
Epidemiology and Molecular Analyses of Influenza B Viruses in Senegal from 2010 to 2019

 ,
,  , , ,
, , ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Clinical Specimens’ Collection
2.2. RNA Extraction and Real Time Reverse Transcription-PCR (qRT-PCR)
2.3. HA/NA Genes Amplification and Sequencing
2.4. Phylogenetic Analysis
2.5. Phylodynamic Analysis
2.6. Aligned Amino Acid Sequence Analysis
2.7. Statistical Analyses
3. Results
3.1. Demographic and Clinical Characteristics of Patients
3.2. Characteristics of Patients Infected with Influenza Viruses
3.3. Characteristics of Patients According the Type of Infection (IAV vs. IBV)
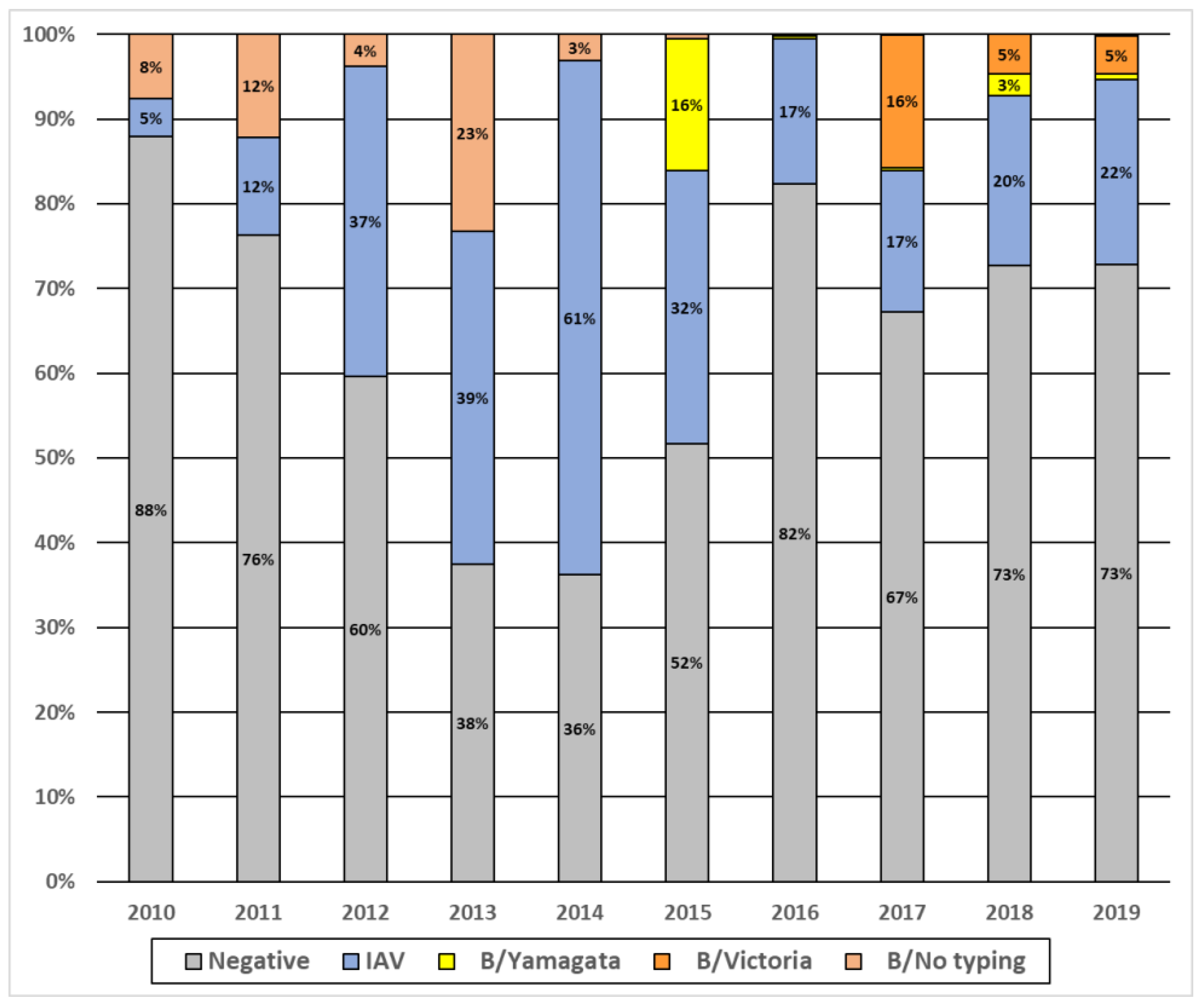
3.4. Pattern of Circulation of Influenza Viruses in Senegal from 2010 to 2019
3.5. Amino Acid Sequences Analysis of IBV Strains Circulating in Senegal
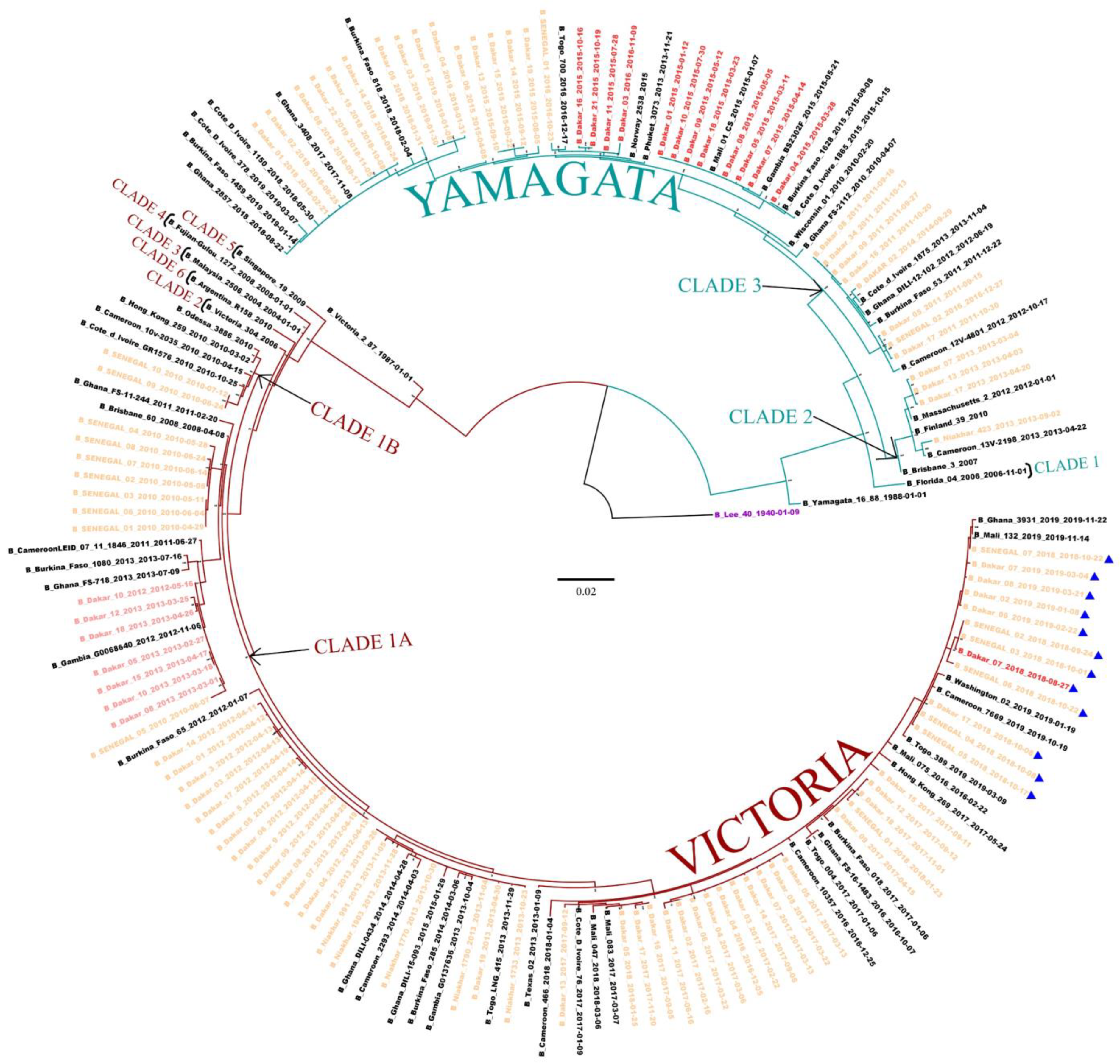
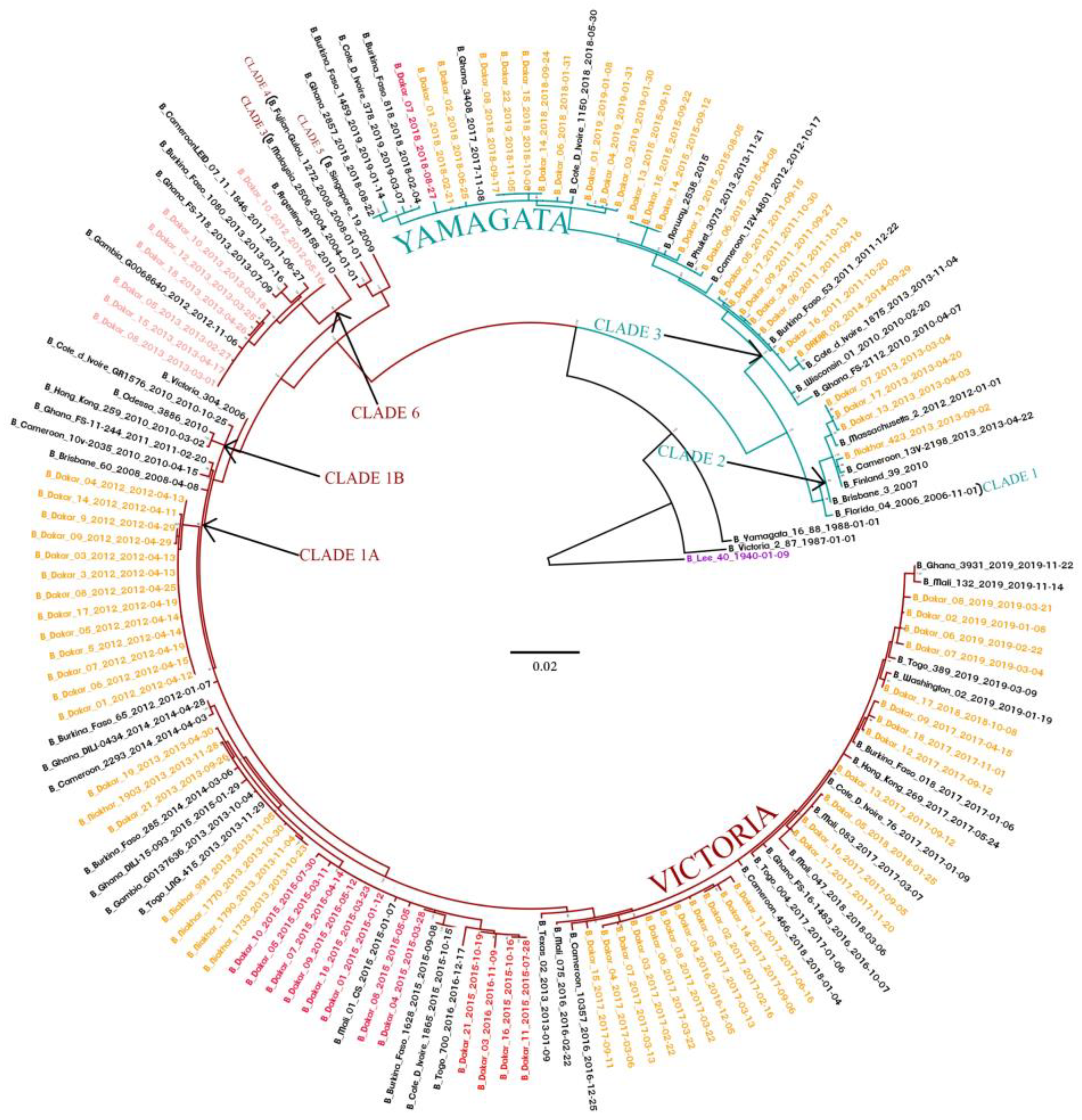
3.6. Phylogenetic Analyses of IVB Strains Circulating in Senegal, 2010–2019
3.7. Bayesian Dated Tree Estimates
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization (WHO). WHO Launches New Global Influenza Strategy; World Health Organization: Geneva, Switzerland, 11 March 2019; Available online: https://www.who.int/news-room/detail/11-03-2019-who-launches-new-global-influenza-strategy (accessed on 19 September 2021).
- Sambala, E.Z.; Mdolo, A.; Banda, R.; Phiri, A.; Wiyeh, A.B.; Wiysonge, C.S. Burden of seasonal influenza in sub-Saharan Africa: A systematic review protocol. BMJ Open 2018, 8, e022949. [Google Scholar] [CrossRef] [PubMed]
- Paul Glezen, W.; Schmier, J.K.; Kuehn, C.M.; Ryan, K.J.; Oxford, J. The Burden of Influenza B: A Structured Literature Review. Am. J. Public Health 2013, 103, e43–e51. [Google Scholar] [CrossRef] [PubMed]
- Seleka, M.; Treurnicht, F.K.; Tempia, S.; Hellferscee, O.; Mtshali, S.; Cohen, A.L.; Buys, A.; McAnerney, J.M.; Besselaar, T.G.; Pretorius, M.; et al. Epidemiology of influenza B/Yamagata and B/Victoria lineages in South Africa, 2005–2014. PLoS ONE 2017, 12, e0177655. [Google Scholar] [CrossRef] [PubMed]
- Irving, S.A.; Patel, D.C.; Kieke, B.A.; Donahue, J.G.; Vandermause, M.F.; Shay, D.K.; Belongia, E.A. Comparison of clinical features and outcomes of medically attended influenza A and influenza B in a defined population over four seasons: 2004–2005 through 2007–2008. Influen. Other Respir Viruses 2012, 6, 37–43. [Google Scholar] [CrossRef]
- McCullers, J.A.; Hayden, F.G. Fatal Influenza B Infections: Time to Reexamine Influenza Research Priorities. J. Infect. Dis. 2012, 205, 870–872. [Google Scholar] [CrossRef]
- World Health Organization. Grippe Saisonnière; World Health Organization: Geneva, Switzerland, 2018; Available online: http://www.who.int/fr/news-room/fact-sheets/detail/influenza-(seasonal) (accessed on 9 October 2018).
- Hause, B.M.; Ducatez, M.; Collin, E.A.; Ran, Z.; Liu, R.; Sheng, Z.; Armien, A.; Kaplan, B.; Chakravarty, S.; Hoppe, A.D.; et al. Isolation of a Novel Swine Influenza Virus from Oklahoma in 2011 Which Is Distantly Related to Human Influenza C Viruses. PLoS Pathog. 2013, 9, e1003176. [Google Scholar] [CrossRef]
- Hay, A.J.; Gregory, V.; Douglas, A.R.; Lin, Y.P. The evolution of human influenza viruses. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2001, 356, 1861–1870. [Google Scholar] [CrossRef]
- Rota, P.A.; Wallis, T.R.; Harmon, M.W.; Rota, J.S.; Kendal, A.P.; Nerome, K. Cocirculation of two distinct evolutionary lineages of influenza type B virus since 1983. Virology 1990, 175, 59–68. [Google Scholar] [CrossRef]
- Chen, R.; Holmes, E.C. The Evolutionary Dynamics of Human Influenza B Virus. J. Mol. Evol. 2008, 66, 655–663. [Google Scholar] [CrossRef]
- Shaw, M.W.; Xu, X.; Li, Y.; Normand, S.; Ueki, R.T.; Kunimoto, G.Y.; Hall, H.; Klimov, A.; Cox, N.J.; Subbarao, K. Reappearance and Global Spread of Variants of Influenza B/Victoria/2/87 Lineage Viruses in the 2000–2001 and 2001–2002 Seasons. Virology 2002, 303, 1–8. [Google Scholar] [CrossRef]
- Langat, P.; Raghwani, J.; Dudas, G.; Bowden, T.A.; Edwards, S.; Gall, A.; Bedford, T.; Rambaut, A.; Daniels, R.S.; Russell, C.A.; et al. Genome-wide evolutionary dynamics of influenza B viruses on a global scale. PLoS Pathog. 2017, 13, e1006749. [Google Scholar] [CrossRef] [PubMed]
- Bedford, T.; Suchard, M.A.; Lemey, P.; Dudas, G.; Gregory, V.; Hay, A.J.; McCauley, J.W.; Russell, C.A.; Smith, D.J.; Rambaut, A. Integrating influenza antigenic dynamics with molecular evolution. eLife 2014, 3, e01914. [Google Scholar] [CrossRef] [PubMed]
- Fall, A.; Ndiaye, N.; Jallow, M.M.; Barry, M.A.; Touré, C.S.B.; Kebe, O.; Kiori, D.E.; Sy, S.; Dia, M.; Goudiaby, D.; et al. Enterovirus D68 Subclade B3 Circulation in Senegal, 2016: Detection from Influenza-like Illness and Acute Flaccid Paralysis Surveillance. Sci. Rep. 2019, 9, 13881. [Google Scholar] [CrossRef]
- Fall, A.; Dia, N.; Kébé, O.; Sarr, F.D.; Kiori, D.E.; Cissé, E.H.A.K.; Sy, S.; Goudiaby, D.; Richard, V.; Diop, O.M.; et al. Enteroviruses and Rhinoviruses: Molecular Epidemiology of the Most Influenza-Like Illness Associated Viruses in Senegal. Am. J. Trop. Med. Hyg. 2016, 95, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Dia, N.; Sarr, F.D.; Thiam, D.; Sarr, T.F.; Espié, E.; OmarBa, I.; Coly, M.; Niang, M.; Richard, V.; 4S Network Group. for the 4S N. Influenza-Like Illnesses in Senegal: Not Only Focus on Influenza Viruses. PLoS ONE 2014, 9, e93227. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Sympos. Ser. 1999, 41, 95–98. [Google Scholar]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Andrew Rambaut Group. FigTree Version 1.4.4—Produce Figures of Phylogenetic Trees; Andrew Rambaut Group. Available online: http://tree.bio.ed.ac.uk/software/figtree (accessed on 6 March 2022).
- Drummond, A.J.; Rambaut, A. BEAST: Bayesian evolutionary analysis by sampling trees. BMC Evolut. Biol. 2007, 8, 32. [Google Scholar] [CrossRef]
- Drummond, A.J.; Rambaut, A.; Shapiro, B.; Pybus, O.G. Bayesian Coalescent Inference of Past Population Dynamics from Molecular Sequences. Mol. Biol. Evol. 2005, 22, 1185–1192. [Google Scholar] [CrossRef]
- Drummond, A.J.; Nicholls, G.K.; Rodrigo, A.G.; Solomon, W. Estimating Mutation Parameters, Population History and Genealogy Simultaneously From Temporally Spaced Sequence Data. Genetics 2002, 161, 1307–1320. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Caini, S.; Huang, Q.S.; Ciblak, M.A.; Kusznierz, G.; Owen, R.; Wangchuk, S.; Henriques, C.M.P.; Njouom, R.; Fasce, R.A.; Yu, H.; et al. Epidemiological and virological characteristics of influenza B: Results of the Global Influenza B Study. Influen. Other Respir. Viruses 2015, 9, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Moa, A.M.; Muscatello, D.J.; Turner, R.M.; MacIntyre, C.R. Epidemiology of influenza B in Australia: 2001–2014 influenza seasons. Influen. Other Respir Viruses 2017, 11, 102–109. [Google Scholar] [CrossRef] [PubMed]
- De Barros, E.N.C.; Cintra, O.; Rossetto, E.; Freitas, L.; Colindres, R. Patterns of influenza B circulation in Brazil and its relevance to seasonal vaccine composition. Braz. J. Infect. Dis. 2016, 20, 81–90. [Google Scholar] [CrossRef][Green Version]
- Lei, N.; Wang, H.-B.; Zhang, Y.-S.; Zhao, J.-H.; Zhong, Y.; Wang, Y.-J.; Huang, L.-Y.; Ma, J.-X.; Sun, Q.; Yang, L.; et al. Molecular evolution of influenza B virus during 2011–2017 in Chaoyang, Beijing, suggesting the free influenza vaccine policy. Sci. Rep. 2019, 9, 2432. [Google Scholar] [CrossRef]
- Emukule, G.O.; Otiato, F.; Nyawanda, B.O.; Otieno, N.A.; Ochieng, C.A.; Ndegwa, L.K.; Muturi, P.; Bigogo, G.; Verani, J.R.; Muthoka, P.M.; et al. The Epidemiology and Burden of Influenza B/Victoria and B/Yamagata Lineages in Kenya, 2012–2016. Open Forum. Infect. Dis. 2019, 6, ofz421. [Google Scholar] [CrossRef]
- Oong, X.Y.; Ng, K.T.; Lam, T.T.-Y.; Pang, Y.K.; Chan, K.G.; Hanafi, N.S.; Kamarulzaman, A.; Tee, K.K. Epidemiological and Evolutionary Dynamics of Influenza B Viruses in Malaysia, 2012–2014. PLoS ONE 2015, 10, e0136254. [Google Scholar] [CrossRef]
- Dapat, C.; Saito, R.; Kyaw, Y.; Naito, M.; Hasegawa, G.; Suzuki, Y.; Dapat, I.C.; Zaraket, H.; Cho, T.M.; Li, D.; et al. Epidemiology of Human Influenza A and B Viruses in Myanmar from 2005 to 2007. Intervirology 2009, 52, 310–320. [Google Scholar] [CrossRef]
- Koul, P.; Potdar, V.; Mir, H.; Chadha, M. The 2015 Outbreak of Severe Influenza in Kashmir, North India: Emergence of a New Clade of A/H1n1 Influenza Virus. PLoS Curr. 2018, 10, e170f. [Google Scholar] [CrossRef]
- Woyessa, A.B.; Mengesha, M.; Belay, D.; Tayachew, A.; Ayele, W.; Beyene, B.; Kassa, W.; Zemelak, E.; Demissie, G.; Amare, B.; et al. Epidemiology of influenza in Ethiopia: Findings from influenza sentinel surveillance and respiratory infection outbreak investigations, 2009–2015. BMC Infect. Dis. 2018, 18, 449. [Google Scholar] [CrossRef]
- Razanajatovo, N.H.; Guillebaud, J.; Harimanana, A.; Rajatonirina, S.; Ratsima, E.H.; Andrianirina, Z.Z.; Rakotoariniaina, H.; Andriatahina, T.; Orelle, A.; Ratovoson, R.; et al. Epidemiology of severe acute respiratory infections from hospital-based surveillance in Madagascar, November 2010 to July 2013. PLoS ONE 2018, 13, e0205124. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Tao, H. Epidemiology and ARIMA model of positive-rate of influenza viruses among children in Wuhan, China: A nine-year retrospective study. Int. J. Infect. Dis. 2018, 74, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Rivas, M.J.; Alegretti, M.; Cóppola, L.; Ramas, V.; Chiparelli, H.; Goñi, N. Epidemiology and Genetic Variability of Circulating Influenza B Viruses in Uruguay, 2012–2019. Microorganisms 2020, 8, 591. [Google Scholar] [CrossRef] [PubMed]
- Heikkinen, T.; Ikonen, N.; Ziegler, T. Impact of Influenza B Lineage-Level Mismatch Between Trivalent Seasonal Influenza Vaccines and Circulating Viruses, 1999–2012. Clin. Infect. Dis. 2014, 59, 1519–1524. [Google Scholar] [CrossRef]
- Horthongkham, N.; Athipanyasilp, N.; Pattama, A.; Kaewnapan, B.; Sornprasert, S.; Srisurapanont, S.; Kantakamalakul, W.; Amaranond, P.; Sutthent, R. Epidemiological, Clinical and Virological Characteristics of Influenza B Virus from Patients at the Hospital Tertiary Care Units in Bangkok during 2011–2014. PLoS ONE 2016, 11, e0158244. [Google Scholar] [CrossRef]
- Coates, B.M.; Staricha, K.L.; Wiese, K.M.; Ridge, K.M. Influenza A Virus Infection, Innate Immunity, and Childhood. JAMA Pediatr. 2015, 169, 956–963. [Google Scholar] [CrossRef]
- Virk, R.K.; Jayakumar, J.; Mendenhall, I.H.; Moorthy, M.; Lam, P.; Linster, M.; Lim, J.; Lin, C.; Oon, L.L.E.; Lee, H.K.; et al. Divergent evolutionary trajectories of influenza B viruses underlie their contemporaneous epidemic activity. Proc. Natl. Acad. Sci. USA 2020, 117, 619–628. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Recommended Composition of Influenza Virus Vaccines for Use in the 2018–2019 Northern Hemisphere Influenza Season; World Health Organization: Geneva, Switzerland, 2018; Available online: https://www.who.int/publications/m/item/recommended-composition-of-influenza-virus-vaccines-for-use-in-the-2018-2019-northern-hemisphere-influenza-season (accessed on 6 March 2022).
- European Centre for Disease Prevention and Control. Influenza Virus Characterisation—Summary Europe, February 2018; ECDC: Stockholm, Sweden, 2018; Available online: https://www.ecdc.europa.eu/sites/default/files/documents/ERLI-Net_report_20-Feb-2018_0.pdf (accessed on 11 May 2022).
- Vijaykrishna, D.; Holmes, E.C.; Joseph, U.; Fourment, M.; Su, Y.C.; Halpin, R.; Lee, R.T.; Deng, Y.-M.; Gunalan, V.; Lin, X.; et al. The contrasting phylodynamics of human influenza B viruses. eLife 2015, 4, e05055. [Google Scholar] [CrossRef]
- Nyasimi, F.M.; Owuor, D.C.; Ngoi, J.M.; Mwihuri, A.G.; Otieno, G.P.; Otieno, J.R.; Githinji, G.; Nyiro, J.U.; Nokes, D.J.; Agoti, C.N. Epidemiological and evolutionary dynamics of influenza B virus in coastal Kenya as revealed by genomic analysis of strains sampled over a single season. Virus Evol. 2020, 6, veaa045. [Google Scholar] [CrossRef]
- Puzelli, S.; Di Martino, A.; Facchini, M.; Fabiani, C.; Calzoletti, L.; Di Mario, G.; Palmieri, A.; Affanni, P.; Camilloni, B.; Chironna, M.; et al. Co-circulation of the two influenza B lineages during 13 consecutive influenza surveillance seasons in Italy, 2004–2017. BMC Infect. Dis 2019, 19, 990. [Google Scholar] [CrossRef]
- Monamele, C.G.; Vernet, M.-A.; Njankouo, M.R.; Kenmoe, S.; Schoenhals, M.; Yahaya, A.A.; Anong, D.N.; Akoachere, J.F.; Njouom, R. Genetic characterization of influenza B virus in Cameroon and high frequency of reassortant strains: Monamele et al. J. Med. Virol. 2018, 90, 1848–1855. [Google Scholar] [CrossRef] [PubMed]
- Tewawong, N.; Suntronwong, N.; Korkong, S.; Theamboonlers, A.; Vongpunsawad, S.; Poovorawan, Y. Evidence for influenza B virus lineage shifts and reassortants circulating in Thailand in 2014–2016. Infect. Genet. Evol. 2017, 47, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Lindstrom, S.E.; Hiromoto, Y.; Nishimura, H.; Saito, T.; Nerome, R.; Nerome, K. Comparative Analysis of Evolutionary Mechanisms of the Hemagglutinin and Three Internal Protein Genes of Influenza B Virus: Multiple Cocirculating Lineages and Frequent Reassortment of the NP, M, and NS Genes. J. Virol. 1999, 73, 4413–4426. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Total (N = 15,156) | Positive (n = 5408) | Negative (n = 9748) | p-Value | Crude OR (95% CI) | Adjusted OR (95% CI) | |
|---|---|---|---|---|---|---|
| Gender n (%) | ||||||
| Female | 7596 (50.1%) | 2827 (52.3%) | 4769 (48.9%) | <0.001 | 1.14 [1.07–1.22] | |
| Male | 7560 (49.9%) | 2581 (47.7%) | 4979 (51.1%) | 1 | ||
| Age Group n (%) | <0.001 | |||||
| [0–5[ | 7907 (52.2%) | 2396 (44.3%) | 5511 (56.5%) | 1 | 1 | |
| [5–15[ | 3372 (22.2%) | 1398 (25.9%) | 1974 (20.3%) | 1.63 (1.50–1.77) | 1.73 (1.58–1.89) | |
| [15–25[ | 1520 (10.0%) | 631 (11.7%) | 889 (9.1%) | 1.63 (1.46–1.83) | 1.77 (1.56–2.01) | |
| [25–50[ | 1772 (11.7%) | 744 (13.8%) | 1028 (10.5%) | 1.66 (1.50–1.85) | 1.81 (1.60–2.04) | |
| 50+ | 585 (3.9%) | 239 (4.4%) | 346 (3.5%) | 1.59 (1.34–1.89) | 1.69 (1.40–2.02) | |
| Symptoms n (%) | ||||||
| Fever | 14,027 (92.6%) | 5046 (93.3%) | 8981 (92.1%) | 0.009 | 1.19 (1.05–1.36) | 1.24 (1.09–1.42) |
| Cough | 12,408 (81.9%) | 4562 (84.4%) | 7846 (80.5%) | <0.001 | 1.31 (1.20–1.43) | 1.36 (1.24–1.49) |
| Rhinorrhea | 6416 (42.3%) | 2431 (45.0%) | 3985 (40.9%) | <0.001 | 1.18 (1.10–1.26) | 1.28 (1.19–1.38) |
| Headache | 2585 (17.1%) | 1120 (20.7%) | 1465 (15.0%) | <0.001 | 1.48 (1.35–1.61) | 1.25 (1.13–1.38) |
| Sore throat | 2401 (15.8%) | 806 (14.9%) | 1595 (16.4%) | 0.020 | 0.90 (0.82–0.98) | 0.71 (0.64–0.78) |
| Myalgia | 2110 (13.9%) | 900 (16.6%) | 1210 (12.4%) | <0.001 | 1.41 (1.28–1.55) | 1.41 (1.28–1.55) |
| Vomiting | 1182 (7.8%) | 362 (6.7%) | 820 (8.4%) | <0.001 | 0.78 (0.69–0.89) | 0.79 (0.70–0.91) |
| Dyspnea | 333 (2.2%) | 129 (2.4%) | 204 (2.1%) | 0.263 | 1.14 (0.91–1.43) | - |
| Arthralgia | 290 (1.9%) | 116 (2.1%) | 174 (1.8%) | 0.137 | 1.21 (0.95–1.53) | 0.79 (0.61–1.01) |
| Conjunctivitis | 113 (0.7%) | 20 (0.4%) | 93 (1.0%) | <0.001 | 0.39 (0.23–0.61) | 0.41 (0.25–0.66) |
| All Influenza (N = 5315) | Influenza A (n = 3993) | Influenza B (n = 1322) | p-Value | Crude OR (95% CI) | Adjusted OR (95% CI) | |
|---|---|---|---|---|---|---|
| Gender n (%) | ||||||
| Female | 2773 (52.2%) | 2100 (52.6%) | 673 (50.9%) | 0.303 | 1 | |
| Male | 2542 (47.8%) | 1893 (47.4%) | 649 (49.1%) | 0.93 (0.83–1.06) | ||
| Age Group n (%) | <0.001 | |||||
| [0–5[ | 2358 (44.4%) | 1789 (44.8%) | 569 (43.0%) | 1 | ||
| [5–15[ | 1380 (26.0%) | 912 (22.8%) | 468 (35.4%) | 0.62 (0.54–0.72) | 0.56 (0.48–0.65) | |
| [15–25[ | 619 (11.6%) | 491 (12.3%) | 128 (9.7%) | 1.22 (0.99–1.52) | 1.13 (0.91–1.41) | |
| [25–50[ | 726 (13.7%) | 603 (15.1%) | 123 (9.3%) | 1.56 (1.26–1.94) | 1.58 (1.27–1.98) | |
| 50+ | 232 (4.4%) | 198 (5.0%) | 34 (2.6%) | 1.85 (1.29–2.74) | 1.88 (1.30–2.79) | |
| Mean age (95% CI) | 11.0 (10.8–11.2) | 13.3 (12.8–13.8) | 10.2 (9.6–10.9) | <0.001 | ||
| Symptoms n (%) | ||||||
| Fever | 4961 (93.3%) | 3775 (94.5%) | 1186 (89.7%) | <0.001 | 1.99 (1.58–2.48) | 2.08 (1.65–2.62) |
| Cough | 4484 (84.4%) | 3315 (83.0%) | 1169 (88.4%) | <0.001 | 0.64 (0.53–0.77) | 0.67 (0.55–0.81) |
| Rhinorrhea | 2395 (45.1%) | 1702 (42.6%) | 693 (52.4%) | <0.001 | 0.67 (0.60–0.76) | 0.71 (0.62–0.81) |
| Headache | 1106 (20.8%) | 850 (21.3%) | 256 (19.4%) | 0.146 | 1.13 (0.96–1.32) | - |
| Sore throat | 791 (14.9%) | 613 (15.4%) | 178 (13.5%) | 0.104 | 1.17 (0.98–1.40) | - |
| Myalgia | 874 (16.4%) | 680 (17.0%) | 194 (14.7%) | 0.050 | 1.19 (1.01–1.42) | - |
| Vomiting | 351 (6.6%) | 265 (6.6%) | 86 (6.5%) | 0.918 | 1.02 (0.80–1.32) | - |
| Dyspnea | 124 (2.3%) | 93 (2.3%) | 31 (2.3%) | 1.000 | 0.99 (0.67–1.52) | - |
| Arthralgia | 116 (2.2%) | 87 (2.2%) | 29 (2.2%) | 1.000 | 0.99 (0.66–1.54) | 0.64 (0.41–1.01) |
| Conjunctivitis | 20 (0.4%) | 16 (0.4%) | 4 (0.3%) | 0.797 | 1.33 (0.49–4.63) | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Touré, C.T.; Fall, A.; Andriamandimby, S.F.; Jallow, M.M.; Goudiaby, D.; Kiori, D.; Sy, S.; Diaw, Y.; Ndiaye, K.N.; Mbaye, F.; et al. Epidemiology and Molecular Analyses of Influenza B Viruses in Senegal from 2010 to 2019. Viruses 2022, 14, 1063. https://doi.org/10.3390/v14051063
Touré CT, Fall A, Andriamandimby SF, Jallow MM, Goudiaby D, Kiori D, Sy S, Diaw Y, Ndiaye KN, Mbaye F, et al. Epidemiology and Molecular Analyses of Influenza B Viruses in Senegal from 2010 to 2019. Viruses. 2022; 14(5):1063. https://doi.org/10.3390/v14051063
Chicago/Turabian StyleTouré, Cheikh Talibouya, Amary Fall, Soa Fy Andriamandimby, Mamadou Malado Jallow, Deborah Goudiaby, Davy Kiori, Sara Sy, Yague Diaw, Koba Ndiende Ndiaye, Fatimata Mbaye, and et al. 2022. "Epidemiology and Molecular Analyses of Influenza B Viruses in Senegal from 2010 to 2019" Viruses 14, no. 5: 1063. https://doi.org/10.3390/v14051063
APA StyleTouré, C. T., Fall, A., Andriamandimby, S. F., Jallow, M. M., Goudiaby, D., Kiori, D., Sy, S., Diaw, Y., Ndiaye, K. N., Mbaye, F., Niang, M. N., Heraud, J. M., & Dia, N. (2022). Epidemiology and Molecular Analyses of Influenza B Viruses in Senegal from 2010 to 2019. Viruses, 14(5), 1063. https://doi.org/10.3390/v14051063