
Intense Innate Immune Responses and Severe Metabolic Disorders in Chicken Embryonic Visceral Tissues Caused by Infection with Highly Virulent Newcastle Disease Virus Compared to the Avirulent Virus: A Bioinformatics Analysis


 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Embryonated Eggs and Viruses
2.2. Virus Infection, Sample Collection and Preparation
2.3. RNA Extraction, cDNA Library Construction and High Throughput Sequencing
2.4. Bioinformatic Analysis of Sequencing Data
2.5. Quantitative Real-Time PCR (qPCR)
3. Results and Discussion
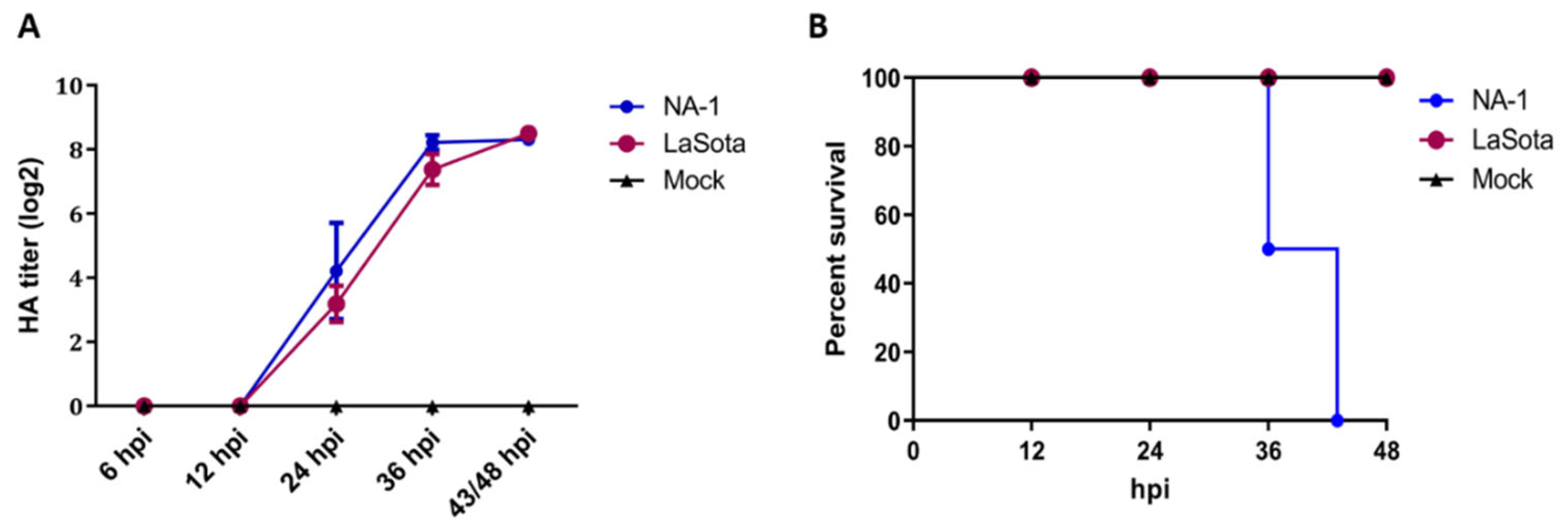
3.1. Distinct Biological Characterization of NDV Highly Virulent Strain NA-1 and Avirulent Vaccine Strain LaSota In Ovo
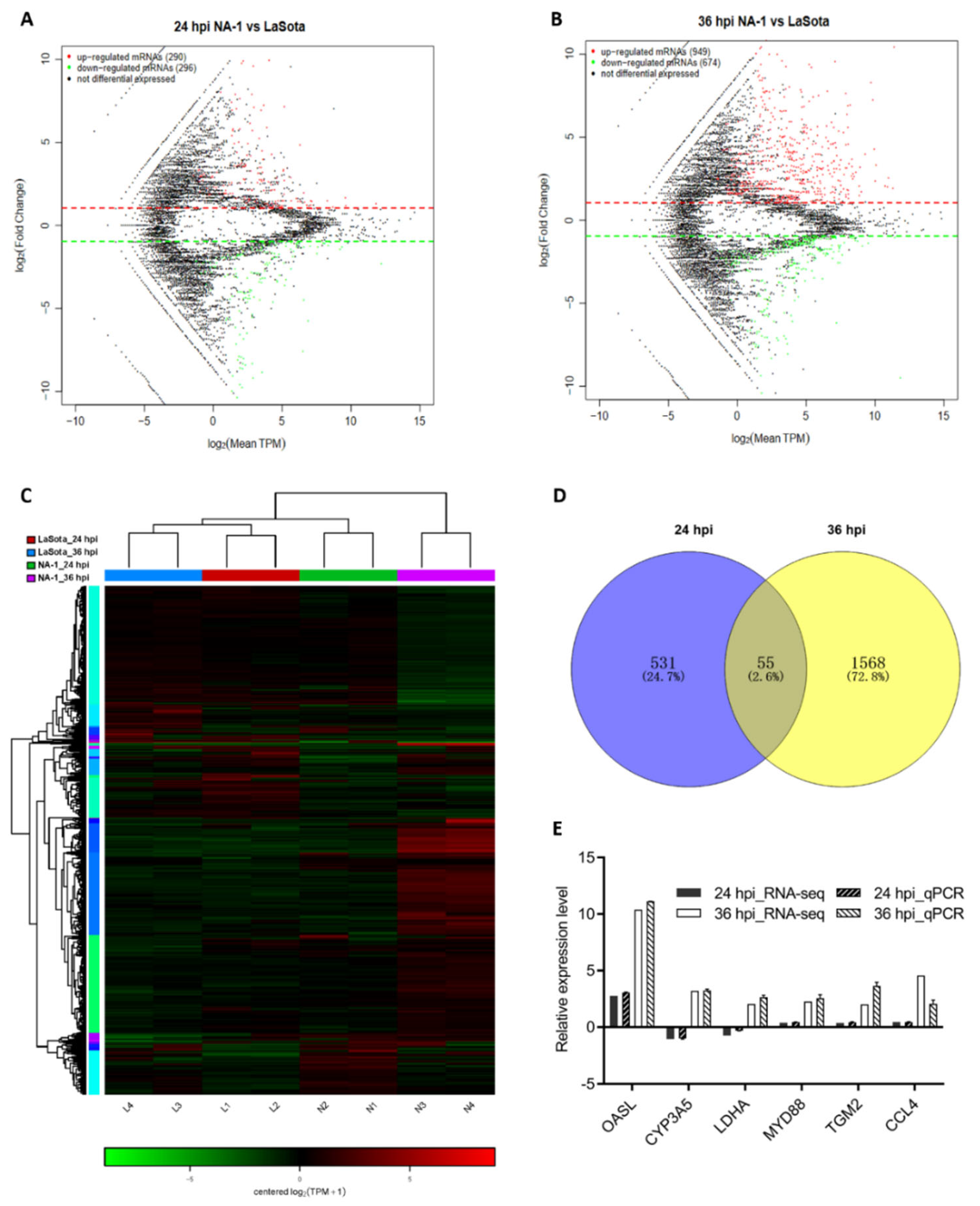
3.2. Global mRNA Expression Patterns in Chicken Embryonic Visceral Tissues after NA-1 and LaSota Infection
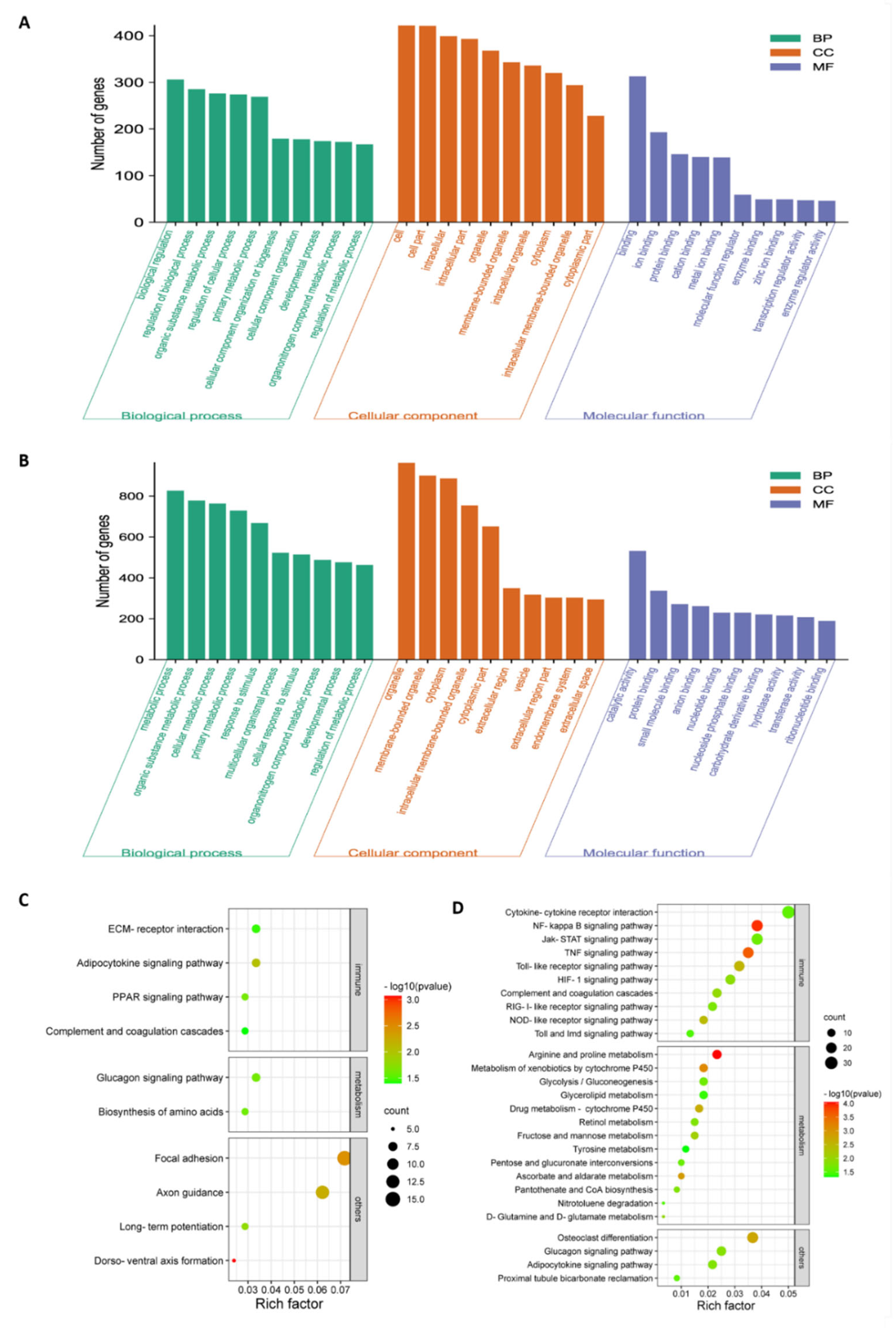
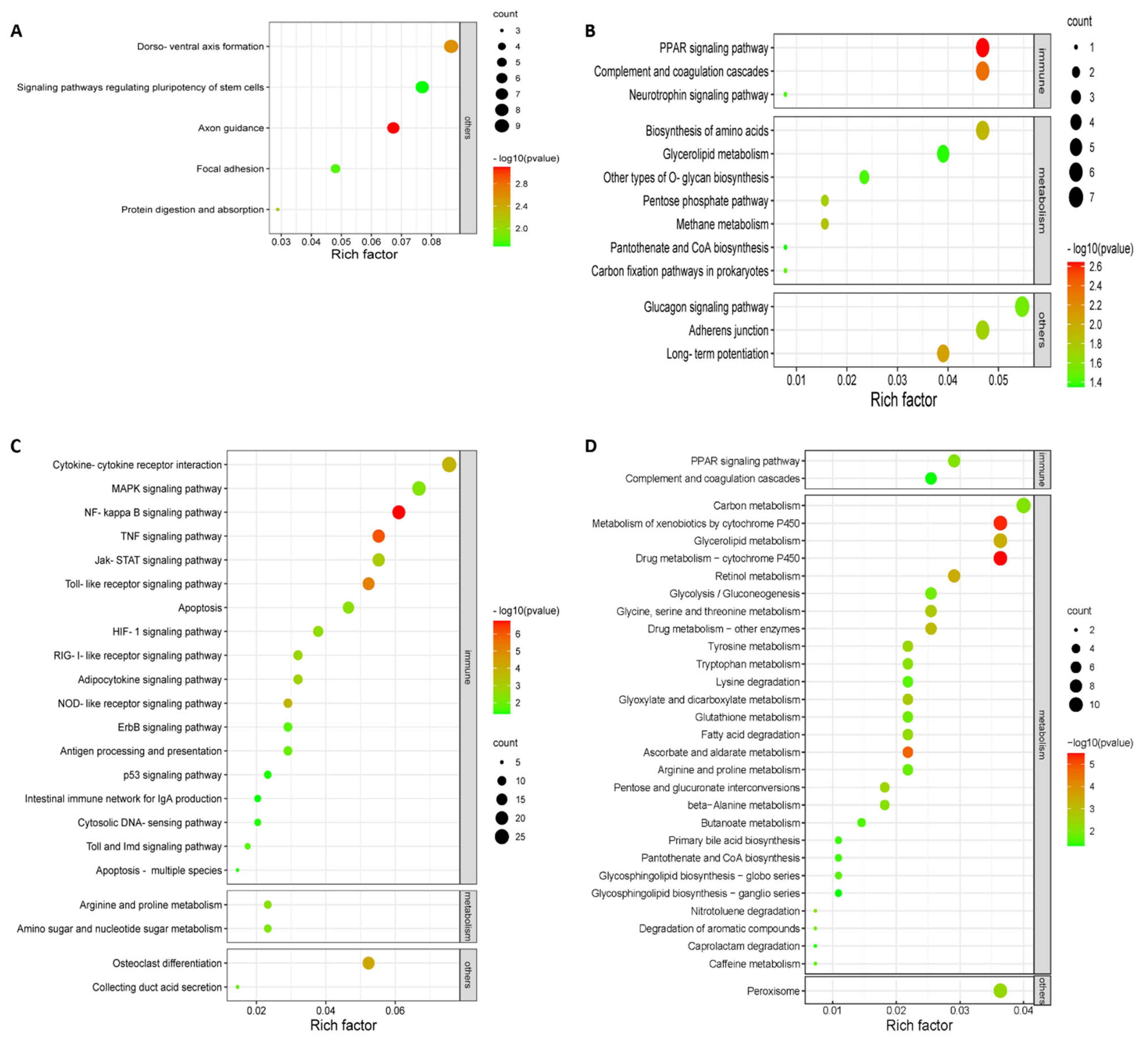
3.3. Highly Virulent Virus Induces a Robust Innate Immune Response in Chicken Embryonic Visceral Tissues Compared to the Avirulent Vaccine Virus
3.4. Severe Metabolic Disorders in Chicken Embryonic Visceral Tissues Caused by Highly Virulent Virus Compared to the Avirulent Vaccine Virus
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Swayne, D.E. Diseases of Poultry, 14th ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2020. [Google Scholar]
- Amarasinghe, G.K.; Ayllón, M.A.; Bào, Y.; Basler, C.F.; Bavari, S.; Blasdell, K.R.; Briese, T.; Brown, P.A.; Bukreyev, A.; Balkema-Buschmann, A.; et al. Taxonomy of the order mononegavirales: Update 2019. Arch. Virol. 2019, 164, 1967–1980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamb, R.A.; Parks, G.D. Paramyxoviridae: The viruses and their replication. In Fields Virology, 6th ed.; Fields, B.N., Knipe, D.M., Howley, P.M., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2013; Volume 1, pp. 957–995. [Google Scholar]
- Saif, Y.M.; Barnes, H.J. Diseases of Poultry, 12th ed.; Saif, Y.M., Fadly, A.M., Glisson, J.R., McDougald, L.R., Nolan, L.K., Swayne, D.E., Eds.; Blackwell: Oxford, UK, 2008; pp. 75–116. [Google Scholar]
- Shittu, I.; Zhu, Z.; Lu, Y.; Hutcheson, J.M.; Stice, S.L.; West, F.D.; Donadeu, M.; Dungu, B.; Fadly, A.M.; Zavala, G.; et al. Development, characterization and optimization of a new suspension chicken-induced pluripotent cell line for the production of Newcastle disease vaccine. Biologicals 2016, 44, 24–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fauzia, E.; Barbhuyan, T.K.; Shrivastava, A.K.; Kumar, M.; Garg, P.; Khan, M.A.; Robertson, A.A.B.; Raza, S.S. Chick embryo: A preclinical model for understanding ischemia-reperfusion Mechanism. Front. Pharm. 2018, 9, 1034. [Google Scholar] [CrossRef] [Green Version]
- Schilling, M.A.; Katani, R.; Memari, S.; Cavanaugh, M.; Buza, J.; Radzio-Basu, J.; Mpenda, F.N.; Deist, M.S.; Lamont, S.J.; Kapur, V. Transcriptional innate immune response of the developing chicken embryo to Newcastle disease virus infection. Front. Genet. 2018, 9, 61. [Google Scholar] [CrossRef] [PubMed]
- Davison, T.F. The immunologists’ debt to the chicken. Br. Poult. Sci. 2003, 44, 6–21. [Google Scholar] [CrossRef]
- Benzarti, E.; Rivas, J.; Sarlet, M.; Franssen, M.; Moula, N.; Savini, G.; Lorusso, A.; Desmecht, D.; Garigliany, M.-M. Usutu Virus Infection of Embryonated Chicken Eggs and a Chicken Embryo-Derived Primary Cell Line. Viruses 2020, 12, 531. [Google Scholar] [CrossRef] [PubMed]
- SSeal, B.S.; King, D.J.; Sellers, H.S. The avian response to Newcastle disease virus. Dev. Comp. Immunol. 2000, 24, 257–268. [Google Scholar] [CrossRef]
- Negash, T.; al-Garib, S.O.; Gruys, E. Comparison of in ovo and post-hatch vaccination with particular reference to infectious bursal disease. A review. Vet. Q. 2004, 26, 76–87. [Google Scholar] [CrossRef]
- Alkie, T.N.; Yitbarek, A.; Hodgins, D.C.; Kulkarni, R.R.; Taha-Abdelaziz, K.; Sharif, S. Development of innate immunity in chicken embryos and newly hatched chicks: A disease control perspective. Avian Pathol. 2019, 48, 288–310. [Google Scholar] [CrossRef]
- Jia, Y.Q.; Wang, X.L.; Wang, X.W.; Yan, C.Q.; Lv, C.J.; Li, X.Q.; Chu, Z.L.; Adam, F.E.A.; Xiao, S.; Zhang, S.X.; et al. Common microRNA(-)mRNA Interactions in Different Newcastle Disease Virus-Infected Chicken Embryonic Visceral Tissues. Int. J. Mol. Sci. 2018, 19, 1291. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Jia, Y.; Wang, X.; Wang, C.; Lv, C.; Li, X.; Chu, Z.; Han, Q.; Xiao, S.; Zhang, S.; et al. MiR-375 has contrasting effects on Newcastle disease virus growth depending on the target gene. Int. J. Biol. Sci. 2019, 15, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Hu, W.; Li, B.; Chen, R.; Shen, W.; Guo, H.; Guo, H.; Li, H. Comparison of viremia, cloacal virus shedding, antibody responses and pathological lesions in adult chickens, quails, and pigeons infected with ALV-A. Sci. Rep. 2019, 9, 3027. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Chang, S.; Ding, Z.; Gao, H.W.; Wan, J.Y.; Liu, W.S.; Liu, L.N.; Gao, Y.; Xu, J. Genomic analysis of Newcastle disease virus strain NA-1 isolated from geese in China. Arch. Virol. 2008, 153, 1281–1289. [Google Scholar] [CrossRef] [PubMed]
- Taylor, L.H. OIE Manual of Diagnostic Tests and Vaccines for Terrestrial Animals; World Organisation for Animal Health: Paris, France, 2014. [Google Scholar]
- Yin, R.; Ding, Z.; Liu, X.; Mu, L.; Cong, Y.; Stoeger, T. Inhibition of Newcastle disease virus replication by RNA interference targeting the matrix protein gene in chicken embryo fibroblasts. J. Virol. Methods 2010, 167, 107–111. [Google Scholar] [CrossRef] [PubMed]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [Green Version]
- Ogata, H.; Goto, S.; Sato, K.; Fujibuchi, W.; Bono, H.; Kanehisa, M. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 27, 29–34. [Google Scholar] [CrossRef] [Green Version]
- Schmittgen, T.D.; Lee, E.J.; Jiang, J.; Sarkar, A.; Yang, L.; Elton, T.S.; Chen, C. Real-time PCR quantification of precursor and mature microRNA. Methods 2008, 44, 31–38. [Google Scholar] [CrossRef] [Green Version]
- Rue, C.A.; Susta, L.; Cornax, I.; Brown, C.C.; Kapczynski, D.R.; Suarez, D.; King, D.J.; Miller, P.J.; Afonso, C.L. Virulent Newcastle disease virus elicits a strong innate immune response in chickens. J. Gen. Virol. 2011, 92, 931–939. [Google Scholar] [CrossRef]
- Liu, W.; Qiu, X.; Song, C.; Sun, Y.; Meng, C.; Liao, Y.; Tan, L.; Ding, Z.; Liu, X.; Ding, C. Deep Sequencing-Based Transcriptome Profiling Reveals Avian Interferon-Stimulated Genes and Provides Comprehensive Insight into Newcastle Disease Virus-Induced Host Responses. Viruses 2018, 10, 162. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Ding, Z.; Liu, X.; Chen, Y.; Li, J.; Tao, Z.; Fei, Y.; Xue, C.; Qian, J.; Wang, X.; et al. Enhanced Replication of Virulent Newcastle Disease Virus in Chicken Macrophages Is due to Polarized Activation of Cells by Inhibition of TLR7. Front. Immunol. 2018, 9, 366. [Google Scholar] [CrossRef]
- Li, R.; Guo, K.; Liu, C.; Wang, J.; Tan, D.; Han, X.; Tang, C.; Zhang, Y.; Wang, J. Strong inflammatory responses and apoptosis in the oviducts of egg-laying hens caused by genotype VIId Newcastle disease virus. BMC Veter. Res. 2016, 12, 255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klasing, K.C. Avian Inflammatory Response: Mediation by Macrophages. Poult. Sci. 1991, 70, 1176–1186. [Google Scholar] [CrossRef] [PubMed]
- Swaggerty, C.L.; Pevzner, I.Y.; Kaiser, P.; Kogut, M.H. Profiling pro-inflammatory cytokine and chemokine mRNA expression levels as a novel method for selection of increased innate immune responsiveness. Vet. Immunol. Immunopathol. 2008, 126, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Tisoncik, J.R.; Korth, M.J.; Simmons, C.P.; Farrar, J.; Martin, T.R.; Katze, M.G. Into the Eye of the Cytokine Storm. Microbiol. Mol. Biol. Rev. 2012, 76, 16–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Elia, R.V.; Harrison, K.; Oyston, P.C.; Lukaszewski, R.A.; Clark, G.C. Targeting the “Cytokine Storm” for Therapeutic Benefit. Clin. Vaccine Immunol. 2013, 20, 319–327. [Google Scholar] [CrossRef]
- Cheng, X.-W.; Lu, J.; Wu, C.-L.; Yi, L.-N.; Xie, X.; Shi, X.-D.; Fang, S.-S.; Zan, H.; Kung, H.-F.; He, M.-L. Three fatal cases of pandemic 2009 influenza A virus infection in Shenzhen are associated with cytokine storm. Respir. Physiol. Neurobiol. 2011, 175, 185–187. [Google Scholar] [CrossRef]
- Iwasaki, A.; Pillai, P.S. Innate immunity to influenza virus infection. Nat. Rev. Immunol. 2014, 14, 315–328. [Google Scholar] [CrossRef]
- Takeuchi, O.; Akira, S. Innate immunity to virus infection. Immunol. Rev. 2009, 227, 75–86. [Google Scholar] [CrossRef]
- Kanneganti, T.D.; Lamkanfi, M.; Núñez, G. Intracellular NOD-like receptors in host defense and disease. Immunity 2007, 27, 549–559. [Google Scholar] [CrossRef]
- Takeuchi, O.; Akira, S. Pattern recognition receptors and inflammation. Cell 2010, 140, 805–820. [Google Scholar] [CrossRef] [Green Version]
- Judge, A.; Dodd, M.S. Metabolism. Underst. Biochem. 2020, 64, 607–647. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, E.L.; Lagunoff, M. Viral activation of cellular metabolism. Virology 2015, 479, 609–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thaker, S.K.; Ch’Ng, J.; Christofk, H.R. Viral hijacking of cellular metabolism. BMC Biol. 2019, 17, 59. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Yin, Y.; Gong, Y.; Qiu, X.; Sun, Y.; Tan, L.; Song, C.; Liu, W.; Liao, Y.; Meng, C.; et al. In Vitro and In Vivo Metabolomic Profiling after Infection with Virulent Newcastle Disease Virus. Viruses 2019, 11, 962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subbaiah, K.C.V.; Valluru, L.; Rajendra, W.; Ramamurthy, C.; Thirunavukkarusu, C.; Subramanyam, R. Newcastle disease virus (NDV) induces protein oxidation and nitration in brain and liver of chicken: Ameliorative effect of vitamin E. Int. J. Biochem. Cell Biol. 2015, 64, 97–106. [Google Scholar] [CrossRef]
- Wang, X.; Jia, Y.; Ren, J.; Liu, H.; Adam, F.A.; Wang, X.; Yang, Z. Insights into the chicken bursa of fabricius response to Newcastle disease virus at 48 and 72 h post-infection through RNA-seq. Vet. Microbiol. 2019, 236, 108389. [Google Scholar] [CrossRef]
- Wang, X.; Jia, Y.; Ren, J.; Liu, H.; Adam, F.A.; Wang, X.; Yang, Z. Hijacking the Supplies: Metabolism as a Novel Facet of Virus-Host Interaction. Front. Immunol. 2019, 10, 1533. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Related Pathways | Upregulated or Downregulated DEGs | NA_1 vs. LaSota_24 hpi | NA_1 vs. LaSota_36 hpi |
|---|---|---|---|
| Host innate immune response | Upregulated | None |
|
| Downregulated |
|
| |
| Metabolism | Upregulated | None |
|
| Downregulated |
|
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, S.; Liu, X.; Mu, J.; Yan, W.; Wang, M.; Chai, H.; Sha, Y.; Jiang, S.; Wang, S.; Ren, Y.; et al. Intense Innate Immune Responses and Severe Metabolic Disorders in Chicken Embryonic Visceral Tissues Caused by Infection with Highly Virulent Newcastle Disease Virus Compared to the Avirulent Virus: A Bioinformatics Analysis. Viruses 2022, 14, 911. https://doi.org/10.3390/v14050911
Cheng S, Liu X, Mu J, Yan W, Wang M, Chai H, Sha Y, Jiang S, Wang S, Ren Y, et al. Intense Innate Immune Responses and Severe Metabolic Disorders in Chicken Embryonic Visceral Tissues Caused by Infection with Highly Virulent Newcastle Disease Virus Compared to the Avirulent Virus: A Bioinformatics Analysis. Viruses. 2022; 14(5):911. https://doi.org/10.3390/v14050911
Chicago/Turabian StyleCheng, Shanyu, Xinxin Liu, Jiaqi Mu, Weiwen Yan, Mengjun Wang, Haoran Chai, Yuxin Sha, Shanshan Jiang, Sijie Wang, Yongning Ren, and et al. 2022. "Intense Innate Immune Responses and Severe Metabolic Disorders in Chicken Embryonic Visceral Tissues Caused by Infection with Highly Virulent Newcastle Disease Virus Compared to the Avirulent Virus: A Bioinformatics Analysis" Viruses 14, no. 5: 911. https://doi.org/10.3390/v14050911
APA StyleCheng, S., Liu, X., Mu, J., Yan, W., Wang, M., Chai, H., Sha, Y., Jiang, S., Wang, S., Ren, Y., Gao, C., Ding, Z., Stoeger, T., Tseren-Ochir, E.-O., Dodovski, A., Alfonso, P., Mingala, C. N., & Yin, R. (2022). Intense Innate Immune Responses and Severe Metabolic Disorders in Chicken Embryonic Visceral Tissues Caused by Infection with Highly Virulent Newcastle Disease Virus Compared to the Avirulent Virus: A Bioinformatics Analysis. Viruses, 14(5), 911. https://doi.org/10.3390/v14050911