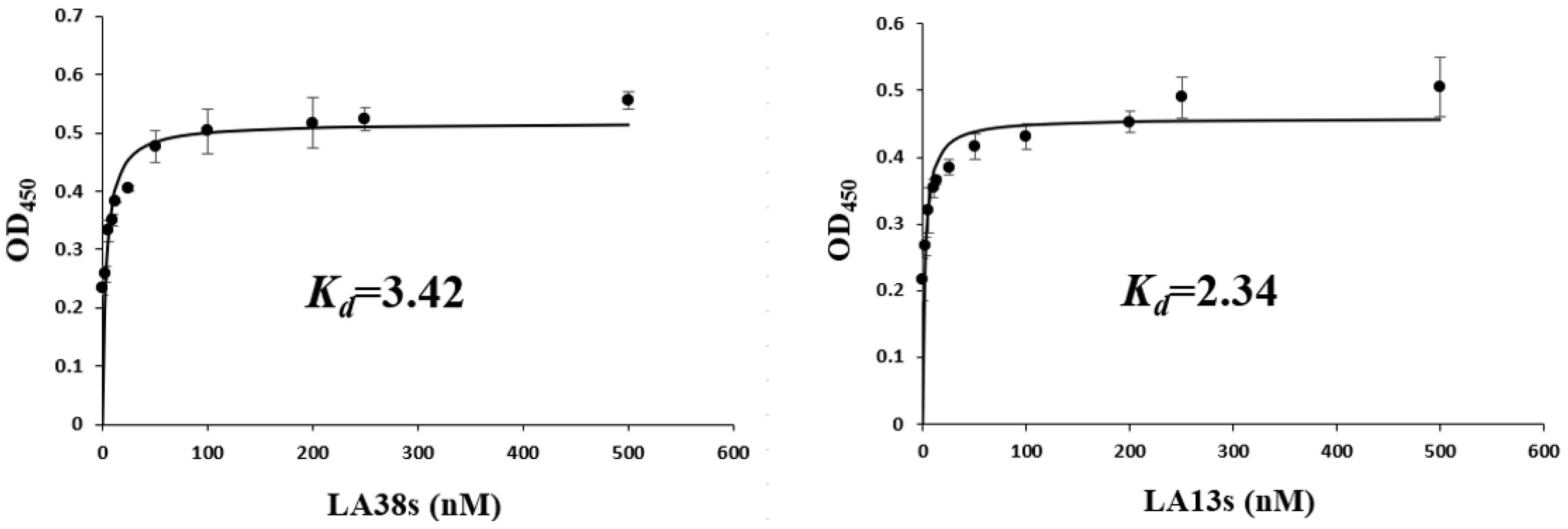
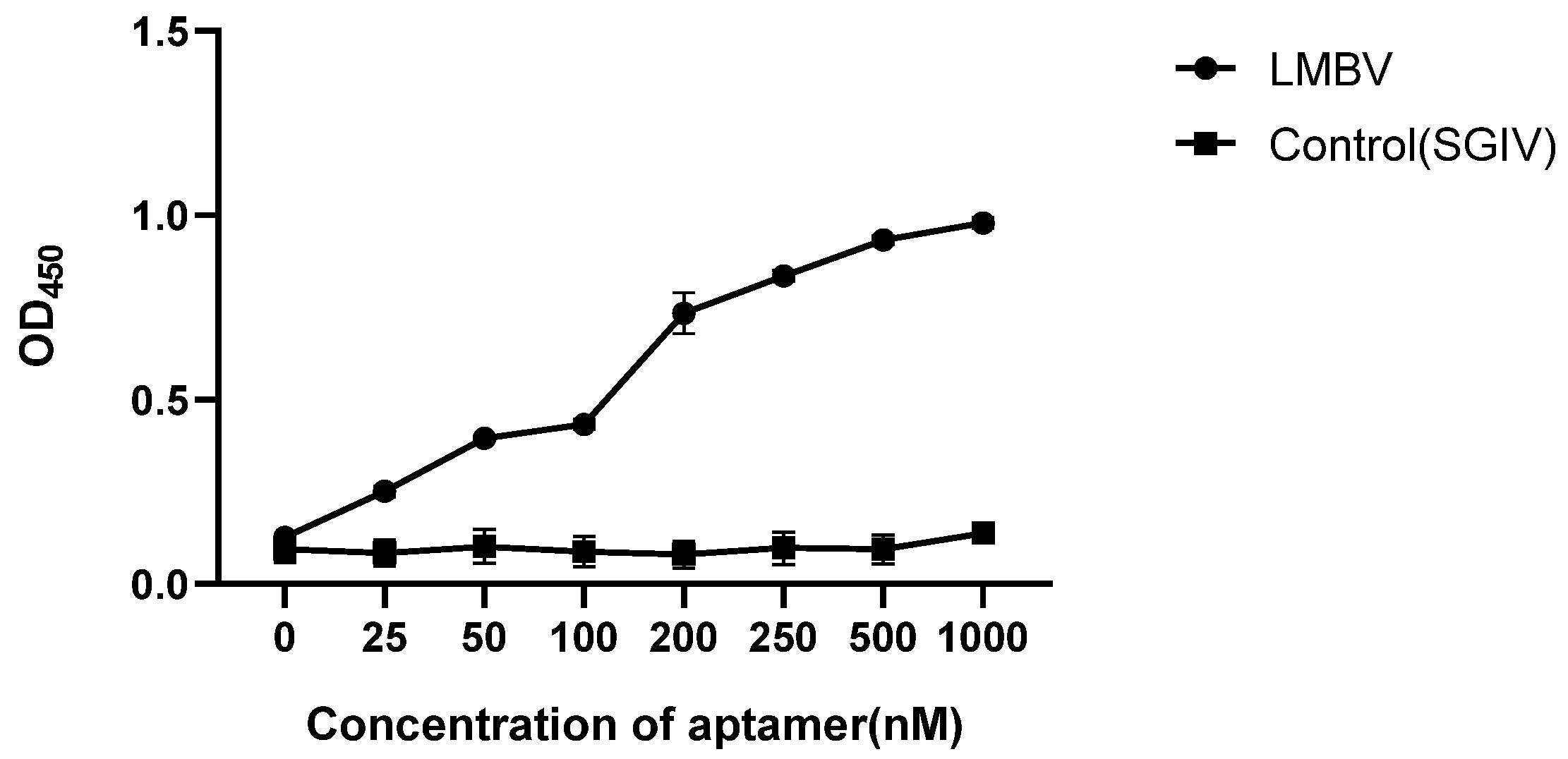
2.4. Affinity Analysis of LA38s and LA13s
2.5. Assembly of Sandwich ELASA
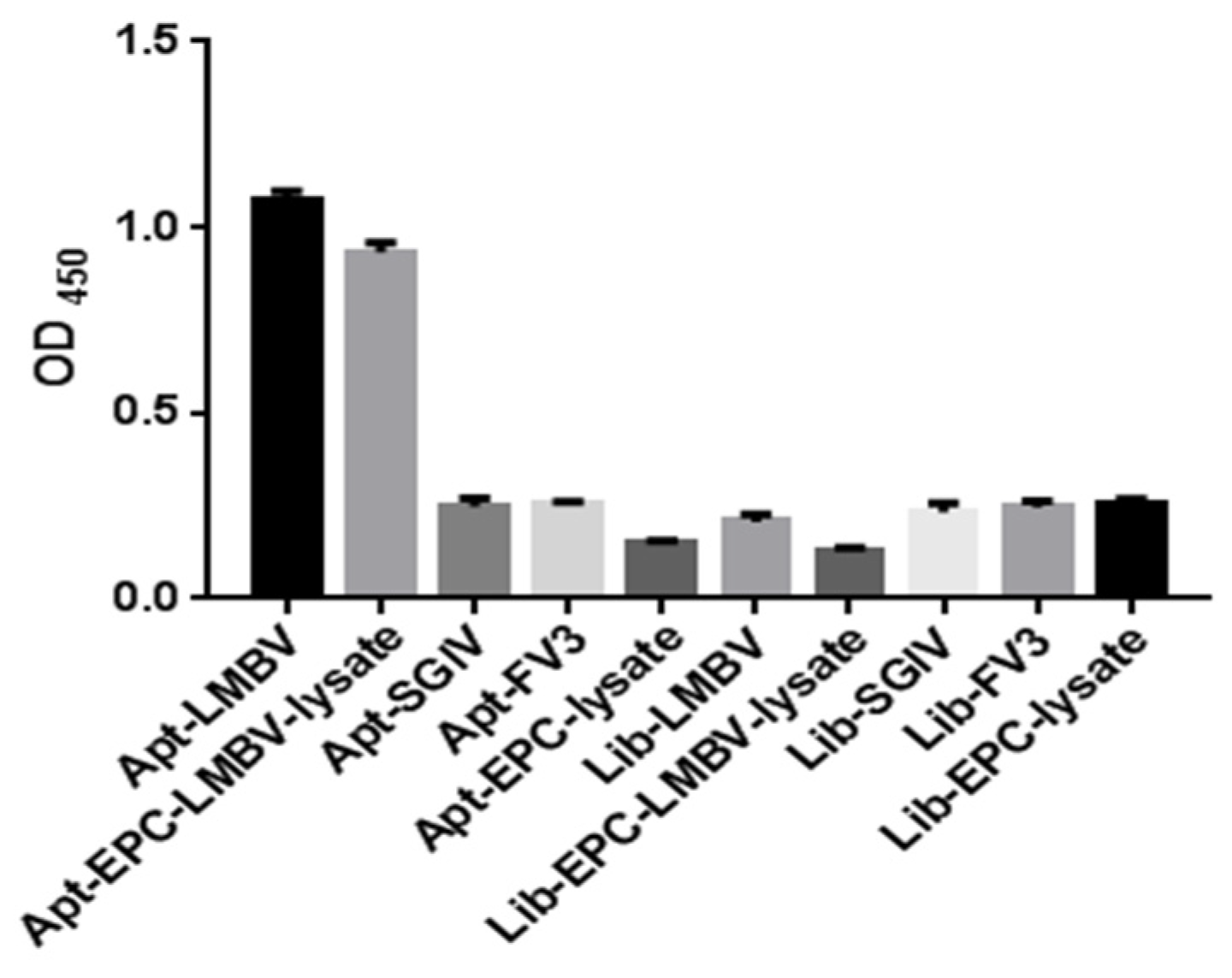
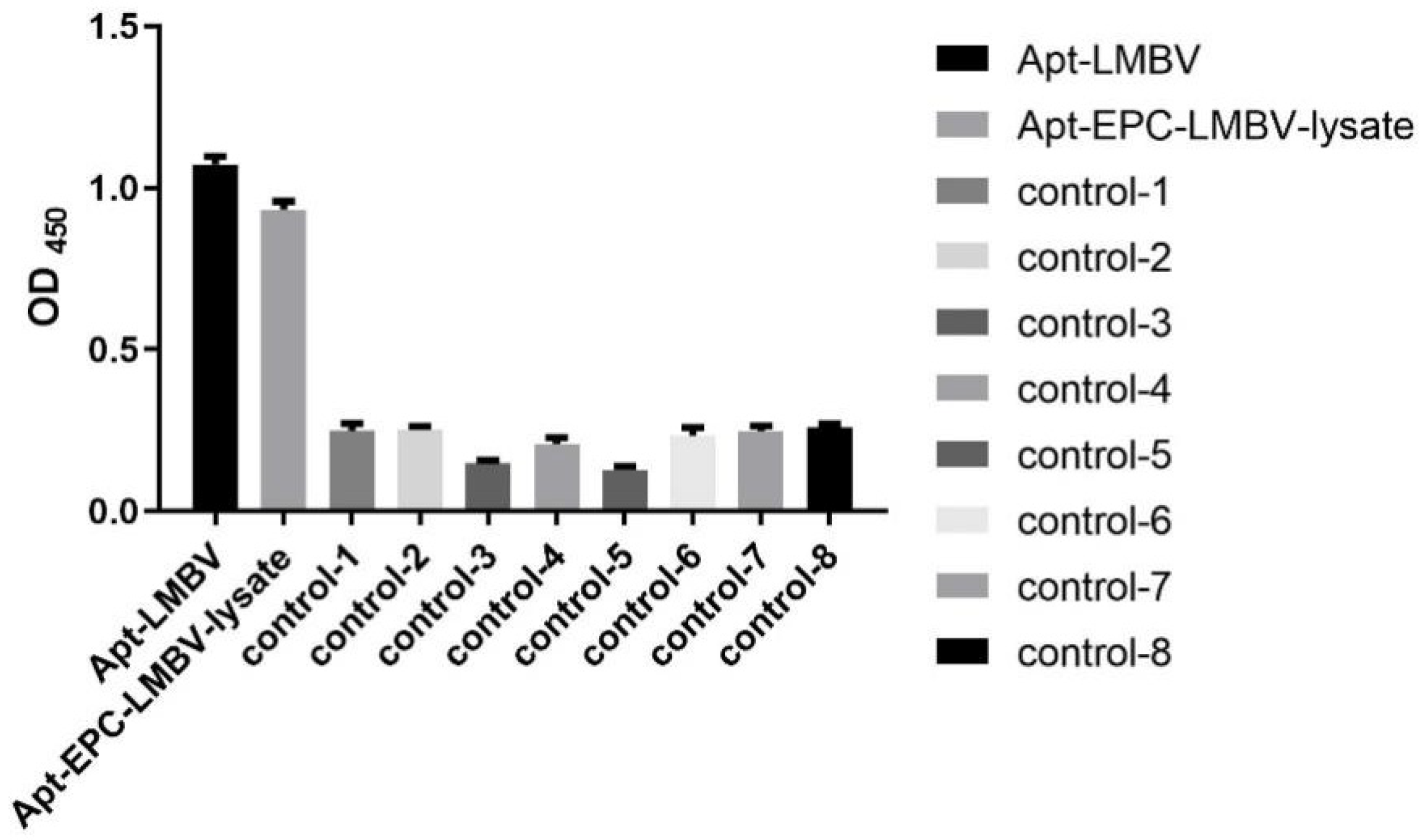
2.6. Specificity Analysis of Sandwich ELASA
2.7. The Optimum Working Concentration of Aptamer in Sandwich ELASA
2.8. Sensitivity Analysis of Sandwich ELASA
2.9. Detection of LMBV in Infected Fish Tissues by Sandwich ELASA
3. Results
3.1. Characterization of the Truncated Aptamers
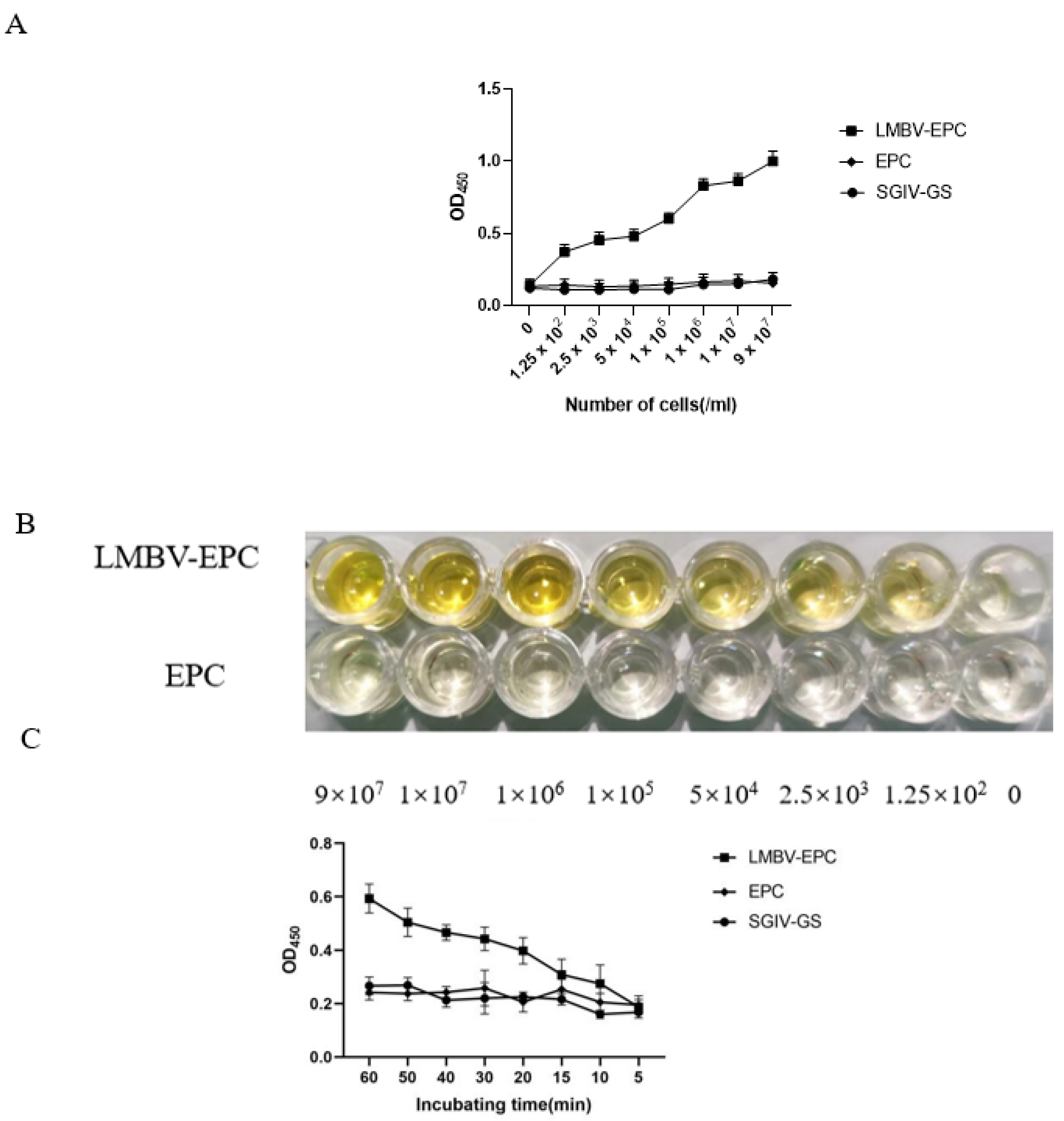
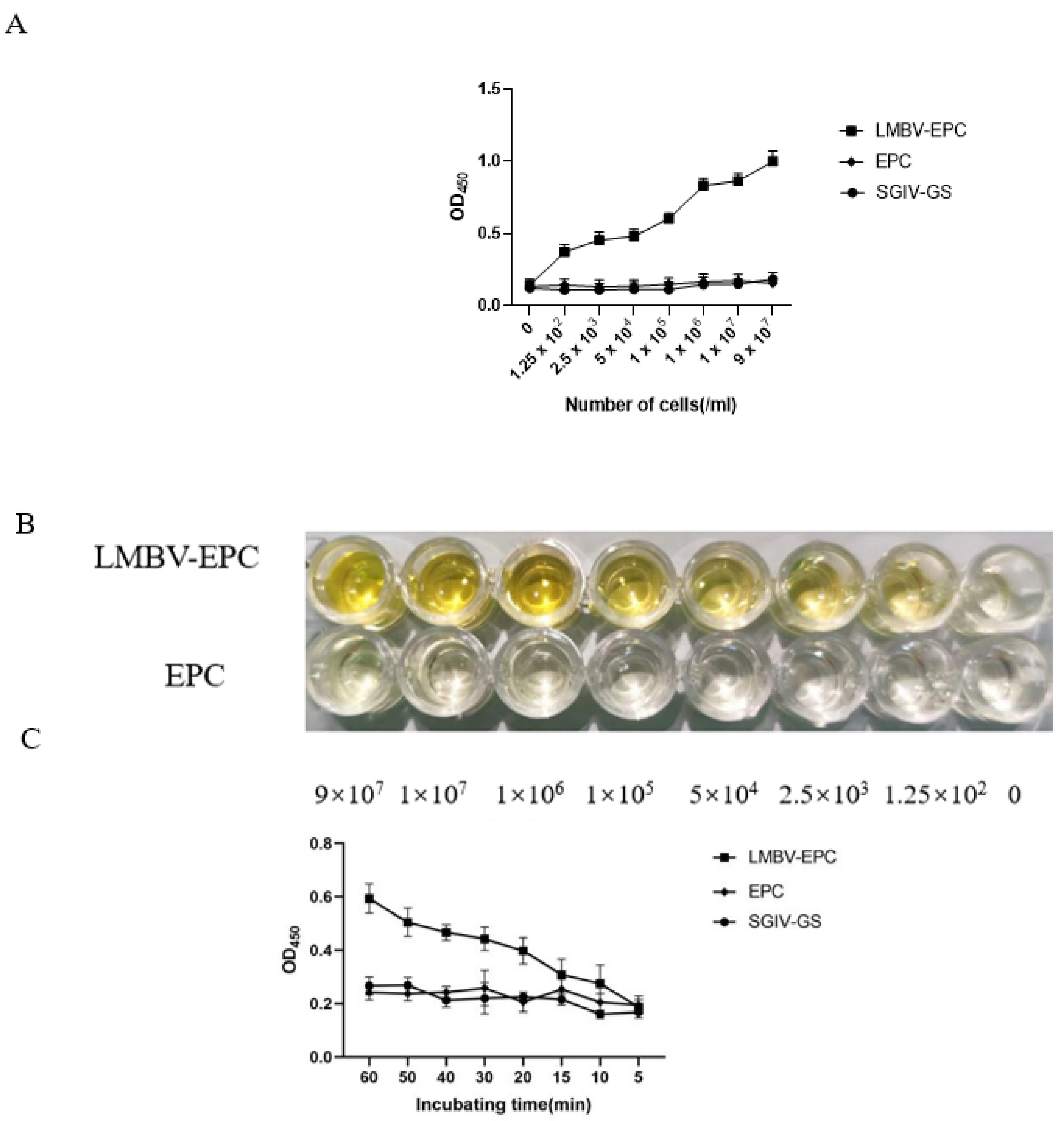
3.2. The Specificity of Sandwich ELASA
3.3. Optimum Working Concentration of Aptamers Used in the Sandwich ELASA
3.4. Sensitivity Analysis of the Sandwich ELASA
3.5. Detection of LMBV in Largemouth Bass by the Sandwich ELASA
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jia, Y.J.; Guo, Z.R.; Ma, R.; Qiu, D.K.; Wang, G.X.; Zhu, B. Protective immunity of largemouth bass immunized with immersed DNA vaccine against largemouth bass ulcerative syndrome virus. Fish Shellfish Immunol. 2020, 107, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.; Jiang, P.; Zhang, J.; Li, X.; Li, S.; Bai, J.; Fan, J.; Xu, P. High-Density Linkage Map and Mapping for Sex and Growth-Related Traits of Largemouth Bass (Micropterus salmoides). Front. Genet. 2019, 10, 960. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.L.; Cui, L.F.; Li, S.M.; Liu, X.Z.; Han, X.; Jiang, K.Y.; Yu, X.J.; Xu, L.J.; Wu, F.X.; Song, D.D.; et al. 2020 China Fishery Statistics Yearbook; China Agricultural Press: Beijing, China, 2020; p. 25. [Google Scholar]
- Huang, X.; Wang, W.; Huang, Y.; Xu, L.; Qin, Q. Involvement of the PI3K and ERK signaling pathways in largemouth bass virus-induced apoptosis and viral replication. Fish Shellfish Immunol. 2014, 41, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Wang, Y.; Feng, J. Rapid diagnosis of largemouth bass ranavirus in fish samples using the loop-mediated isothermal amplification method. Mol. Cell. Probes 2020, 52, 101569. [Google Scholar] [CrossRef]
- Plumb, J.A.; Zilberg, D. The lethal dose of largemouth bass virus in juvenile largemouth bass and the comparative susceptibility of striped bass. J. Aquat. Anim. Health 1999, 11, 246–252. [Google Scholar] [CrossRef]
- Zilberg, D.; Grizzle, J.M.; Plumb, J.A. Preliminary description of lesions in juvenile largemouth bass injected with largemouth bass virus. Dis. Aquat. Org. 2000, 39, 143–146. [Google Scholar] [CrossRef]
- Boonthai, T.; Loch, T.P.; Yamashita, C.J.; Smith, G.D.; Winters, A.D.; Kiupel, M.; Brenden, T.O.; Faisal, M. Laboratory investigation into the role of largemouth bass virus (Ranavirus, Iridoviridae) in smallmouth bass mortality events in Pennsylvania rivers. BMC Vet. Res. 2018, 14, 62. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Wang, S.; Wang, S.; Ren, T.; Gao, Z.; Gu, Q.; Zhang, W.; Liu, Y.H.; Wang, N.; Zhang, M.; et al. Development of multiplex real-time PCR assay for detection of ranaviruses. High Technol. Lett. 2015, 25, 746–752. [Google Scholar]
- Ma, D.; Bai, J.; Deng, G.; Li, S.; Ye, X.; Jiang, X. Sequence analysis of MCP gene from largemouth bass ulcerative syndrome virus and rapid detection by PCR assay. J. Fish. Sci. China 2010, 17, 1149–1156. [Google Scholar]
- Davydova, A.; Vorobjeva, M.; Pyshnyi, D.; Altman, S.; Vlassov, V.; Venyaminova, A. Aptamers against pathogenic microorganisms. Crit. Rev. Microbiol. 2016, 42, 847–865. [Google Scholar] [CrossRef] [Green Version]
- Keefe, A.D.; Pai, S.; Ellington, A. Aptamers as therapeutics. Nat. Rev. Drug Discov. 2010, 9, 537–550. [Google Scholar] [CrossRef] [PubMed]
- Ellington, A.D.; Szostak, J.W. In vitro selection of RNA molecules that bind specific ligands. Nature 1990, 346, 818–822. [Google Scholar] [CrossRef] [PubMed]
- Song, K.M.; Lee, S.; Ban, C. Aptamers and their biological applications. Sensors 2012, 12, 612–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, M.H.; Elsherbiny, M.E.; Emara, M. Updates on Aptamer Research. Int. J. Mol. Sci. 2019, 20, 2511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, X.; Wu, J.; Gu, J. Application of Aptamers in Virus Detection and Antiviral Therapy. Front. Microbiol. 2019, 10, 1462. [Google Scholar] [CrossRef] [Green Version]
- Zhu, G.; Ye, M.; Donovan, M.J.; Song, E.; Zhao, Z.; Tan, W. Nucleic acid aptamers: An emerging frontier in cancer therapy. Chem. Commun. 2012, 48, 10472–10480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tombelli, S.; Minunni, M.; Mascini, M. Analytical applications of aptamers. Biosens. Bioelectron. 2005, 20, 2424–2434. [Google Scholar] [CrossRef]
- Van Dorst, B.; Mehta, J.; Bekaert, K.; Rouah-Martin, E.; De Coen, W.; Dubruel, P.; Blust, R.; Robbens, J. Recent advances in recognition elements of food and environmental biosensors: A review. Biosens. Bioelectron. 2010, 26, 1178–1194. [Google Scholar] [CrossRef]
- Qin, Q.; Chang, S.; Ngoh, G.; Gibson-Kueh, S.; Shi, C.; Lam, T.J. Characterization of a novel ranavirus isolated from grouper, Epinephelus tauvina. Dis. Aquat. Org. 2003, 53, 1–9. [Google Scholar] [CrossRef]
- Qin, Q.; Wu, T.; Jia, T.; Hegde, A.; Zhang, R. Development and characterization of a new tropical marine fish cell line from grouper, Epinephelus coioides susceptible to iridovirus and nodavirus. J. Virol. Methods 2006, 131, 58–64. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Wang, L.Q.; Liu, J.; Zhang, Z.; Zhou, L.; Huang, X.; Wei, J.; Yang, M.; Qin, Q.; Wang, S. Generation and identification of novel DNA aptamers with antiviral activities against largemouth bass virus (LMBV). Aquaculture 2021, 547, 737478. [Google Scholar] [CrossRef]
- Liang, H.R.; Hu, G.Q.; Zhang, T.; Yang, Y.J.; Zhao, L.L.; Qi, Y.L.; Wang, H.L.; Gao, Y.W.; Yang, S.T.; Xia, X.Z. Isolation of ssDNA aptamers that inhibit rabies virus. Int. Immunopharmacol. 2012, 14, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Zhou, L.; Wei, J.; Yu, Y.; Yang, M.; Wei, S.; Qin, Q. Development and characterization of aptamer-based enzyme-linked apta-sorbent assay for the detection of Singapore grouper iridovirus infection. J. Appl. Microbiol. 2016, 121, 634–643. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Li, P.; Ni, S.; Yu, Y.; Yang, M.; Wei, S.; Qin, Q. Rapid and sensitive detection of redspotted grouper nervous necrosis virus (RGNNV) infection by aptamer–coat protein–aptamer sandwich enzyme-linked apta-sorbent assay (ELASA). J. Fish Dis. 2017, 40, 1831–1838. [Google Scholar] [CrossRef]
- Plumb, J.A.; Grizzle, J.M.; Young, H.E.; Noyes, A.D.; Lamprecht, S. An iridovirus isolated from wild largemouth bass. J. Aquat. Anim. Health 1996, 8, 265–270. [Google Scholar] [CrossRef]
- Beck, B.H.; Bakal, R.S.; Brunner, C.J.; Grizzle, J.M. Virus distribution and signs of disease after immersion exposure to largemouth bass virus. J. Aquat. Anim. Health 2006, 18, 176–183. [Google Scholar] [CrossRef]
- Grizzle, J.M.; Brunner, C.J. Review of largemouth bass virus. Fisheries 2003, 28, 10–14. [Google Scholar] [CrossRef]
- Ho, P.Y.; Chen, Y.C.; Maekawa, S.; Hu, H.H.; Tsai, A.W.; Chang, Y.F.; Wang, P.C.; Chen, S.C. Efficacy of recombinant protein vaccines for protection against Nocardia seriolae infection in the largemouth bass Micropterus salmoides. Fish Shellfish Immunol. 2018, 78, 35–41. [Google Scholar] [CrossRef]
- Ma, D.M.; Deng, G.C.; Bai, J.J.; Jiang, X.Y.; Cao, T.T.; Cai, L. Prokaryotic expression of MCP gene from largemouth bass ulcerative syndrome virus and the immune effect analysis of recombinant protein. Biotechnol. Bull. 2016, 32, 139–144. [Google Scholar]
- Byadgi, O.; Chen, C.W.; Wang, P.C.; Tsai, M.A.; Chen, S.C. De novo transcriptome analysis of differential functional gene expression in largemouth bass (Micropterus salmoides) after challenge with nocardia seriolae. Int. J. Mol. Sci. 2016, 17, 1315. [Google Scholar] [CrossRef] [Green Version]
- Balamurugan, S.; Obubuafo, A.; Soper, S.A.; Spivak, D.A. Surface immobilization methods for aptamer diagnostic applications. Anal. Bioanal. Chem. 2008, 390, 1009–1021. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, C.; Papamichael, K.; Guilbault, G.; Schwarzacher, T.; Gariepy, J.; Missailidis, S. DNA aptamers against the MUC1 tumour marker: Design of aptamer–antibody sandwich ELISA for the early diagnosis of epithelial tumours. Anal. Bioanal. Chem. 2008, 390, 1039–1050. [Google Scholar] [CrossRef] [PubMed]
- Toh, S.Y.; Citartan, M.; Gopinath, S.; Tang, T. Aptamers as a replacement for antibodies in enzyme-linked immunosorbent assay. Biosens. Bioelectron. 2015, 64, 392–403. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′–3′) |
|---|---|
| LMBV-MCP-qRT-PCR-F | TCGCCACTTATGACAGCCTTGA |
| LMBV-MCP-qRT-PCR-R | GACCTGGGCACTCCTACGGA |
| Actin-qRT-PCR-F | TACGAGCTGCCTGACGGACA |
| Actin-qRT-PCR-R | GGCTGTGATCTCCTTCTGCA |
| Tested Samples | Positive Samples | Positive Ratios (%) | ||
|---|---|---|---|---|
| ELASA | qRT-PCR | ELASA | qRT-PCR | |
| 30 | 14 | 18 | 46.7 | 60 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Zhang, Z.; Li, J.; Huang, X.; Wei, J.; Yang, J.; Guan, L.; Wen, X.; Wang, S.; Qin, Q. A Novel Sandwich ELASA Based on Aptamer for Detection of Largemouth Bass Virus (LMBV). Viruses 2022, 14, 945. https://doi.org/10.3390/v14050945
Zhang X, Zhang Z, Li J, Huang X, Wei J, Yang J, Guan L, Wen X, Wang S, Qin Q. A Novel Sandwich ELASA Based on Aptamer for Detection of Largemouth Bass Virus (LMBV). Viruses. 2022; 14(5):945. https://doi.org/10.3390/v14050945
Chicago/Turabian StyleZhang, Xinyue, Zemiao Zhang, Junrong Li, Xiaohong Huang, Jingguang Wei, Jiahui Yang, Lingfeng Guan, Xiaozhi Wen, Shaowen Wang, and Qiwei Qin. 2022. "A Novel Sandwich ELASA Based on Aptamer for Detection of Largemouth Bass Virus (LMBV)" Viruses 14, no. 5: 945. https://doi.org/10.3390/v14050945
APA StyleZhang, X., Zhang, Z., Li, J., Huang, X., Wei, J., Yang, J., Guan, L., Wen, X., Wang, S., & Qin, Q. (2022). A Novel Sandwich ELASA Based on Aptamer for Detection of Largemouth Bass Virus (LMBV). Viruses, 14(5), 945. https://doi.org/10.3390/v14050945