
Climate Anomalies and Spillover of Bat-Borne Viral Diseases in the Asia–Pacific Region and the Arabian Peninsula



Abstract
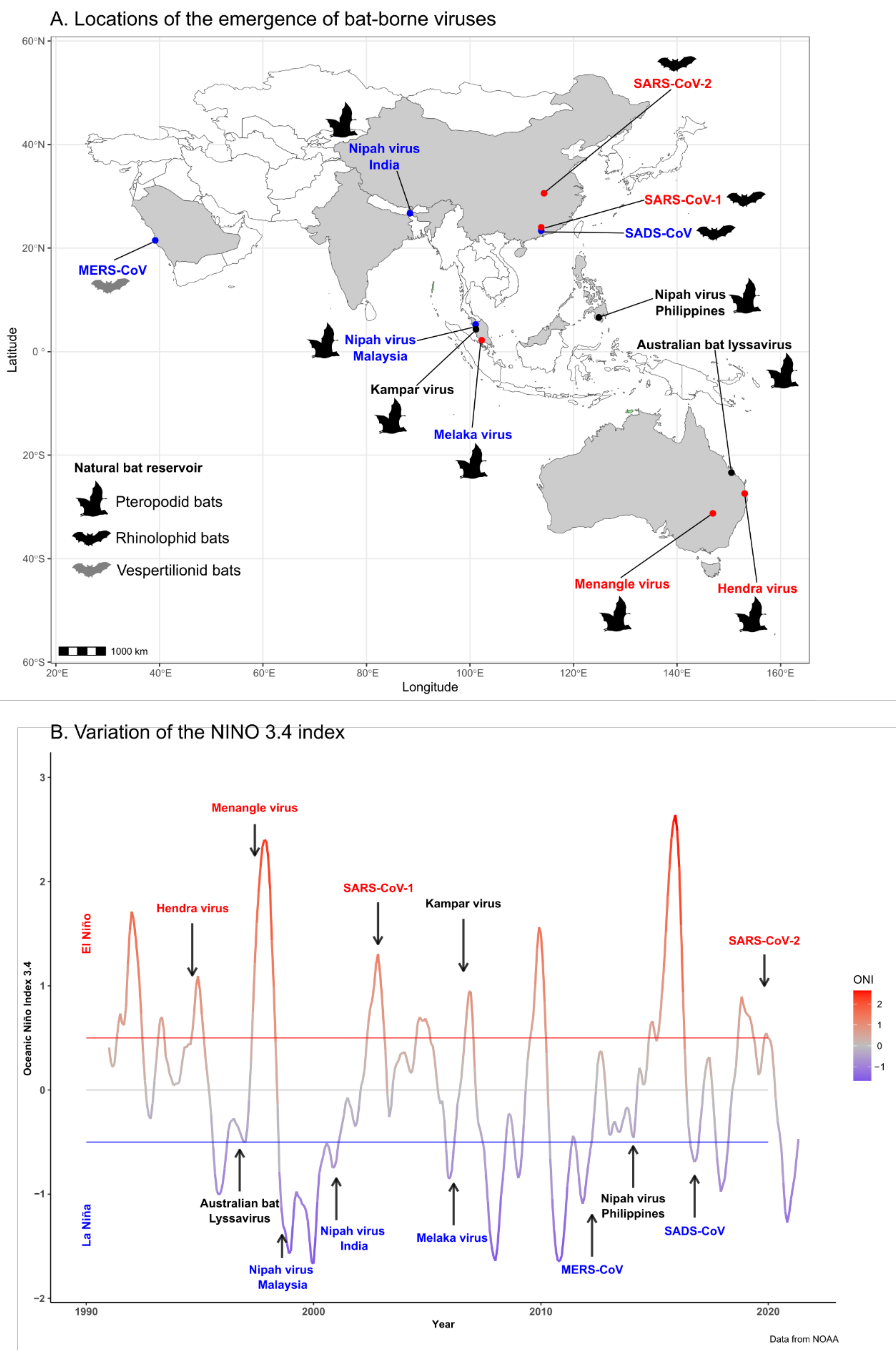
:1. Introduction
2. Materials and Methods
2.1. Data
2.2. Statistical Analyses
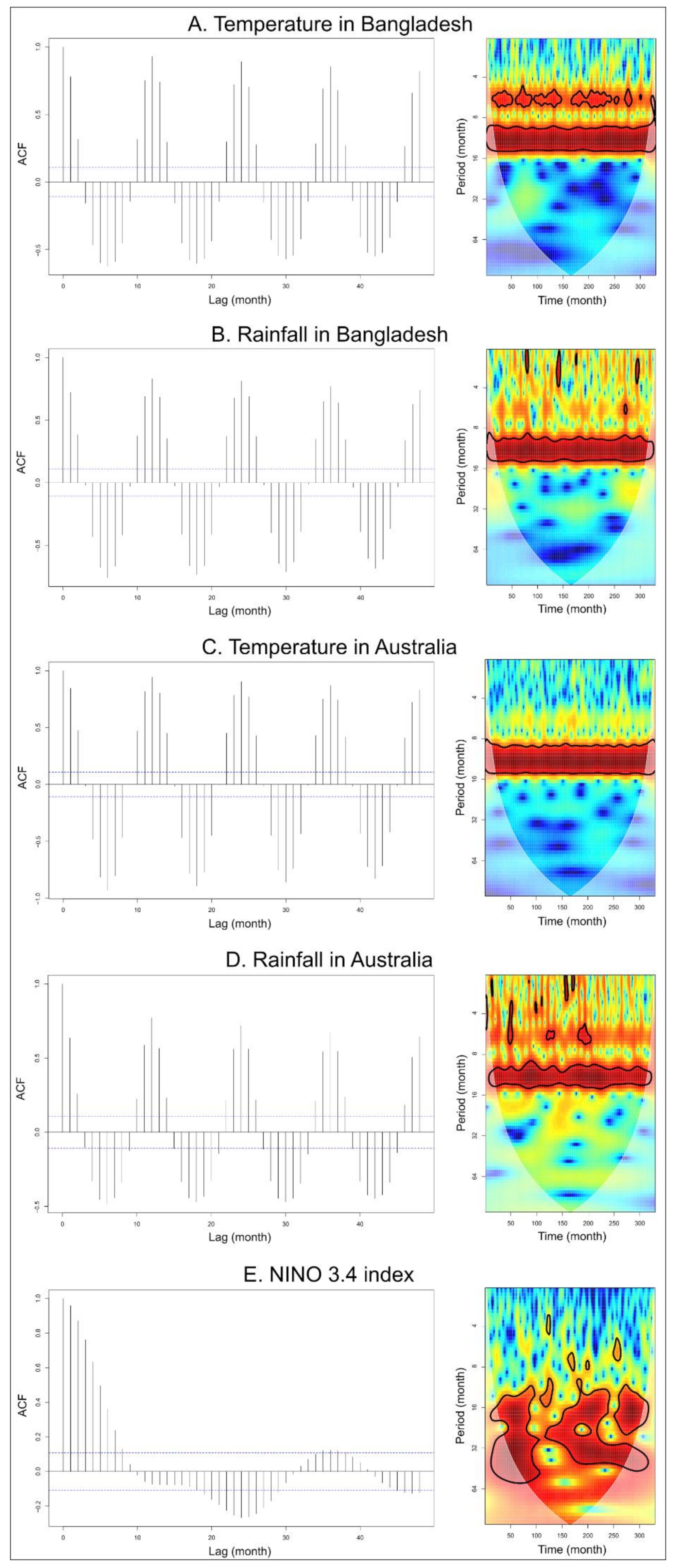
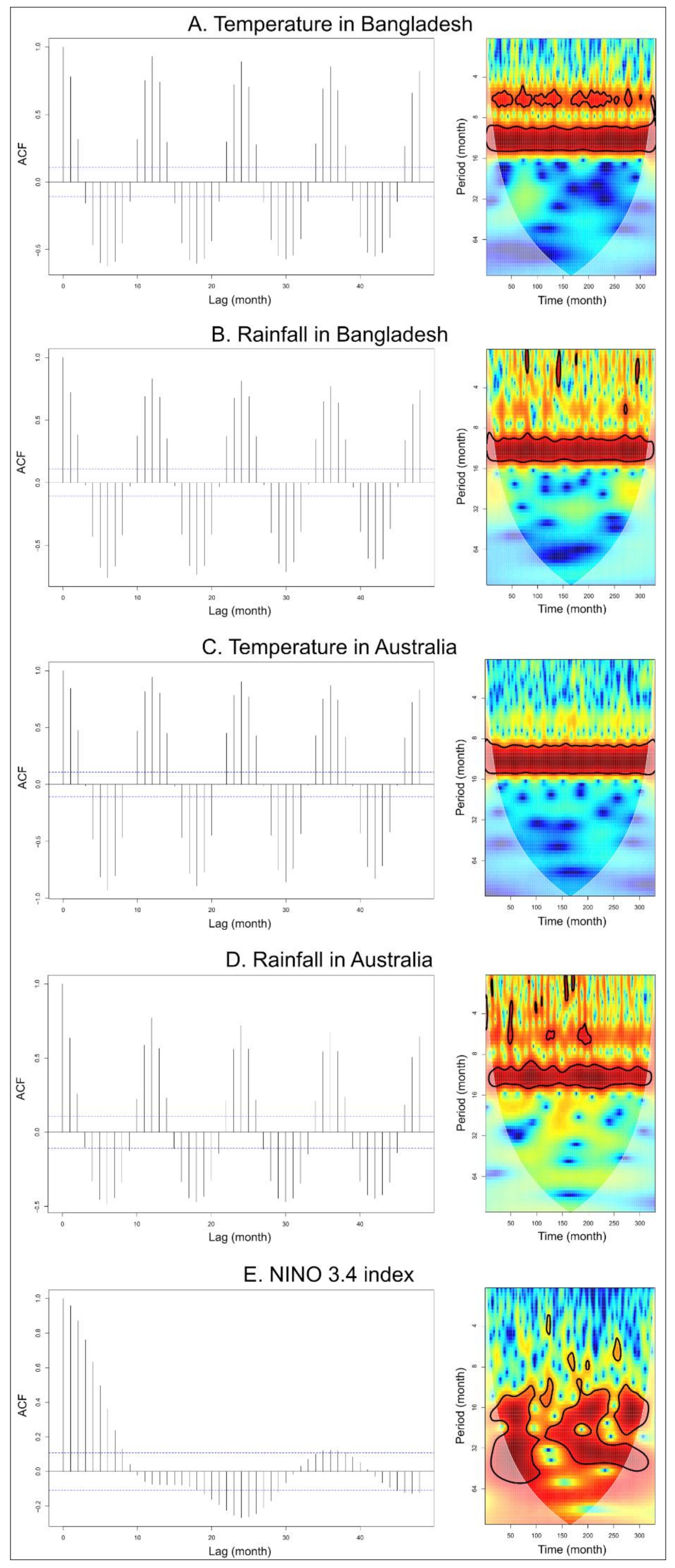
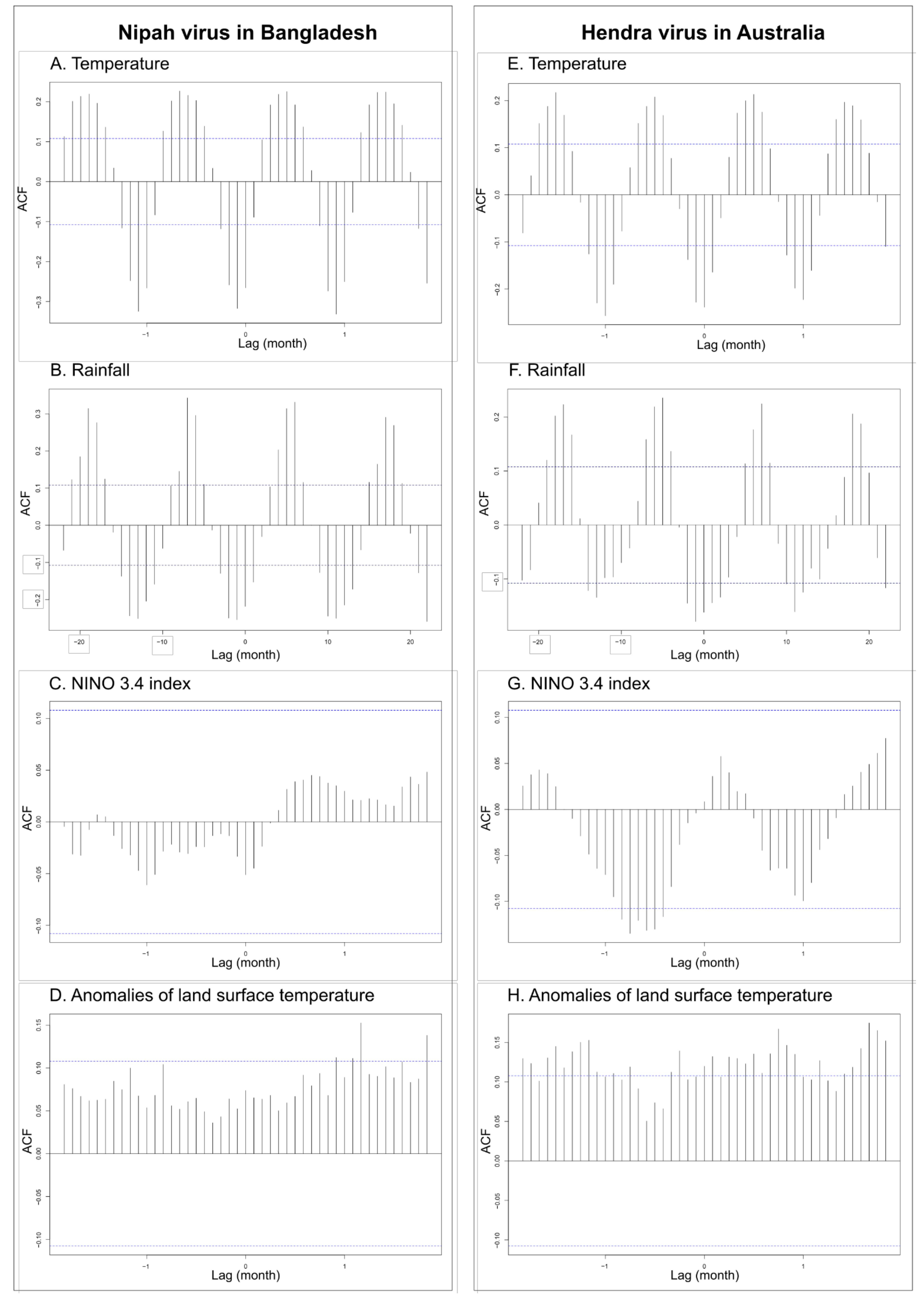
2.2.1. Time-Series Analysis
2.2.2. Logistic Regression with Time Lag Analysis
2.2.3. Structural Equation Modelling
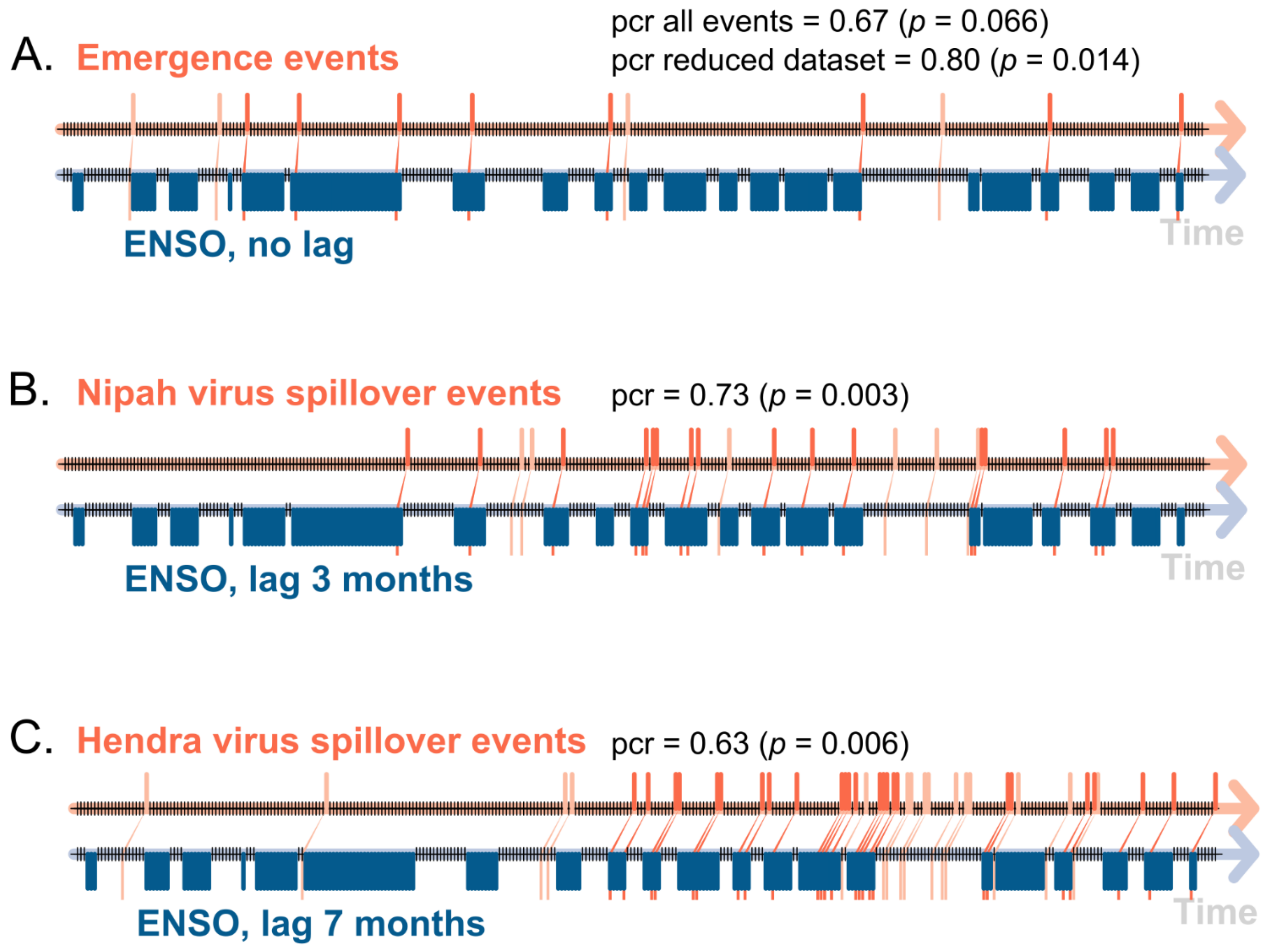
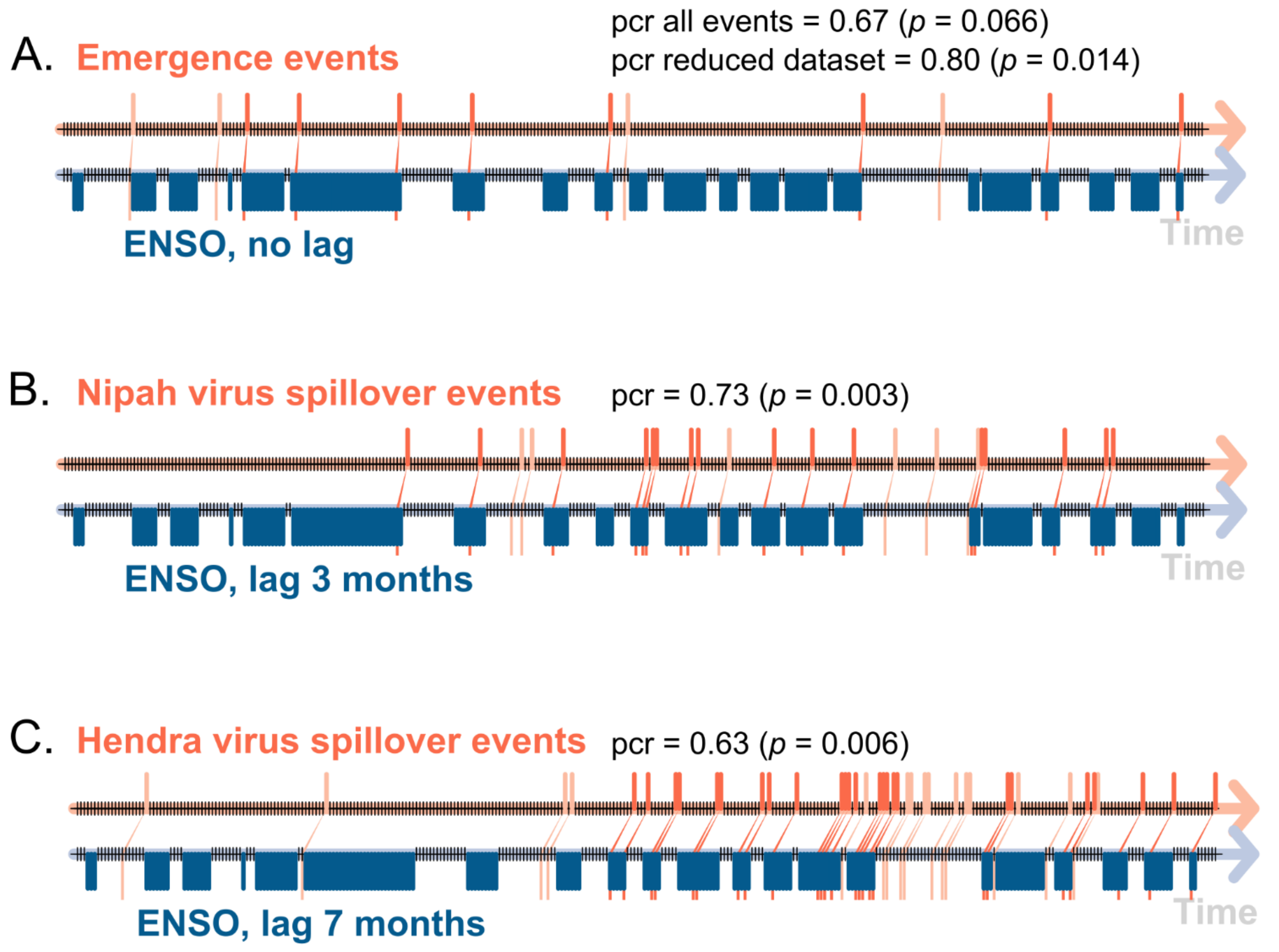
2.2.4. Event Coincidence Analysis
3. Results
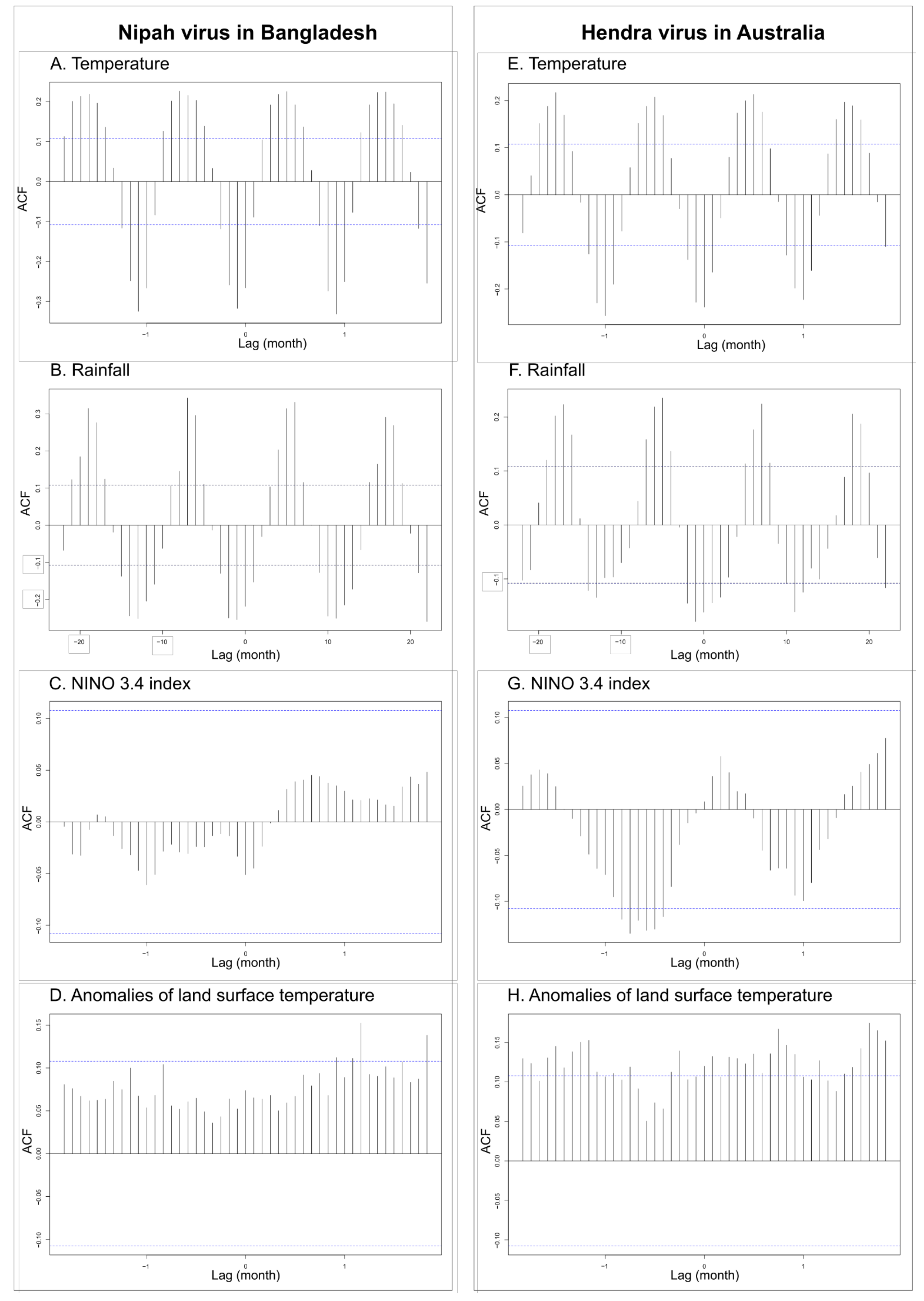
3.1. Time-Series Analyses for HeV and NiV
3.2. Logistic Regression Analyses
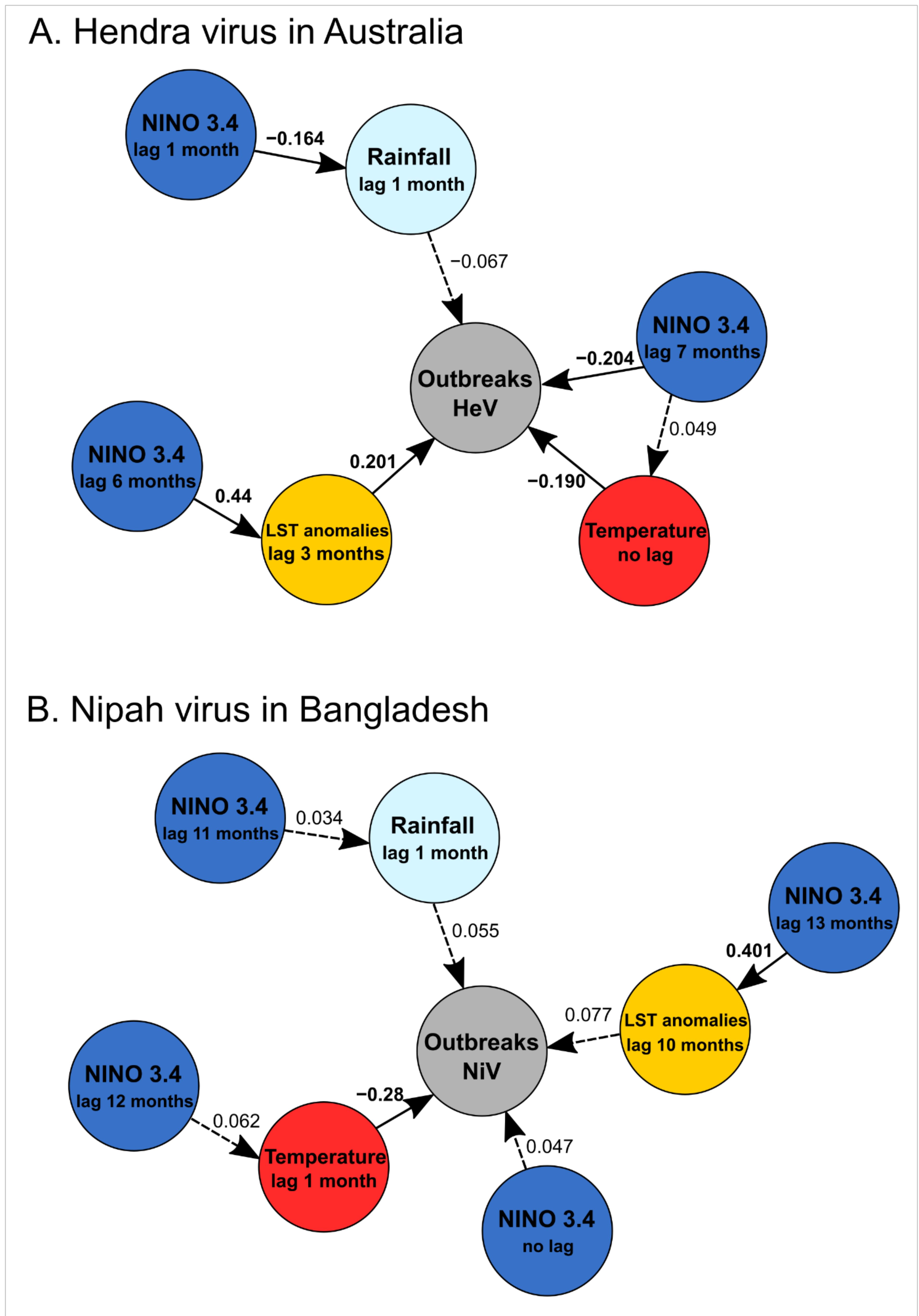
3.3. Structural Equation Modelling
3.4. Event Coincidence Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- McIntyre, K.M.; Setzkorn, C.; Hepworth, P.J.; Morand, S.; Morse, A.P.; Baylis, M. Systematic Assessment of the Climate Sensitivity of Important Human and Domestic Animals Pathogens in Europe. Sci. Rep. 2017, 7, 7134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anyamba, A.; Chretien, J.-P.; Small, J.; Tucker, C.J.; Linthicum, K.J. Developing global climate anomalies suggest potential disease risks for 2006–2007. Int. J. Health Geogr. 2006, 5, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anyamba, A.; Chretien, J.-P.; Britch, S.C.; Soebiyanto, R.P.; Small, J.L.; Jepsen, R.; Forshey, B.M.; Sanchez, J.L.; Smith, R.D.; Harris, R.; et al. Global Disease Outbreaks Associated with the 2015–2016 El Niño Event. Sci. Rep. 2019, 9, 1930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morand, S.; Owers, K.A.; Waret-Szkuta, A.; McIntyre, K.M.; Baylis, M. Climate variability and outbreaks of infectious diseases in Europe. Sci. Rep. 2013, 3, 1774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanley, D.E.; Bourassa, M.A.; O’Brien, J.J.; Smith, S.R.; Spade, E.R. A Quantitative Evaluation of ENSO Indices. J. Clim. 2003, 16, 1249–1258. [Google Scholar] [CrossRef]
- Kogan, F.N. Satellite-Observed Sensitivity of World Land Ecosystems to El Niño/La Niña. Remote Sens. Environ. 2000, 74, 445–462. [Google Scholar] [CrossRef]
- Rifai, S.W.; Girardin, C.A.J.; Berenguer, E.; del Aguila-Pasquel, J.; Dahlsjö, C.A.L.; Doughty, C.E.; Jeffery, K.J.; Moore, S.; Oliveras, I.; Riutta, T.; et al. ENSO Drives interannual variation of forest woody growth across the tropics. Philos. Trans. R. Soc. B Biol. Sci. 2018, 373, 20170410. [Google Scholar] [CrossRef] [Green Version]
- McPhaden, M.J.; Santoso, A.; Cai, W. El Niño Southern Oscillation in a Changing Climate; John Wiley & Sons: Hoboken, NJ, USA, 2020. [Google Scholar]
- Iizumi, T.; Luo, J.-J.; Challinor, A.J.; Sakurai, G.; Yokozawa, M.; Sakuma, H.; Brown, M.; Yamagata, T. Impacts of El Niño Southern Oscillation on the global yields of major crops. Nat. Commun. 2014, 5, 3712. [Google Scholar] [CrossRef] [Green Version]
- Kovats, R.S.; Bouma, M.J.; Hajat, S.; Worrall, E.; Haines, A. El Niño and health. Lancet 2003, 362, 1481–1489. [Google Scholar] [CrossRef]
- Rodó, X.; Pascual, M.; Fuchs, G.; Faruque, A. ENSO and cholera: A nonstationary link related to climate change? Proc. Natl. Acad. Sci. USA 2002, 99, 12901–12906. [Google Scholar] [CrossRef] [Green Version]
- Nicholls, N. A method for predicting murray valley encephalitis in southeast australia using the southern oscillation. Aust. J. Exp. Biol. Med Sci. 1986, 64, 587–594. [Google Scholar] [CrossRef]
- Chaves, L.F.; Calzada, J.E.; Valderrama, A.; Saldana, A. Cutaneous leishmaniasis and sand fly fluctuations are associated with El Niño in Panamá. PLoS Negl. Trop. Dis. 2014, 8, e3210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vincenti-Gonzalez, M.F.; Tami, A.; Lizarazo, E.; Grillet, M.E. ENSO-driven climate variability promotes periodic major outbreaks of dengue in Venezuela. Sci. Rep. 2018, 8, 5727. [Google Scholar] [CrossRef] [PubMed]
- Pramanik, M.; Singh, P.; Kumar, G.; Ojha, V.P.; Dhiman, R.C. El Niño Southern Oscillation as an early warning tool for dengue outbreak in India. BMC Public Health 2020, 20, 1498. [Google Scholar] [CrossRef] [PubMed]
- Caminade, C.; Turner, J.; Metelmann, S.; Hesson, J.C.; Blagrove, M.S.C.; Solomon, T.; Morse, A.P.; Baylis, M. Global risk model for vector-borne transmission of Zika virus reveals the role of El Niño 2015. Proc. Natl. Acad. Sci. USA 2017, 114, 119–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhiman, R.C.; Sarkar, S. El Niño Southern Oscillation as an early warning tool for malaria outbreaks in India. Malar. J. 2017, 16, 122. [Google Scholar] [CrossRef] [Green Version]
- Giles, J.R.; Eby, P.; Parry, H.; Peel, A.J.; Plowright, R.K.; Westcott, D.A.; McCallum, H. Environmental drivers of spatiotemporal foraging intensity in fruit bats and implications for Hendra virus ecology. Sci. Rep. 2018, 8, 9555. [Google Scholar] [CrossRef]
- Daszak, P.; Zambrana-Torrelio, C.; Bogich, T.L.; Fernandez, M.; Epstein, J.H.; Murray, K.A.; Hamilton, H. Interdisciplinary approaches to understanding disease emergence: The past, present, and future drivers of Nipah virus emergence. Proc. Natl. Acad. Sci. USA 2012, 110 (Suppl. 1), 3681–3688. [Google Scholar] [CrossRef] [Green Version]
- Rojas, O.; Li, Y.; Cumani, R. Understanding the Drought Impact of El Niño on the Global Agricultural Areas: An Assessment Using FAO’s Agricultural Stress Index (ASI); Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2014. [Google Scholar]
- Donges, J.F.; Schleussner, C.-F.; Siegmund, J.F.; Donner, R.V. Event coincidence analysis for quantifying statistical interrelationships between event time series. Eur. Phys. J. Spéc. Top. 2016, 225, 471–487. [Google Scholar] [CrossRef] [Green Version]
- Corman, V.M.; Muth, D.; Niemeyer, D.; Drosten, C. Chapter Eight—Hosts and Sources of Endemic Human Coronaviruses. In Advances in Virus Research 100; Kielian, M., Mettenleiter, T.C., Roossinck, M.J., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 163–188. [Google Scholar]
- Rahman, M.; Chakraborty, A. Nipah virus outbreaks in Bangladesh: A deadly infectious disease. WHO South-East Asia J. Public Health 2012, 1, 208–212. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.Z.; Islam, M.M.; Hossain, M.E.; Rahman, M.M.; Islam, A.; Siddika, A.; Sultana, S.; Klena, J.; Flora, M.; Daszak, P.; et al. Genetic diversity of Nipah virus in Bangladesh. Int. J. Infect. Dis. 2021, 102, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Ang, B.S.P.; Lim, T.C.C.; Wang, L.; Kraft, C.S. Nipah Virus Infection. J. Clin. Microbiol. 2018, 56, e01875-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Field, H.; McCall, B.; Barrett, J. Australian Bat Lyssavirus Infection in a Captive Juvenile Black Flying Fox. Emerg. Infect. Dis. 1999, 5, 438–440. [Google Scholar] [CrossRef] [PubMed]
- Chant, K.; Chan, R.; Smith, M.; Dwyer, D.E.; Kirkland, P. Probable human infection with a newly described virus in the family Paramyxoviridae. Emerg. Infect. Dis. 1998, 4, 273–275. [Google Scholar] [CrossRef] [Green Version]
- Ge, X.-Y.; Li, J.-L.; Yang, X.-L.; Chmura, A.A.; Zhu, G.; Epstein, J.H.; Mazet, J.K.; Hu, B.; Zhang, W.; Peng, C.; et al. Isolation and characterization of a bat SARS-like coronavirus that uses the ACE2 receptor. Nature 2013, 503, 535–538. [Google Scholar] [CrossRef]
- Chua, K.B.; Voon, K.; Crameri, G.; Tan, H.S.; Rosli, J.; McEachern, J.A.; Suluraju, S.; Yu, M.; Wang, L.-F. Identification and Characterization of a New Orthoreovirus from Patients with Acute Respiratory Infections. PLoS ONE 2008, 3, e3803. [Google Scholar] [CrossRef]
- Zaki, A.M.; Van Boheemen, S.; Bestebroer, T.M.; Osterhaus, A.D.M.E.; Fouchier, R.A.M. Isolation of a Novel Coronavirus from a Man with Pneumonia in Saudi Arabia. N. Engl. J. Med. 2012, 367, 1814–1820. [Google Scholar] [CrossRef]
- Ching, P.K.G.; de los Reyes, V.C.; Sucaldito, M.N.; Tayag, E.; Columna-Vingno, A.B.; Malbas, F.F.; Bolo, G.C.; Sejvar, J.J.; Eagles, D.; Playford, G.; et al. Outbreak of Henipavirus Infection, Philippines, 2014. Emerg. Infect. Dis. 2015, 21, 328–331. [Google Scholar] [CrossRef]
- Gong, L.; Li, J.; Zhou, Q.; Xu, Z.; Chen, L.; Zhang, Y.; Xue, C.; Wen, Z.; Cao, Y. A New Bat-HKU2-like Coronavirus in Swine, China, 2017. Emerg. Infect. Dis. 2017, 3, 1607–1609. [Google Scholar] [CrossRef] [Green Version]
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R.; et al. A Novel Coronavirus from Patients with Pneumonia in China, 2019. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef]
- Albers, S.; Campitelli, E. Rsoi: Import Various Northern and Southern Hemisphere Climate Indices. R Package Version 05. Available online: https://cran.r-project.org/web/packages/rsoi (accessed on 17 May 2022).
- Bjornstad, O.N.; Cai, J. ncf: Spatial Covariance Functions. R Package Version 12-8. Available online: https://cran.r-project.org/web/packages/ncf (accessed on 17 May 2022).
- Rösch, A.; Schmidbauer, H. WaveletComp 1.1: A Guided Tour through the R Package. Available online: http://www.hs-stat.com/projects/WaveletComp/WaveletComp_guided_tour.pdf (accessed on 17 May 2022).
- Gouhier, T.; Grinsted, A.; Simko, V. biwavelet: Conduct Univariate and Bivariate Wavelet Analyses. R Package Version 02017. Available online: https://cran.r-project.org/web/packages/biwavelet/ (accessed on 17 May 2022).
- R Core Team. R: A language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Venables, W.; Ripley, B.D. Statistics Complements to Modern Applied Statistics with S, 4th ed.; Springer: Berlin/Heidelberg, Germany, 2002. [Google Scholar]
- Lefcheck, J.S. piecewiseSEM: Piecewise structural equation modelling in r for ecology, evolution, and systematics. Methods Ecol. Evol. 2016, 7, 573–579. [Google Scholar] [CrossRef]
- Siegmund, J.F.; Sanders, T.G.; Heinrich, I.; Van der Maaten, E.; Simard, S.; Helle, G.; Donner, R. Meteorological Drivers of Extremes in Daily Stem Radius Variations of Beech, Oak, and Pine in Northeastern Germany: An Event Coincidence Analysis. Front. Plant Sci. 2016, 7, 733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siegmund, J.F.; Siegmund, N.; Donner, R.V. CoinCalc—A new R package for quantifying simultaneities of event series. Comput. Geosci. 2017, 98, 64–72. [Google Scholar] [CrossRef] [Green Version]
- Global Volcanism Program. Report on Mayon (Philippines). In Bulletin of the Global Volcanism Network; Wunderman, R., Ed.; Smithsonian Institution: Washington, DC, USA, 2013; Volume 38. [Google Scholar] [CrossRef]
- Brierley, L.; Vonhof, M.J.; Olival, K.J.; Daszak, P.; Jones, K.E. Quantifying Global Drivers of Zoonotic Bat Viruses: A Process-Based Perspective. Am. Nat. 2016, 187, E53–E64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olival, K.J.; Hosseini, P.R.; Zambrana-Torrelio, C.; Ross, N.; Bogich, T.L.; Daszak, P. Host and viral traits predict zoonotic spillover from mammals. Nature 2017, 546, 646–650. [Google Scholar] [CrossRef]
- Letko, M.; Seifert, S.N.; Olival, K.J.; Plowright, R.K.; Munster, V.J. Bat-borne virus diversity, spillover and emergence. Nat. Rev. Genet. 2020, 18, 461–471. [Google Scholar] [CrossRef]
- Latinne, A.; Hu, B.; Olival, K.J.; Zhu, G.; Zhang, L.; Li, H.; Chmura, A.A.; Field, H.E.; Zambrana-Torrelio, C.; Epstein, J.H.; et al. Origin and cross-species transmission of bat coronaviruses in China. Nat. Commun. 2020, 11, 4235. [Google Scholar] [CrossRef]
- Wells, K.; Morand, S.; Wardeh, M.; Baylis, M. Distinct spread of DNA and RNA viruses among mammals amid prominent role of domestic species. Glob. Ecol. Biogeogr. 2020, 29, 470–481. [Google Scholar] [CrossRef]
- Beyer, R.M.; Manica, A.; Mora, C. Shifts in global bat diversity suggest a possible role of climate change in the emergence of SARS-CoV-1 and SARS-CoV-2. Sci. Total Environ. 2021, 767, 145413. [Google Scholar] [CrossRef]
- Buceta, J.; Johnson, K. Modeling the Ebola zoonotic dynamics: Interplay between enviroclimatic factors and bat ecology. PLoS ONE 2017, 12, e0179559. [Google Scholar] [CrossRef] [Green Version]
- McFarlane, R.; Becker, N.; Field, H. Investigation of the Climatic and Environmental Context of Hendra Virus Spillover Events 1994–2010. PLoS ONE 2011, 6, e28374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurley, E.S.; Hegde, S.T.; Hossain, K.; Sazzad, H.M.S.; Hossain, M.J.; Rahman, M.; Sharker, M.A.Y.; Salje, H.; Islam, M.S.; Epstein, J.H.; et al. Convergence of Humans, Bats, Trees, and Culture in Nipah Virus Transmission, Bangladesh. Emerg. Infect. Dis. 2017, 23, 1446–1453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chowdhury, S.; Khan, S.U.; Crameri, G.; Epstein, J.H.; Broder, C.C.; Islam, A.; Peel, A.J.; Barr, J.; Daszak, P.; Wang, L.-F.; et al. Serological Evidence of Henipavirus Exposure in Cattle, Goats and Pigs in Bangladesh. PLOS Neglected Trop. Dis. 2014, 8, e3302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hegde, S.T.; Sazzad, H.M.S.; Hossain, M.J.; Alam, M.-U.; Kenah, E.; Daszak, P.; Rollin, P.; Rahman, M.; Luby, S.; Gurley, E.S. Investigating Rare Risk Factors for Nipah Virus in Bangladesh: 2001–2012. EcoHealth 2016, 13, 720–728. [Google Scholar] [CrossRef] [Green Version]
- Cortes, M.C.; Cauchemez, S.; Lefrancq, N.; Luby, S.P.; Hossain, M.J.; Sazzad, H.; Rahman, M.; Daszak, P.; Salje, H.; Gurley, E.S. Characterization of the Spatial and Temporal Distribution of Nipah Virus Spillover Events in Bangladesh, 2007–2013. J. Infect. Dis. 2018, 217, 1390–1394. [Google Scholar] [CrossRef]
- McKee, C.D.; Islam, A.; Luby, S.P.; Salje, H.; Hudson, P.J.; Plowright, R.K.; Gurley, E. The Ecology of Nipah Virus in Bangladesh: A Nexus of Land-Use Change and Opportunistic Feeding Behavior in Bats. Viruses 2021, 13, 169. [Google Scholar] [CrossRef]
- Olival, K.J.; Latinne, A.; Islam, A.; Epstein, J.H.; Hersch, R.; Engstrand, R.C.; Gurley, E.S.; Amato, G.; Luby, S.P.; Daszak, P. Population genetics of fruit bat reservoir informs the dynamics, distribution and diversity of Nipah virus. Mol. Ecol. 2020, 29, 970–985. [Google Scholar] [CrossRef]
- Epstein, J.H.; Anthony, S.J.; Islam, A.; Kilpatrick, A.M.; Khan, S.A.; Balkey, M.D.; Ross, N.; Smith, I.; Zambrana-Torrelio, C.; Tao, Y.; et al. Nipah virus dynamics in bats and implications for spillover to humans. Proc. Natl. Acad. Sci. USA 2020, 117, 29190–29201. [Google Scholar] [CrossRef]
- Wahiduzzaman, M.; Luo, J.-J. A statistical analysis on the contribution of El Niño–Southern Oscillation to the rainfall and temperature over Bangladesh. Meteorol. Atmos. Phys. 2021, 133, 55–68. [Google Scholar] [CrossRef]
- Cash, B.A.; Rodó, X.; Kinter, J.L.; Yunus, M. Disentangling the Impact of ENSO and Indian Ocean Variability on the Regional Climate of Bangladesh: Implications for Cholera Risk. J. Clim. 2010, 23, 2817–2831. [Google Scholar] [CrossRef]
- Sharmin, S.; Glass, K.; Viennet, E.; Harley, D. Interaction of Mean Temperature and Daily Fluctuation Influences Dengue Incidence in Dhaka, Bangladesh. PLOS Neglected Trop. Dis. 2015, 9, e0003901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chowdhury, M.R.; Ndiaye, O. Climate change and variability impacts on the forests of Bangladesh—A diagnostic discussion based on CMIP5 GCMs and ENSO. Int. J. Clim. 2017, 37, 4768–4782. [Google Scholar] [CrossRef]
- Islam, A.R.M.T.; Salam, R.; Yeasmin, N.; Kamruzzaman, M.; Shahid, S.; Fattah, M.A.; Uddin, A.S.; Shahariar, M.H.; Mondol, A.H.; Jhajharia, D.; et al. Spatiotemporal distribution of drought and its possible associations with ENSO indices in Bangladesh. Arab. J. Geosci. 2021, 14, 2681. [Google Scholar] [CrossRef]
- Plowright, R.K.; Peel, A.J.; Streicker, D.G.; Gilbert, A.T.; McCallum, H.; Wood, J.; Baker, M.; Restif, O. Transmission or Within-Host Dynamics Driving Pulses of Zoonotic Viruses in Reservoir–Host Populations. PLOS Neglected Trop. Dis. 2016, 10, e0004796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Páez, D.J.; Giles, J.; Mccallum, H.; Field, H.; Jordan, D.; Peel, A.J.; Plowright, R.K. Conditions affecting the timing and magnitude of Hendra virus shedding across pteropodid bat populations in Australia. Epidemiol. Infect. 2017, 145, 3143–3153. [Google Scholar] [CrossRef] [Green Version]
- Becker, D.; Eby, P.; Madden, W.; Peel, A.; Plowright, R. Ecological conditions experienced by bat reservoir hosts predict the intensity of Hendra virus excretion over space and time. bioRxiv 2021. [Google Scholar] [CrossRef]
- Plowright, R.K.; Field, H.E.; Smith, C.; Divljan, A.; Palmer, C.; Tabor, G.; Daszak, P.; Foley, J.E. Reproduction and nutritional stress are risk factors for Hendra virus infection in little red flying foxes (Pteropus scapulatus). Proc. R. Soc. B Boil. Sci. 2008, 275, 861–869. [Google Scholar] [CrossRef] [Green Version]
- McMichael, L.; Edson, D.; Smith, C.; Mayer, D.; Smith, I.; Kopp, S.; Meers, J.; Field, H. Physiological stress and Hendra virus in flying-foxes (Pteropus spp.), Australia. PLoS ONE 2017, 12, e0182171. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, A.; Baker, M.L.; Kulcsar, K.; Misra, V.; Plowright, R.; Mossman, K. Novel Insights into Immune Systems of Bats. Front. Immunol. 2020, 11, 26. [Google Scholar] [CrossRef]
- Edson, D.; Peel, A.J.; Huth, L.; Mayer, D.G.; Vidgen, M.E.; McMichael, L.; Broos, A.; Melville, D.; Kristoffersen, J.; de Jong, C.; et al. Time of year, age class and body condition predict Hendra virus infection in Australian black flying foxes (Pteropus alecto). Epidemiol. Infect. 2019, 147, e240. [Google Scholar] [CrossRef] [Green Version]
- Liqiang, H. El Nino makes trouble for broad areas of China, may hurt rice farmers. China Daily. 2019. Available online: http://www.chinadaily.com.cn/a/201903/01/WS5c7888eba3106c65c34ec1b7.html (accessed on 17 May 2022).
- Tiwari, A.; Singh, S.; Soni, V.; Kumar, R. Environmental Impact of Recent Volcanic Eruption from Mt. Mayon Over 1 South-East Asia. J. Geogr. Nat. Disast. 2021, 10, 547. [Google Scholar]
- Batumbo Boloweti, D.; Giraudoux, P.; Deniel, C.; Garnier, E.; Mauny, F.; Kasereka, C.M.; Kizungu, R.; Muyembe, J.J.; Bompangue, D.; Bornette, G. Volcanic activity controls cholera outbreaks in the East African Rift. PLoS Negl. Trop. Dis. 2020, 14, e0008406. [Google Scholar] [CrossRef] [PubMed]
- Miranda, A.V.; Wiyono, L.; Rocha, I.C.N.; Cedeño, T.D.D.; Lucero-Prisno, D.E.I. Strengthening Virology Research in the Association of Southeast Asian Nations: Preparing for Future Pandemics. Am. J. Trop. Med. Hyg. 2021, 105, 1141–1143. [Google Scholar] [CrossRef] [PubMed]
- Coker, R.J.; Hunter, B.M.; Rudge, J.W.; Liverani, M.; Hanvoravongchai, P. Emerging infectious diseases in southeast Asia: Regional challenges to control. Lancet 2011, 377, 599–609. [Google Scholar] [CrossRef]
- Keesing, F.; Ostfeld, R.S. Impacts of biodiversity and biodiversity loss on zoonotic diseases. Proc. Natl. Acad. Sci. USA 2021, 118, e2023540118. [Google Scholar] [CrossRef]
- Cai, W.; Ng, B.; Wang, G.; Santoso, A.; Wu, L.; Yang, K. Increased ENSO sea surface temperature variability under four IPCC emission scenarios. Nat. Clim. Chang. 2022, 12, 228–231. [Google Scholar] [CrossRef]
- Hu, K.; Huang, G.; Huang, P.; Kosaka, Y.; Xie, S.-P. Intensification of El Niño-induced atmospheric anomalies under greenhouse warming. Nat. Geosci. 2021, 14, 377–382. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
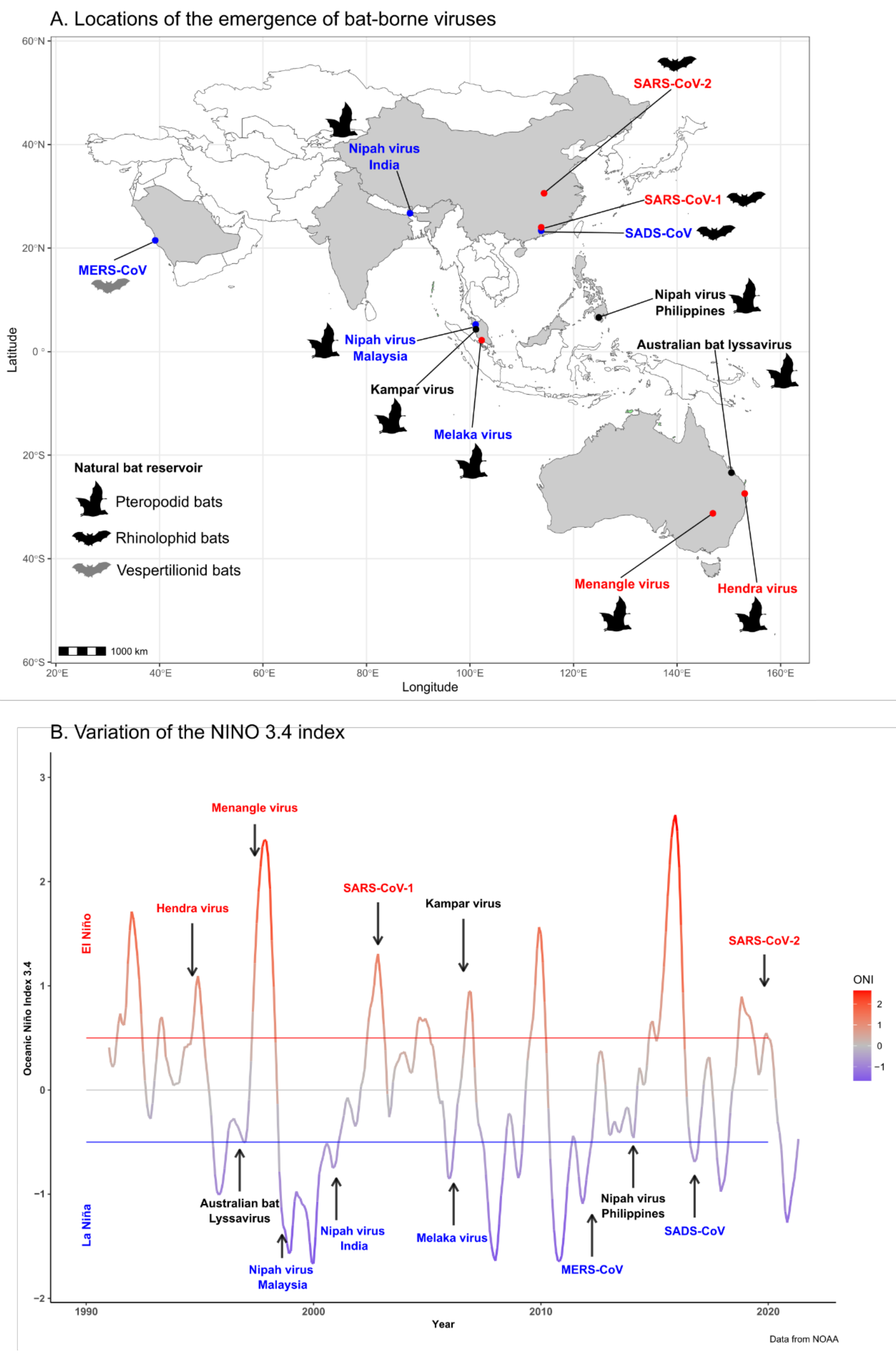
| Emergence | Viral Family | Natural Reservoir | Intermediate host | Date, Location | ENSO Phase | References |
|---|---|---|---|---|---|---|
| Hendra virus | Paramyxoviridae | Pteropodid bats | Horse | Aug 1994, Australia | Warm Phase/El Niño | Giles et al., 2018 [18] |
| Australian bat lyssavirus | Rhabdoviridae | Pteropodid bats | None | Oct 1996, Australia | Neutral Phase | Field et al., 1999 [26] |
| Menangle virus | Paramyxoviridae | Pteropodid bats | Pig | Jun 1997, Australia | Warm Phase/El Niño | Chant et al., 1998 [27] |
| Nipah virus | Paramyxoviridae | Pteropodid bats | Pig | Sep 1998, Malaysia | Cool Phase/La Niña | Ang et al., 2018 [25] |
| Nipah virus | Paramyxoviridae | Pteropodid bats | None | Jan 2001, India | Cool Phase/La Niña | Ang et al., 2018 [25] |
| SARS-CoV-1 | Coronaviridae | Rhinolophid bats | Small carnivores | Nov 2002, China | Warm Phase/El Niño | Ge et al., 2013 [28] |
| Melaka virus | Reoviridae | Pteropodid bats | None | Mar 2006, Malaysia | Cool Phase/La Niña | Chua et al., 2008 [29] |
| Kampar virus | Reoviridae | Pteropodid bats | None | Aug 2006, Malaysia | Neutral Phase | Chua et al., 2008 [29] |
| MERS-CoV | Coronaviridae | Vespertilionid bats | Camel | Apr 2012, Middle East | Cool Phase/La Niña | Zaki et al., 2012 [30] |
| Nipah virus | Paramyxoviridae | Pteropodid bats | Horse | Mar 2014, The Philippines | Neutral phase | Ching et al., 2015 [31] |
| SADS-CoV | Coronaviridae | Rhinolophid bats | Pig (no human cases) | Oct 2016, China | Cool Phase/La Nina | Gong et al., 2017 [32] |
| SARS-CoV-2 | Coronaviridae | Rhinolophid bats | ? | December 2019, China | Warm Phase/El Niño | Zhu et al., 2020 [33] |
| Country | Date (Month/Year) | ENSO Phase |
|---|---|---|
| Bangladesh | April 2001 | Neutral Phase |
| Bangladesh | January 2003 | Warm Phase/El Niño |
| Bangladesh | January 2004 | Neutral Phase |
| Bangladesh | April 2004 | Neutral Phase |
| Bangladesh | January 2005 | Warm Phase/El Niño |
| Bangladesh | January 2007 | Warm Phase/El Niño |
| Bangladesh | March 2007 | Neutral Phase |
| Bangladesh | April 2007 | Neutral Phase |
| India | April 2007 | Neutral Phase |
| Bangladesh | February 2008 | Cool Phase/La Niña |
| Bangladesh | April 2008 | Cool Phase/La Niña |
| Bangladesh | January 2009 | Cool Phase/La Niña |
| Bangladesh | February 2010 | Warm Phase/El Niño |
| Bangladesh | January 2011 | Cool Phase/La Niña |
| Bangladesh | January 2012 | Cool Phase/La Niña |
| Bangladesh | January 2013 | Neutral Phase |
| Bangladesh | January 2014 | Warm Phase/El Niño |
| Bangladesh | January 2015 | Warm Phase/El Niño |
| Bangladesh | February 2015 | Neutral Phase |
| Bangladesh | March 2015 | Warm Phase/El Niño |
| Bangladesh | February 2017 | Neutral Phase |
| Bangladesh | February 2018 | Cool Phase/La Niña |
| Bangladesh | April 2018 | Cool Phase/La Niña |
| India | May 2018 | Neutral Phase |
| India | June 2019 | Neutral Phase |
| Country | Date (Month/Year) | ENSO Phase |
|---|---|---|
| Australia | September 1994 | Warm Phase/El Niño |
| Australia | January 1999 | Cool Phase/La Niña |
| Australia | October 2004 | Warm Phase/El Niño |
| Australia | December 2004 | Warm Phase/El Niño |
| Australia | June 2006 | Neutral Phase |
| Australia | October 2006 | Warm Phase/El Niño |
| Australia | June 2007 | Neutral Phase |
| Australia | July 2007 | Cool Phase/La Niña |
| Australia | June 2008 | Cool Phase/La Niña |
| Australia | July 2008, | Neutral Phase |
| Australia | July 2009 | Neutral Phase |
| Australia | September 2009 | Warm Phase/El Niño |
| Australia | May 2010 | Neutral Phase |
| Australia | June 2011 (4 events) | Cool Phase/La Niña |
| Australia | July 2011 (8 events) | Neutral phase |
| Australia | August 2011 (5 events) | Cool Phase/La Niña |
| Australia | October 2011 | Cool Phase/La Niña |
| Australia | January 2012 | Cool Phase/La Niña |
| Australia | May 2012 (2 events) | Neutral Phase |
| Australia | June 2012 | Neutral Phase |
| Australia | July 2012 (2 events) | Neutral Phase |
| Australia | September 2012 | Neutral Phase |
| Australia | October 2012 | Neutral Phase |
| Australia | January 2013 | Neutral Phase |
| Australia | February 2013 | Neutral Phase |
| Australia | June 2013 (2 events) | Neutral Phase |
| Australia | July 2013 (4 events) | Neutral Phase |
| Australia | March 2014 | Neutral Phase |
| Australia | June 2014 (2 events) | Neutral Phase |
| Australia | July 2014 | Neutral Phase |
| Australia | June 2015 | Warm Phase/El Niño |
| Australia | July 2015 | Warm Phase/El Niño |
| Australia | September 2015 | Warm Phase/El Niño |
| Australia | December 2016 | Cool Phase/La Niña |
| Australia | May 2017 | Neutral Phase |
| Australia | July 2017 | Neutral Phase |
| Australia | August 2017 (2 events) | Neutral Phase |
| Australia | September 2018 | Neutral Phase |
| Australia | June 2019 | Warm Phase/El Niño |
| Australia | June 2020 | Neutral Phase |
| First Time-Series | Second Time-Series | Lag | Correlation (p Value) |
|---|---|---|---|
| Spillover events of HeV (Australia) | NINO 3.4 index | 7 months | 0.13 (0.018) |
| Rainfall (Australia) | 1 month | 0.18 (0.002) | |
| Temperature (Australia) | 0 month | 0.24 (< 0.001) | |
| Land surface temperature anomalies | 3 months | 0.14 (0.012) | |
| Spillover events of NiV (Bangladesh) | NINO 3.4 index Rainfall (Bangladesh) | 0 month 1 month | 0.05 (0.35 0.22 (0.008) |
| Temperature (Bangladesh) | 1 month | 0.32 (0.008) | |
| Land surface temperature anomalies | 10 months | 0.10 (0.047) | |
| NINO 3.4 index | Rainfall (Australia) | 0 month | 0.16 (0.007) |
| Temperature (Australia) | 7 months | 0.08 (0.35) | |
| Rainfall (Bangladesh) | 10 months | 0.03 (0.58) | |
| Temperature (Bangladesh) | 11 months | 0.06 (0.27) | |
| Land surface temperature anomalies | 3 months | 0.40 (<0.0001) |
| Response Variable | Predictor Variable | Estimate (Std Err) | Odds Ratio (2.5–97.5 %) | p | R2 (Global) |
|---|---|---|---|---|---|
| HeV spillover events (Australia) | NINO 3.4 index (lag = 7 months) | −0.72 (0.23) | 0.49 (0.30–0.75) | 0.002 | |
| Rainfall (lag = 1 month) | −0.01 (0.01) | 0.99 (0.96–1.00 | 0.16 | ||
| Temperature (lag = 0 month) | −0.11 (0.04) | 0.90 (0.82–0.98) | 0.013 | ||
| Land surface temperature anomalies (lag = 3 months) | 3.43 (1.03) | 30.96 (4.32–254.30) | 0.001 | 0.21 | |
| NiV spillover events (Bangladesh) | Rainfall (lag = 1 month) | −0.03 (0.01) | 0.97 (0.95–0.99) | 0.008 | 0.30 |
| Model | Response Variable | Predictor Variable | Estimate (Std Err), df | Standardized Estimate | p | R2 (Individual) |
|---|---|---|---|---|---|---|
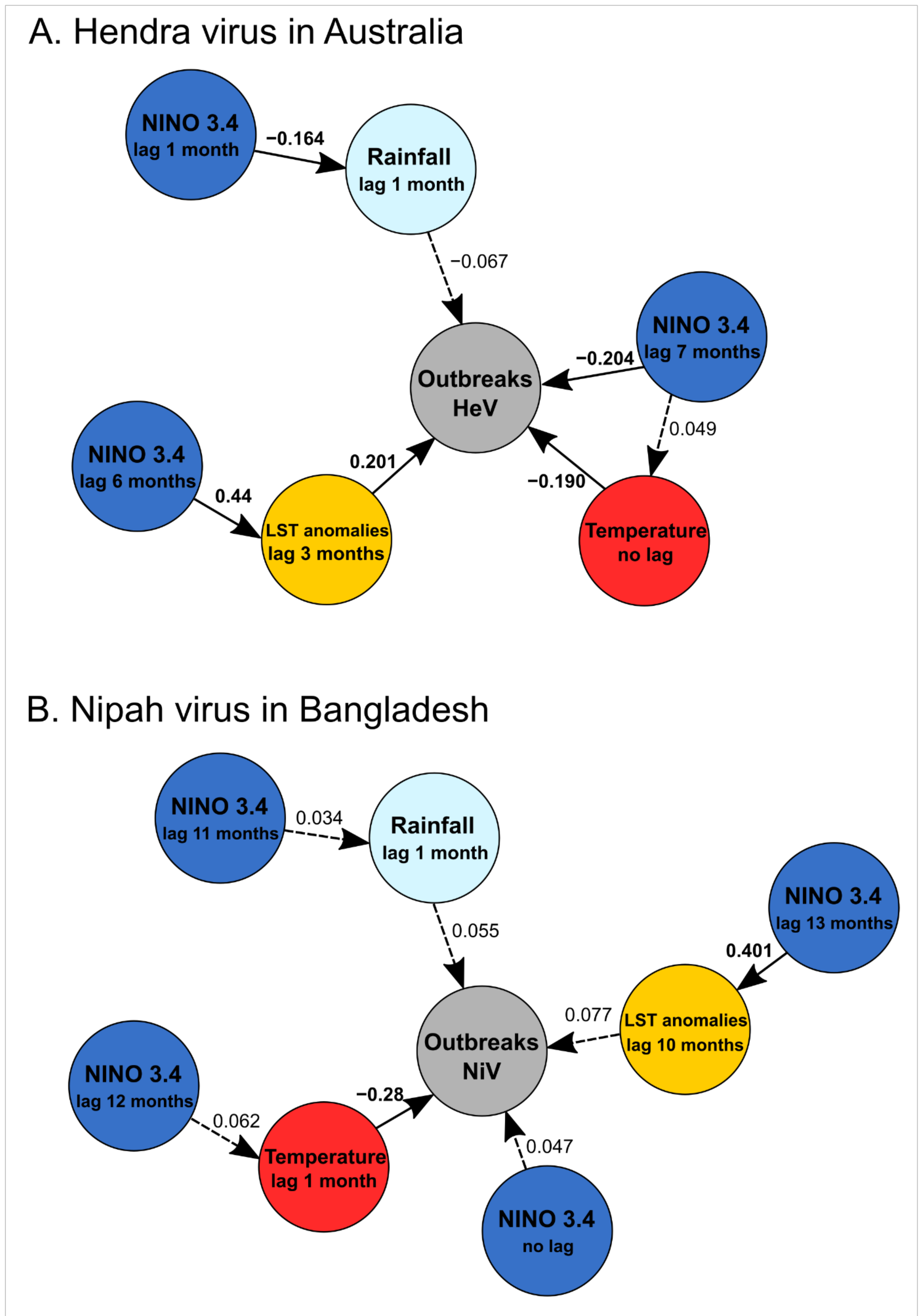
| HeV | Spillover events of HeV | ‘NINO 3.4′ (lag = 7 months) | −0.077 (0.022), 313 | −0.204 | <0.001 | |
| Rainfall (lag = 1 month) | −0.001 (0.001), 313 | −0.067 | 0.28 | |||
| Temperature (lag = 0 month) | −0.078 (0.022), 313 | −0.190 | 0.003 | |||
| LST anom (lag = 3 months) | 0.3123 (0.095), 313 | 0.201 | <0.001 | 0.11 | ||
| Rainfall (lag = 1 month) | ‘NINO 3.4′ (lag = 1 month) | −5.223 (1.7670), 316 | −0.1638 | 0.003 | 0.027 | |
| Temperature | ‘NINO 3.4′ (lag = 7 months) | 0.271 (0.313), 316 | 0.049 | 0.389 | 0.002 | |
| LST anomalies (lag = 3 months) | ‘NINO 3.4′(lag = 6 months) | 0.101 (0.012), 316 | 0.434 | <0.001 | 0.19 | |
| NiV | Spillover events of NiV | ‘NINO 3.4′ (lag = 0 month) | −0.014 (0.015), 312 | −0.887 | 0.38 | |
| Rainfall (lag = 1 month) | −0.0001 (0.0001), 312 | −0.718 | 0.47 | |||
| Temperature (lag = 1 month) | −0.020 (0.005), 312 | −3.689 | 0.003 | |||
| LST anom (lag = 10 month) | 0.098 (0.067), 312 | 1.443 | 0.15 | 0.12 | ||
| Rainfall (lag = 1 month) | NINO 3.4′ (lag = 11 months) | 6.730 (11.178), 315 | 0.602 | 0.55 | 0.001 | |
| Temperature (lag = 1 month) | NINO 3.4′ (lag = 12 months) | 0.254 (0.231), 315 | 1.100 | 0.272 | 0.004 | |
| LST anomalies (lag = 10 months) | ‘NINO 3.4′ (lag = 13 months) | 0.092 (0.012), 315 | 7.774 | <0.001 | 0.16 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Latinne, A.; Morand, S. Climate Anomalies and Spillover of Bat-Borne Viral Diseases in the Asia–Pacific Region and the Arabian Peninsula. Viruses 2022, 14, 1100. https://doi.org/10.3390/v14051100
Latinne A, Morand S. Climate Anomalies and Spillover of Bat-Borne Viral Diseases in the Asia–Pacific Region and the Arabian Peninsula. Viruses. 2022; 14(5):1100. https://doi.org/10.3390/v14051100
Chicago/Turabian StyleLatinne, Alice, and Serge Morand. 2022. "Climate Anomalies and Spillover of Bat-Borne Viral Diseases in the Asia–Pacific Region and the Arabian Peninsula" Viruses 14, no. 5: 1100. https://doi.org/10.3390/v14051100
APA StyleLatinne, A., & Morand, S. (2022). Climate Anomalies and Spillover of Bat-Borne Viral Diseases in the Asia–Pacific Region and the Arabian Peninsula. Viruses, 14(5), 1100. https://doi.org/10.3390/v14051100