Abstract
Metagenomic approaches used for virus diagnostics allow for rapid and accurate detection of all viral pathogens in the plants. In order to investigate the occurrence of viruses and virus-like organisms infecting grapevine from the Ampelographic collection Kromberk in Slovenia, we used Ion Torrent small RNA sequencing (sRNA-seq) and the VirusDetect pipeline to analyze the sRNA-seq data. The used method revealed the presence of: Grapevine leafroll-associated virus 1 (GLRaV-1), Grapevine leafroll-associated virus 2 (GLRaV-2), Grapevine leafroll-associated virus 3 (GLRaV-3), Grapevine rupestris stem pitting-associated virus (GRSPaV), Grapevine fanleaf virus (GFLV) and its satellite RNA (satGFLV), Grapevine fleck virus (GFkV), Grapevine rupestris vein feathering virus (GRVFV), Grapevine Pinot gris virus (GPGV), Grapevine satellite virus (GV-Sat), Hop stunt viroid (HSVd), and Grapevine yellow speckle viroid 1 (GYSVd-1). Multiplex reverse transcription-polymerase chain reaction (mRT-PCR) was developed for validation of sRNA-seq predicted infections, including various combinations of viruses or viroids and satellite RNA. mRT-PCR could further be used for rapid and cost-effective routine molecular diagnosis, including widespread, emerging, and seemingly rare viruses, as well as viroids which testing is usually overlooked.
1. Introduction
Grapevine is one of the most susceptible plants to viral infections. More than 86 viruses belonging to different families and genera have been reported to infect grapevine [1], and their number is constantly growing. Recently, two novel members of the genus Vitivirus have been identified in South Africa [2].
Most grapevine viruses have an RNA genome, including viruses associated with four major and widespread disease complexes (infectious degeneration and decline, leafroll, rugose wood, and fleck disease complex) [3]. Viruses with a DNA genome have also been identified in grapevine, and they are associated with vein-clearing and vine decline syndrome [4], red blotch disease [5,6], roditis leaf discoloration [7], and fruit tree decline syndrome [8].
Viral pathogens are spread over long distances by infected material (nursery productions), whereas infections within a vineyard or an area are transmitted mechanically and by insects, mites, or nematodes [3]. Viruses and virus-like organisms can cause severe developmental and morphological malformations, affect grapevine physiological activity and metabolism, reduce yield, decrease quality of grapes and wines, and shorten vineyard life, resulting in high economic losses [9,10,11,12]. For example, estimated economic losses caused by Grapevine leafroll-associated virus 3 (GLRaV-3) in California are more than USD 90 million annually [13]. Therefore, rapid, effective, and reliable detection is crucial to limit their spread.
High-throughput sequencing technology (HTS), which targets all nucleic acid types, enables rapid and accurate detection, including previously described and novel viruses and virus-like organisms [14,15,16,17]. An approach that enables virus discovery through HTS technology and assembly of small RNAs (small RNA sequencing, sRNA-seq) has proven to be highly efficient in the detection of new RNA and DNA grapevine viruses [4,7,18,19], virome studies [20,21,22,23], and to evaluate the efficacy of different elimination methods such as chemotherapy, somatic embryogenesis, and meristem tissue culture [24,25]. All in silico predicted grapevine viral infections are most commonly validated using RT-PCR [21,22,23,24,25,26]. Several other molecular diagnostic methods as well as immunological detection methods, and biological indexing are used in plant virology [27]. However, most routine diagnostic assays can only be used for detection of one target virus/virus-like organism. Multiplex RT-PCR (mRT-PCR)/multiplex PCR (mPCR), which enables simultaneous amplification of several viral entities in a single reaction, is less labor intensive, time saving and cost-effective, especially when a large number of samples needs to be tested for mixed infections. To date, mRT-PCR/mPCR has been used to detect various herbaceous and woody plant-infecting viruses and viroids [28,29,30,31,32,33,34,35,36,37,38,39,40,41,42], including those infecting grapevine [43,44,45,46,47,48,49]. However, mRT-PCR/mPCR has not been used for validation of HTS-predicted viral infections in grapevines thus far.
The aim of the presented work was to perform sRNA-seq for the diagnosis of grapevine viral pathogens in six grapevine varieties from the Ampelographic collection Kromberk, Slovenia, and to develop an mRT-PCR assay for the validation of sRNA-seq data that could be further used for rapid and cost-effective routine molecular diagnosis in large-scale surveys.
2. Materials and Methods
2.1. Plant Material
A total of 13 cuttings from six grapevine varieties, two red, ‘Cipro’ (‘Rosenmuscateller’) and ‘Pokalca’ (‘Schioppettino’), and four white, ‘Malvazija’ (‘Malvasia d’Istria’), ‘Volovnik’ (‘Vela pergola’), ‘Rebula’ (‘Ribolla gialla’), and ‘Poljšakica’, were collected from the Ampelographic collection Kromberk near Nova Gorica, Slovenia, in 2017 (Figure 1). Cuttings were sprouted in water at room temperature (21 °C) at the Biotechnical Faculty, University of Ljubljana. Developed young leaves were sampled and stored at –80 °C for further analysis.

Figure 1.
(a) Location of the Ampelographic collection Kromberk 45°57′40.8″ N 13°39′44.7″ E; (b) redding of the interveinal areas caused by GLRaV-3 on the ‘Pokalca’ variety; (c) shoot malformation (shorten internodes) caused by GFLV on the ‘Rebula’ variety.
2.2. Small RNA Isolation, Library Construction, sRNA-Seq and Bioinformatics Analysis
The selected samples were pooled together into four pools representing either samples of the same variety (L1, L2, and L3) or of different varieties (L4). Small RNAs (sRNAs) were isolated using mirVana™ miRNA Isolation Kit (Ambion, Life Technologies, Waltham, MA, USA) according to the manufacturer’s instructions for the enrichment of sRNAs. The quantity and quality of sRNAs were assessed using the Agilent 2100 Bioanalyzer (Agilent Technologies, Inc., Santa Clara, CA, USA) according to the manufacturer’s instructions. Libraries of sRNAs were constructed using the Ion Total RNA-Seq Kit v2 (Ion Torrent™, Waltham, MA, USA) and were barcoded using the Xpress™ RNA-Seq Barcode 1–16 Kit (Ion Torrent™, Waltham, MA, USA) according to the manufacturer’s instructions. The yield and size distribution of the amplified cDNA libraries were determined using the Agilent 2100 Bioanalyzer (Agilent Technologies, Inc., Santa Clara, CA, USA). Libraries were pooled at equimolar concentrations and prepared for sequencing using the Ion PI™ Hi-Q™ OT2 200 Kit and Ion PI™ Hi-Q™ Sequencing 200 Kit (Ion Torrent™, Waltham, MA, USA) according to the manufacturer’s instructions. Sequencing was performed on Ion PI™ chips v3 using an Ion Proton™ System (Ion Torrent™, Waltham, MA, USA), according to the manufacturer’s instructions. Raw sequencing data were deposited in the National Center for Biotechnology Information (NCBI) Sequence Read Archive (SRA) database under BioProject number PRJNA667593, BioSamples: SAMN16378719-SAMN16378722. The sRNA-seq data were analyzed using the VirusDetect pipeline with default parameters [50]. The pipeline performs reference-guided assembly using the Burrows–Wheeler Aligner (BWA) and de novo assembly using the Velvet Genomic Assembler. The plant virus database was used as reference, and the grapevine genome was selected to subtract host sRNAs.
2.3. mRT-PCR for Validation of sRNA-Seq Predicted Viral Pathogens
Confirmation of sRNA-seq-predicted infections was performed by mRT-PCR. Total RNA was extracted from 100 mg of frozen leaves using the RNeasy Plant Mini Kit (Qiagen, Hilden, Germany). First strand cDNA synthesis was performed using the High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems™, Foster City, CA, USA) according to the manufacturer’s instructions. mRT-PCR was performed using the KAPA2G Fast Multiplex PCR Kit (KAPA Biosystems, Wilmington, MA, USA). The reaction mixture was prepared using 12.5 µL of KAPA2G Fast Multiplex Mix (KAPA2G Fast HotStart DNA Polymerase, KAPA2G Buffer A, 0.2 mM of each dNTP, 3 mM MgCl2, and stabilizers), 0.2 µL (0.1 µL for GRVFV) of each 10 µM forward and reverse primer (final concentration 0.08 µM; for GRVFV 0.04 µM), 1 µL of pooled cDNA, and nuclease-free water up to 25 µL. Primers are listed in Table 1. Amplification was performed in a thermal cycler (Applied Biosystems™, Waltham, MA, USA) under the following conditions: initial denaturation at 95 °C for 3 min, 35 cycles consisting of a denaturation step at 95 °C for 15 s, annealing at 58 °C for 30 s, extension at 72 °C for 1 min, and a final extension at 72 °C for 1 min. The amplified products were analyzed by electrophoresis on 1.2% agarose gel, stained with ethidium bromide, and visualized under a UV transilluminator. Amplicons sizes were determined by comparison with the GeneRuler™ 100 bp Plus DNA Ladder (Thermo Fisher Scientific, Waltham, MA, USA).

Table 1.
List of primers used for mRT-PCR detection.
Table 1.
List of primers used for mRT-PCR detection.
| Viral Pathogen | Primer Name | Primer Sequence (5′-3′) | Product Size * | Tm * | GC % * | Amplified Region | Reference |
|---|---|---|---|---|---|---|---|
| GLRaV-3 | LR3-8504V | ATGGCATTTGAACTGAAATT | 942 bp | 51.81 | 30.00 | CP | [51] |
| LR3-9445C | CTACTTCTTTTGCAATAGTT | 48.91 | 30.00 | ||||
| GLRaV-2 | LRaV-2 (1) | AGGCGGATCGACGAATAC | 821 bp | 56.64 | 55.56 | hsp70-like protein, p63 | [52] |
| LRaV-2 (2) | ATCCTGTCCGGCGCTGTG | 62.46 | 66.67 | ||||
| GPGV | Pg-Mer-F1 | GGAGTTGCCTTCGTTTACGA | 770 bp | 58.21 | 50.00 | MP/CP | [53] |
| Pg-Mer-R1 | GTACTTGATTCGCCTCGCTCA | 60.47 | 52.38 | ||||
| GRVFV | GRVFV_6090F | CATCGTTCTGATCCTCAGCC | 516 bp | 58.14 | 55.00 | polyprotein | [54] |
| GRVFV_6605R | AGAGACGCTGACCATGCCAC | 62.51 | 60.00 | ||||
| GFLV | GFLV_13_16_F | TGACACGTGCCTTTATTGGA | 488 bp | 57.45 | 45.00 | polyprotein, segment RNA2 | [23] |
| GFLV_13_16_R | CTCAAGTTGGGGAAGGTCAA | 57.34 | 50.00 | ||||
| GLRaV-1 | CPd2/F | GTTACGGCCCTTTGTTTATTATGG | 398 bp | 58.42 | 41.67 | CPd2 | [55] |
| CPd2/R | CGACCCCTTTATTGTTTGAGTATG | 57.88 | 41.67 | ||||
| GRSPaV | RSP 48 | AGCTGGGATTATAAGGGAGGT | 330 bp | 57.63 | 47.62 | CP | [56] |
| RSP 49 | CCAGCCGTTCCACCACTAAT | 60.04 | 55.00 | ||||
| GV-Sat | GV-Sat_for | CCCGGACTCACATTAAGTCAA | 305 bp | 57.67 | 47.62 | ORF1, ORF2, 3′UTR | [57] |
| GV-Sat_rev | GCACAAGCGAGATAACAGCA | 58.92 | 50.00 | ||||
| GFkV | GFkVf | TGACCAGCCTGCTGTCTCTA | 179 bp | 60.25 | 55.00 | CP | [44] |
| GFkVr | TGGACAGGGAGGTGTAGGAG | 59.96 | 60.00 | ||||
| satGFLV | FP3-F | GTGGSCCCGCRAGTGT | 870 bp | degenerative primer pair | hypothetical protein | [58] | |
| RP-R | TAAWGAGCAACCAAAATCCCA | ||||||
| HSVd | HSV-78P | AACCCGGGGCAACTCTTCTC | ~300 bp | 62.13 | 60.00 | complete genome | [59] |
| HSV-83M | AACCCGGGGCTCCTTTCTCA | 63.34 | 60.00 | ||||
| GYSVd-1 | - | TGTGGTTCCTGTGGTTTCAC | ~368 bp | 58.24 | 50.00 | complete genome | [60] |
| - | ACCACAAGCAAGAAGATCCG | 58.19 | 50.00 | ||||
* Determined with Primer-BLAST.
3. Results
3.1. Viruses and Virus-like Organisms Detected by sRNA-Seq
sRNA-seq of pooled grapevine samples resulted in 17,195,263–18,713,942 reads per pool (Table 2). Of the total reads, 50.12–67.22% were mapped to the grapevine genome, while 3.06–11.98% were mapped to viral genomes (Table 2). After concatenating unique reference-guided contigs and unique de novo assembled contigs and after removing redundancies, 461–1102 unique viral contigs were generated (Table 2). The total number of reference viral sequences identified by BLASTN search per library is presented in Table 2, while for each viral pathogen, it is presented in Table S1.
Table 2.
Summary of results obtained with VirusDetect.
Out of identified references, we selected one complete genome sequence per viral pathogen in each library that had the highest coverage. The used method revealed the presence of: Grapevine leafroll-associated virus 1 (GLRaV-1), Grapevine leafroll-associated virus 2 (GLRaV-2), Grapevine leafroll-associated virus 3 (GLRaV-3), Grapevine rupestris stem pitting-associated virus (GRSPaV), Grapevine fanleaf virus (GFLV) and its satellite RNA (satGFLV), Grapevine fleck virus (GFkV), Grapevine rupestris vein feathering virus (GRVFV), Grapevine Pinot gris virus (GPGV), Grapevine satellite virus (GV-Sat), Hop stunt viroid (HSVd) and Grapevine yellow speckle viroid 1 (GYSVd-1). The highest number of viral entities (nine) was found in the library that was a mixture of three different varieties (L4). Eight viral pathogens were detected in the library of variety ‘Cipro’ (L1), while in the other two libraries (L2 and L3), the number of identified viral pathogens was seven (Table 3). The coverage with references from the database was between 61.99% (GRVFV, L1) and 99.96% (GLRaV-3, L2) (Table 3; Figure 2), with a sequencing depth between 7X (GRSPaV, L1) and 5313.2 X (satGFLV, L2) (Table 3). GLRaV-1, GLRaV-2, and GV-Sat were present only in one library (L1). GFkV was detected in L3 and L4, and GFLV and satGFLV were detected in L2 and L4. GFLV possesses a bipartite genome; thus, the sRNA-seq data for RNA1 and RNA2 are shown in Table 3. GLRaV-3 was detected in three libraries (L2, L3, and L4) and had the highest coverage (99.80–99.96%) among viruses in all three libraries. GRSPaV, GPGV, and GRVFV were detected in all libraries. GRSPaV had low sequencing depth in all libraries (7X, 8.9X, 10.4X, and 9.7X, respectively). GRVFV had the lowest references coverage in all libraries (61.99%, 70.24%, 68.47%, and 70.21%, respectively). Considering viroids, HSVd was detected in all libraries, while GYSVd-1 was absent only in L2 (Table 3).
Table 3.
Viruses and virus-like organisms detected with BLASTN search (VirusDetect pipeline).
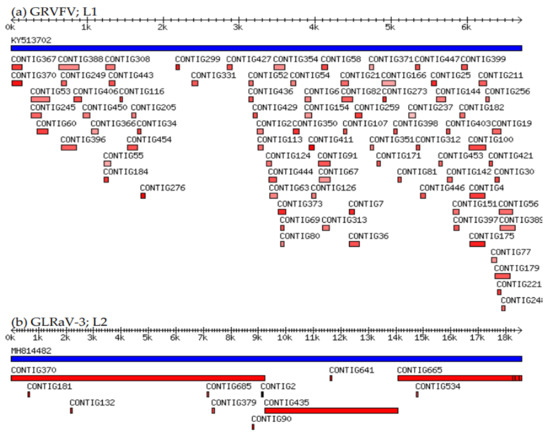
Figure 2.
Virus-assembled contigs (red bars) mapped to complete reference genome sequence (blue bars): (a) GRVFV, virus with the lowest reference genome coverage (61.99%; L1); (b) GLRaV-3, virus with the highest reference genome coverage (99.96%; L2).
3.2. mRT-PCR for Validation of sRNA-Seq Predicted Viral Pathogens
Primer combinations with different expected amplified fragments were chosen for mRT-PCR to allow for differentiation on the agarose gel. All primers corresponded to those found in the literature (Table 1). The primers for GV-Sat and GFLV had been designed in our previous studies [23,57]. Several parameters such as primer concentration (0.04–0.2 µM), annealing temperature (55–60 °C), number of cycles (30–35), and amount of cDNA (1 µL and 2 µL) were optimized to determine the best conditions for simultaneous amplification of the predicted infections. As under-amplified amplicons were obtained with a higher primers concentration (0.2 µM), it was reduced to 0.08 µM. With this primers concentration (0.08 µM) and an annealing temperature of 55 °C, all predicted viruses were amplified in all libraries, although nonspecific banding patterns of approximately 250 bp were also observed. In an effort to reduce these background bands, the annealing temperature was increased to 58 °C, and the concentration of the primer pair (GRVFV_6090F/GRVFV_6605R) amplifying 516 bp of GRVFV polyprotein product (Table 1) was decreased to 0.04 µM. Better results were obtained with a lower amount of cDNA (1 µL), compared with 2 µL (data not shown). Under these conditions (primer concentration 0.08 µM and 0.04 µM for GRVFV, annealing temperature 58 °C, 35 cycles and 1 µL of cDNA), specific RT-PCR amplification products of the expected sizes were obtained for all viral pathogens in all libraries. Different combinations of viruses were amplified simultaneously in all four libraries: L1 (GV-Sat, GRSPaV, GLRaV-1, GRVFV, GPGV, and GLRaV-2); L2 (GRSPaV, GFLV, GRVFV, GPGV, GLRaV-3); L3 (GFkV, GRSPaV, GRVFV, GPGV, GLRaV-3); L4 (GFkV, GRSPaV, GFLV, GRVFV, GPGV, GLRaV-3) (Figure 3). In addition, different combinations of viroids/satGFLV were amplified simultaneously: L1 and L3 (HSVd, GYSVd-1), L2 (HSVd, satGFLV), L4 (HSVd, GYSVd-1, satGFLV) (Figure 3).
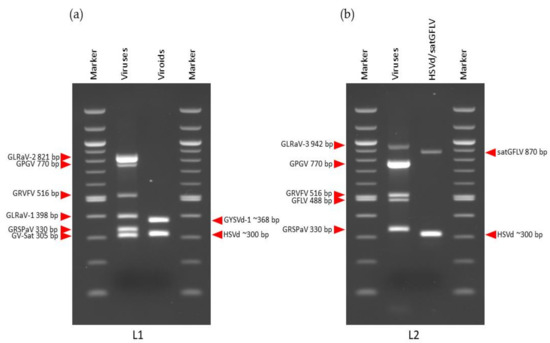
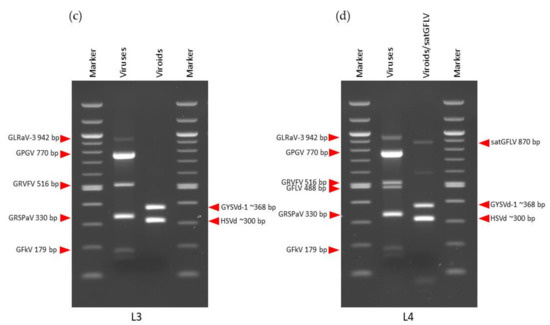
Figure 3.
Validation of sRNA-seq-predicted viruses and virus-like organisms with mRT-PCR: (a) L1; (b) L2; (c) L3; (d) L4.
4. Discussion
Thirteen grapevines of six important autochthonous and local varieties were screened for viruses and virus-like organisms with sRNA-seq. A total of 70,902,637 reads were generated, and 5,272,381 (7.44%) were mapped to viral reference sequences, while 42,342,716 (59.72%) originated from grapevine. The BLASTN search of the unique viral-assembled contigs revealed the presence of widespread viruses associated with four major disease complexes, emerging virus, GV-Sat (first report in Slovenia) [57], as well as worldwide-distributed viroids. A high number of contigs and their short length were observed for GRSPaV, GFLV, GRVFV and GFkV, which is in accordance with our previous study [23], and may be related with their high genetic variability. For example, GFLV (RNA1) reference sequence (JX513889), which is 7340 nt long, was covered with 127 contigs (Figure S1). In contrast, GLRaV-2 reference sequence (FJ436234), which is 16,486 nt long, was covered with only eight contigs, from which one was long enough to cover 99.86% of the references (Figure S2).
Additionally, in this study, we described the application of the mRT-PCR approach for validation of the sRNA-seq data. Simultaneous amplifications of different combinations of nine viruses or two viroids and satGFLV were performed. According to the KAPA2G Fast Multiplex Kit protocol, employed primers should have a similar temperature melting (Tm) and GC content of 40–60%. In our study, the Tm of primers used for virus amplification was not similar; the lowest Tm had a primer pair for GLRaV-3 amplification (51.81 °C for forward and 48.91 °C for reverse primer) (Table 1). Considering GC content, according to the protocol primers, a GC content higher than 60% may require higher and/or longer denaturation temperature and time, while a GC content lower than 40% may require increased primer concentrations, additional MgCl2 and/or annealing temperature lower than 60 °C (KAPA2G Fast Multiplex PCR Kit, https://www.n-genetics.com/products/1104/1023/12664.pdf, accessed on 1 April 2022). In this study, the lowest GC content had again primers for GLRaV-3 amplification (30%), while all other primers for amplification of predicted viruses in L2, L3, and L4 had GC content in the range of 40–60%. In L1, all primers for virus amplification had a GC content of 40–60%, except for the reverse primer of GLRaV-2 (66.67%) (Table 1). Although the primers in our study had differences in Tm and GC content in all cases, successful amplifications were obtained (Figure 3). Thus far, the highest number of grapevine viral pathogens amplified using mRT-PCR was nine (ArMV, GFLV, GVA, GVB, GRSPaV, GFkV, GLRaV-1, GLRaV-2, and GLRaV-3) [44,47]. Nassuth et al. (2000) [43] reported simultaneous detection of ArMV, GRSPaV, and malate dehydrogenase mRNA for GLRaV-3, GVA, GVB and RubiscoL mRNA. Simultaneous detection of grapevine-infecting viruses belonging to the Nepovirus genus were reported by Digiaro et al. (2007) [45]. Hajizadeh et al. (2012) [46] developed mRT-PCR for simultaneous detection of five grapevine viroids. Simultaneous amplification of viruses and viroids have also been reported: GFLV, GYSVd-1, and GYSVd-2, in addition, HSVd was included instead of plant internal control [48], and for GPGV, GFkV, HSVd and GYSVd-1 [49]. In this study, a cumulative number of viral pathogens was minimum 7 and maximum 9 per library. Considering that viroids may form dimers or even multimers that are also visible on agarose gel, the viroids were separately amplified. Some studies found that mRT-PCR is less sensitive compared to singleplex RT-PCR, which specifically targets one viral pathogen [61,62]. Lower detection sensitivity has also been reported when more than five primer pairs were used in a single reaction to detect stone fruit viruses [36].
However, in this study, we showed that mRT-PCR is highly effective, reliable, and sensitive, enabling validation of all viral pathogens predicted with sRNA-seq.
High-throughput screening and high-throughput validation of viral entities for some important old grapevine varieties from the Ampelographic collection Kromberk, Slovenia, was performed. The mRT-PCR protocol described herein provides a simple, time-saving, cost-efficient method for the rapid and reliable validation of sRNA-seq data and successful detection of viral pathogens belonging to different families and genera.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/v14050921/s1. Table S1: Number of reference viral sequences identified by BLASTN search (VirusDetect pipeline); Figure S1: GFLV (RNA1)-assembled contigs (red bars) mapped to reference sequence (JX513889) (blue bars); Figure S2: GLRaV-2-assembled contigs (red bars) mapped to reference sequence (FJ436234) (blue bars).
Author Contributions
Conceptualization, N.Š. and V.M.; methodology, V.M., J.J., D.R., A.Š. and N.Š.; formal analysis, V.M., J.J. and N.Š.; writing—original draft preparation, V.M.; writing—review and editing, V.M., J.J., D.R., A.Š. and N.Š.; supervision, N.Š.; project administration, N.Š.; funding acquisition, J.J. and N.Š. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Slovenian Research Agency (SRA–ARRS), grant number P4-0077, research program: Genetics and Modern Technologies of Crops, awarded to the research group of the Chair of Genetics, Biotechnology, Statistics and Plant breeding at the University of Ljubljana, Biotechnical Faculty. In addition, a scholarship for nationals of the Western Balkans for postgraduate study in Slovenia (Public Scholarship, Development, Disability and Maintenance Fund of the Republic of Slovenia, 245. Public Announcement) was awarded to V.M.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets generated and analyzed during the current study are available in the NCBI Sequence Read Archive (SRA) repository (https://www.ncbi.nlm.nih.gov/sra/, accessed on 1 April 2022) under the BioProject accession number PRJNA667593.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Fuchs, M. Grapevine Viruses: A Multitude of Diverse Species with Simple but Overall Poorly Adopted Management Solutions in the Vineyard. J. Plant Pathol. 2020, 102, 643–653. [Google Scholar] [CrossRef]
- Read, D.A.; Thompson, G.D.; Cordeur, N.L.; Swanevelder, D.; Pietersen, G. Genomic Characterization of Grapevine Viruses N and O: Novel Vitiviruses from South Africa. Arch. Virol. 2022, 167, 611–614. [Google Scholar] [CrossRef] [PubMed]
- Martelli, G.P. An Overview on Grapevine Viruses, Viroids, and the Diseases They Cause. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Springer: Berlin/Heidelberg, Germany, 2017; pp. 31–46. ISBN 9783319577067. [Google Scholar]
- Zhang, Y.; Singh, K.; Kaur, R.; Qiu, W. Association of a Novel DNA Virus with the Grapevine Vein-Clearing and Vine Decline Syndrome. Phytopathology 2011, 101, 1081–1090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krenz, B.; Thompson, J.R.; Fuchs, M.; Perry, K.L. Complete Genome Sequence of a New Circular DNA Virus from Grapevine. J. Virol. 2012, 86, 7715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al Rwahnih, M.; Dave, A.; Anderson, M.M.; Rowhani, A.; Uyemoto, J.K.; Sudarshana, M.R. Association of a DNA Virus with Grapevines Affected by Red Blotch Disease in California. Phytopathology 2013, 103, 1069–1076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maliogka, V.I.; Olmos, A.; Pappi, P.G.; Lotos, L.; Efthimiou, K.; Grammatikaki, G.; Candresse, T.; Katis, N.I.; Avgelis, A.D. A Novel Grapevine Badnavirus Is Associated with the Roditis Leaf Discoloration Disease. Virus Res. 2015, 203, 47–55. [Google Scholar] [CrossRef]
- Basso, M.F.; da Silva, J.C.F.; Fajardo, T.V.M.; Fontes, E.P.B.; Zerbini, F.M. A Novel, Highly Divergent SsDNA Virus Identified in Brazil Infecting Apple, Pear and Grapevine. Virus Res. 2015, 210, 27–33. [Google Scholar] [CrossRef] [Green Version]
- Andret-Link, P.; Laporte, C.; Valat, L.; Ritzenthaler, C.; Demangeat, G.; Vigne, E.; Laval, V.; Pfeiffer, P.; Stussi-Garaud, C.; Fuchs, M. Grapevine Fanleaf Virus: Still a Major Threat to the Grapevine Industry. J. Plant Pathol. 2004, 86, 183–195. [Google Scholar]
- Maree, H.J.; Almeida, R.P.P.; Bester, R.; Chooi, K.M.; Cohen, D.; Dolja, V.V.; Fuchs, M.F.; Golino, D.A.; Jooste, A.E.C.; Martelli, G.P.; et al. Grapevine Leafroll-Associated Virus 3. Front. Microbiol. 2013, 4, 82. [Google Scholar] [CrossRef] [Green Version]
- Endeshaw, S.T.; Sabbatini, P.; Romanazzi, G.; Schilder, A.C.; Neri, D. Effects of Grapevine Leafroll Associated Virus 3 Infection on Growth, Leaf Gas Exchange, Yield and Basic Fruit Chemistry of Vitis vinifera L. Cv. Cabernet Franc. Sci. Hortic. 2014, 170, 228–236. [Google Scholar] [CrossRef]
- Rumbaugh, A.C.; Sudarshana, M.R.; Oberholster, A. Grapevine Red Blotch Disease Etiology and Its Impact on Grapevine Physiology and Berry and Wine Composition. Horticulturae 2021, 7, 552. [Google Scholar] [CrossRef]
- Cheon, J.Y.; Fenton, M.; Gjerdseth, E.; Wang, Q.; Gao, S.; Krovetz, H.; Lu, L.; Shim, L.; Williams, N.; Lybbert, T.J. Heterogeneous Benefits of Virus Screening for Grapevines in California. Am. J. Enol. Vitic. 2020, 71, 231–241. [Google Scholar] [CrossRef]
- Al Rwahnih, M.; Daubert, S.; Golino, D.; Rowhani, A. Deep Sequencing Analysis of RNAs from a Grapevine Showing Syrah Decline Symptoms Reveals a Multiple Virus Infection That Includes a Novel Virus. Virology 2009, 387, 395–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fajardo, T.V.M.; Silva, F.N.; Eiras, M.; Nickel, O. High-Throughput Sequencing Applied for the Identification of Viruses Infecting Grapevines in Brazil and Genetic Variability Analysis. Trop. Plant Pathol. 2017, 42, 250–260. [Google Scholar] [CrossRef] [Green Version]
- Massart, S.; Candresse, T.; Gil, J.; Lacomme, C.; Predajna, L.; Ravnikar, M.; Reynard, J.S.; Rumbou, A.; Saldarelli, P.; Škoric, D.; et al. A Framework for the Evaluation of Biosecurity, Commercial, Regulatory, and Scientific Impacts of Plant Viruses and Viroids Identified by NGS Technologies. Front. Microbiol. 2017, 8, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hily, J.M.; Candresse, T.; Garcia, S.; Vigne, E.; Tannière, M.; Komar, V.; Barnabé, G.; Alliaume, A.; Gilg, S.; Hommay, G.; et al. High-Throughput Sequencing and the Viromic Study of Grapevine Leaves: From the Detection of Grapevine-Infecting Viruses to the Description of a New Environmental Tymovirales Member. Front. Microbiol. 2018, 9, 1782. [Google Scholar] [CrossRef]
- Giampetruzzi, A.; Roumi, V.; Roberto, R.; Malossini, U.; Yoshikawa, N.; La Notte, P.; Terlizzi, F.; Credi, R.; Saldarelli, P. A New Grapevine Virus Discovered by Deep Sequencing of Virus- and Viroid-Derived Small RNAs in Cv Pinot Gris. Virus Res. 2012, 163, 262–268. [Google Scholar] [CrossRef]
- Blouin, A.G.; Keenan, S.; Napier, K.R.; Barrero, R.A.; MacDiarmid, R.M. Identification of a Novel Vitivirus from Grapevines in New Zealand. Arch. Virol. 2018, 163, 281–284. [Google Scholar] [CrossRef]
- Czotter, N.; Molnar, J.; Szabó, E.; Demian, E.; Kontra, L.; Baksa, I.; Szittya, G.; Kocsis, L.; Deak, T.; Bisztray, G.; et al. NGS of Virus-Derived Small RNAs as a Diagnostic Method Used to Determine Viromes of Hungarian Vineyards. Front. Microbiol. 2018, 9, 9. [Google Scholar] [CrossRef] [Green Version]
- Demian, E.; Jaksa-Czotter, N.; Molnar, J.; Tusnady, G.E.; Kocsis, L.; Varallyay, E. Grapevine Rootstocks Can Be a Source of Infection with Non-Regulated Viruses. Eur. J. Plant Pathol. 2020, 156, 897–912. [Google Scholar] [CrossRef] [Green Version]
- Navrotskaya, E.; Porotikova, E.; Yurchenko, E.; Galbacs, Z.N.; Varallyay, E.; Vinogradova, S. High-Throughput Sequencing of Small Rnas for Diagnostics of Grapevine Viruses and Viroids in Russia. Viruses 2021, 13, 2432. [Google Scholar] [CrossRef] [PubMed]
- Miljanić, V.; Jakše, J.; Kunej, U.; Rusjan, D.; Škvarč, A.; Štajner, N. Virome Status of Preclonal Candidates of Grapevine Varieties (Vitis vinifera L.) From the Slovenian Wine-Growing Region Primorska as Determined by High-Throughput Sequencing. Front. Microbiol. 2022, 13, 830866. [Google Scholar] [CrossRef] [PubMed]
- Eichmeier, A.; Kominkova, M.; Pecenka, J.; Kominek, P. High-Throughput Small RNA Sequencing for Evaluation of Grapevine Sanitation Efficacy. J. Virol. Methods 2019, 267, 66–70. [Google Scholar] [CrossRef] [PubMed]
- Turcsan, M.; Demian, E.; Varga, T.; Jaksa-Czotter, N.; Szegedi, E.; Olah, R.; Varallyay, E. Hts-Based Monitoring of the Efficiency of Somatic Embryogenesis and Meristem Cultures Used for Virus Elimination in Grapevine. Plants 2020, 9, 1782. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Lara, A.; Golino, D.; Al Rwahnih, M. Genomic Characterization of Grapevine Virus J, a Novel Virus Identified in Grapevine. Arch. Virol. 2018, 163, 1965–1967. [Google Scholar] [CrossRef] [Green Version]
- Zherdev, A.V.; Vinogradova, S.V.; Byzova, N.A.; Porotikova, E.V.; Kamionskaya, A.M.; Dzantiev, B.B. Methods for the Diagnosis of Grapevine Viral Infections: A Review. Agriculture 2018, 8, 195. [Google Scholar] [CrossRef] [Green Version]
- Bertolini, E.; Olmos, A.; Martínez, M.C.; Gorris, M.T.; Cambra, M. Single-Step Multiplex RT-PCR for Simultaneous and Colourimetric Detection of Six RNA Viruses in Olive Trees. J. Virol. Methods 2001, 96, 33–41. [Google Scholar] [CrossRef]
- Menzel, W.; Jelkmann, W.; Maiss, E. Detection of Four Apple Viruses by Multiplex RT-PCR Assays with Coamplification of Plant MRNA as Internal Control. J. Virol. Methods 2002, 99, 81–92. [Google Scholar] [CrossRef]
- Tuo, D.; Shen, W.; Yang, Y.; Yan, P.; Li, X.; Zhou, P. Development and Validation of a Multiplex Reverse Transcription PCR Assay for Simultaneous Detection of Three Papaya Viruses. Viruses 2014, 6, 3893–3906. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Liu, X.; Ge, B.; Li, M.; Hong, B. A Multiplex RT-PCR for Simultaneous Detection and Identification of Five Viruses and Two Viroids Infecting Chrysanthemum. Arch. Virol. 2015, 160, 1145–1152. [Google Scholar] [CrossRef] [PubMed]
- Dobhal, S.; Arif, M.; Olson, J.; Mendoza-Yerbafría, A.; Aguilar-Moreno, S.; Perez-Garcia, M.; Ochoa-Corona, F.M. Sensitive Detection and Discrimination Method for Studying Multiple Infections of Five Major Plant Viruses Infecting Ornamental Plants in Nursery Environments. Ann. Appl. Biol. 2015, 166, 286–296. [Google Scholar] [CrossRef]
- Zhang, W.; Zhang, Z.; Fan, G.; Gao, Y.; Wen, J.; Bai, Y.; Qiu, C.; Zhang, S.; Shen, Y.; Meng, X. Development and Application of a Universal and Simplified Multiplex RT-PCR Assay to Detect Five Potato Viruses. J. Gen. Plant Pathol. 2017, 83, 33–45. [Google Scholar] [CrossRef]
- Li, X.; Li, Y.; Hu, W.; Li, Y.; Li, Y.; Chen, S.; Wang, J. Simultaneous Multiplex RT-PCR Detection of Four Viruses Associated with Maize Lethal Necrosis Disease. J. Virol. Methods 2021, 298, 114286. [Google Scholar] [CrossRef]
- Thompson, J.R.; Wetzel, S.; Klerks, M.M.; Vašková, D.; Schoen, C.D.; Špak, J.; Jelkmann, W. Multiplex RT-PCR Detection of Four Aphid-Borne Strawberry Viruses in Fragaria Spp. in Combination with a Plant MRNA Specific Internal Control. J. Virol. Methods 2003, 111, 85–93. [Google Scholar] [CrossRef]
- Sánchez-Navarro, J.A.; Aparicio, F.; Herranz, M.C.; Minafra, A.; Myrta, A.; Pallás, V. Simultaneous Detection and Identification of Eight Stone Fruit Viruses by One-Step RT-PCR. Eur. J. Plant Pathol. 2005, 111, 77–84. [Google Scholar] [CrossRef]
- Roy, A.; Fayad, A.; Barthe, G.; Brlansky, R.H. A Multiplex Polymerase Chain Reaction Method for Reliable, Sensitive and Simultaneous Detection of Multiple Viruses in Citrus Trees. J. Virol. Methods 2005, 129, 47–55. [Google Scholar] [CrossRef]
- Park, K.S.; Bae, Y.J.; Jung, E.J.; Kang, S.J. RT-PCR-Based Detection of Six Garlic Viruses and Their Phylogenetic Relationships. J. Microbiol. Biotechnol. 2005, 15, 1110–1114. [Google Scholar]
- Kwon, J.Y.; Hong, J.S.; Kim, M.J.; Choi, S.H.; Min, B.E.; Song, E.G.; Kim, H.H.; Ryu, K.H. Simultaneous Multiplex PCR Detection of Seven Cucurbit-Infecting Viruses. J. Virol. Methods 2014, 206, 133–139. [Google Scholar] [CrossRef]
- Kwak, H.R.; Kim, M.K.; Shin, J.C.; Lee, Y.J.; Seo, J.K.; Lee, H.U.; Jung, M.N.; Kim, S.H.; Choi, H.S. The Current Incidence of Viral Disease in Korean Sweet Potatoes and Development of Multiplex RT-PCR Assays for Simultaneous Detection of Eight Sweet Potato Viruses. Plant Pathol. J. 2014, 30, 416–424. [Google Scholar] [CrossRef] [Green Version]
- Yao, B.; Wang, G.; Ma, X.; Liu, W.; Tang, H.; Zhu, H.; Hong, N. Simultaneous Detection and Differentiation of Three Viruses in Pear Plants by a Multiplex RT-PCR. J. Virol. Methods 2014, 196, 113–119. [Google Scholar] [CrossRef]
- Ali, R.N.; Dann, A.L.; Cross, P.A.; Wilson, C.R. Multiplex RT-PCR Detection of Three Common Viruses Infecting Orchids. Arch. Virol. 2014, 159, 3095–3099. [Google Scholar] [CrossRef] [PubMed]
- Nassuth, A.; Pollari, E.; Helmeczy, K.; Stewart, S.; Kofalvi, S.A. Improved RNA Extraction and One-Tube RT-PCR Assay for Simultaneous Detection of Control Plant RNA plus Several Viruses in Plant Extracts. J. Virol. Methods 2000, 90, 37–49. [Google Scholar] [CrossRef]
- Gambino, G.; Gribaudo, I. Simultaneous Detection of Nine Grapevine Viruses by Multiplex Reverse Transcription-Polymerase Chain Reaction with Coamplification of a Plant RNA as Internal Control. Phytopathology 2006, 96, 1223–1229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Digiaro, M.; Elbeaino, T.; Martelli, G.P. Development of Degenerate and Species-Specific Primers for the Differential and Simultaneous RT-PCR Detection of Grapevine-Infecting Nepoviruses of Subgroups A, B and C. J. Virol. Methods 2007, 141, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Hajizadeh, M.; Navarro, B.; Bashir, N.S.; Torchetti, E.M.; Di Serio, F. Development and Validation of a Multiplex RT-PCR Method for the Simultaneous Detection of Five Grapevine Viroids. J. Virol. Methods 2012, 179, 62–69. [Google Scholar] [CrossRef]
- Gambino, G. Multiplex Rt-Pcr Method for the Simultaneous Detection of Nine Grapevine Viruses. Methods Mol. Biol. 2015, 1236, 39–47. [Google Scholar] [CrossRef]
- Ahmadi, G.; Hajizadeh, M.; Roumi, V. A Multiplex RT-PCR for Simultaneous Detection of the Agents of Yellow Speckle and Vein Banding Diseases in Grapevine. J. Plant Pathol. 2017, 99, 261–266. [Google Scholar] [CrossRef]
- Komínková, M.; Komínek, P. Development and Validation of RT-PCR Multiplex Detection of Grapevine Viruses and Viroids in the Czech Republic. J. Plant Pathol. 2020, 102, 511–515. [Google Scholar] [CrossRef]
- Zheng, Y.; Gao, S.; Padmanabhan, C.; Li, R.; Galvez, M.; Gutierrez, D.; Fuentes, S.; Ling, K.S.; Kreuze, J.; Fei, Z. VirusDetect: An Automated Pipeline for Efficient Virus Discovery Using Deep Sequencing of Small RNAs. Virology 2017, 500, 130–138. [Google Scholar] [CrossRef]
- Fajardo, T.V.M.; Dianese, É.C.; Eiras, M.; Cerqueira, D.M.; Lopes, D.B.; Ferreira, M.A.S.V.; Martins, C.R.F. Variability of the Coat Protein Gene of Grapevine Leafroll-Associated Virus 3 in Brazil. Fitopatol. Bras. 2007, 32, 335–340. [Google Scholar] [CrossRef]
- Abou-Ghanem, N.; Sabanadzovic, S.; Minafra, A.; Saldarelli, P.; Martelli, G.P. Some Properties of Grapevine Leafroll-Associated Virus 2 and Molecular Organization of the 3′region of the Viral Genome. J. Plant Pathol. 1998, 80, 37–46. [Google Scholar]
- Beuve, M.; Candresse, T.; Tannières, M.; Lemaire, O. First Report of Grapevine Pinot Gris Virus (Gpgv) in Grapevine in France. Plant Dis. 2015, 99, 293. [Google Scholar] [CrossRef] [PubMed]
- Glasa, M.; Predajňa, L.; Wetzel, T.; Rheinpfalz, D.L.R.; Šoltys, K.; Sabanadzovic, S. First Report of Grapevine Rupestris Vein Feathering Virus in Grapevine in Slovakia. Plant Dis. 2019, 103, 170. [Google Scholar] [CrossRef]
- Karthikeyan, G.; Alabi, O.J.; Naidu, R.A. Occurrence of Grapevine Leafroll-Associated Virus 1 in Two Ornamental Grapevine Cultivars in Washington State. Plant Dis. 2011, 95, 613. [Google Scholar] [CrossRef]
- Nolasco, G.; Mansinho, A.; Teixeira Santos, M.; Soares, C.; Sequeira, Z.; Sequeira, C.; Correia, P.K.; Sequeira, O.A. Large Scale Evaluation of Primers for Diagnosis of Rupestris Stem Pitting Associated Virus-1. Eur. J. Plant Pathol. 2000, 106, 311–318. [Google Scholar] [CrossRef]
- Miljanić, V.; Jakše, J.; Beber, A.; Rusjan, D.; Škvarč, A.; Štajner, N. First Report of Grapevine Satellite Virus in Slovenia. J. Plant Pathol. 2021, 103, 1329–1330. [Google Scholar] [CrossRef]
- Čepin, U.; Gutiérrez-Aguirre, I.; Ravnikar, M.; Pompe-Novak, M. Frequency of Occurrence and Genetic Variability of Grapevine Fanleaf Virus Satellite RNA. Plant Pathol. 2016, 65, 510–520. [Google Scholar] [CrossRef]
- Sano, T.; Mimura, R.; Ohshima, K. Phylogenetic Analysis of Hop and Grapevine Isolates of Hop Stunt Viroid Supports a Grapevine Origin for Hop Stunt Disease. Virus Genes 2001, 22, 53–59. [Google Scholar] [CrossRef]
- Ward, L.I.; Burnip, G.M.; Liefting, L.W.; Harper, S.J.; Clover, G.R.G. First Report of Grapevine Yellow Speckle Viroid 1 and Hop Stunt Viroid in Grapevine (Vitis vinifera) in New Zealand. Plant Dis. 2011, 95, 617. [Google Scholar] [CrossRef]
- Tao, Y.; Man, J.; Wu, Y. Development of a Multiplex Polymerase Chain Reaction for Simultaneous Detection of Wheat Viruses and a Phytoplasma in China. Arch. Virol. 2012, 157, 1261–1267. [Google Scholar] [CrossRef]
- Nam, M.; Lee, Y.H.; Park, C.Y.; Lee, M.A.; Bae, Y.S.; Lim, S.; Lee, J.H.; Moon, J.S.; Lee, S.H. Development of Multiplex RT-PCR for Simultaneous Detection of Garlic Viruses and the Incidence of Garlic Viral Disease in Garlic Genetic Resources. Plant Pathol. J. 2015, 31, 90–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).