
Transduction of HEK293 Cells with BacMam Baculovirus Is an Efficient System for the Production of HIV-1 Virus-like Particles

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines, Media, and Culture Conditions
2.2. BacMam Transfer Plasmid Construction
2.3. Generation of BacMam Working Stock and Titration
2.4. Butyric Acid Toxicity Assays
2.5. VLP Production in HEK293 Cells
2.6. D-Optimal Design
2.7. Statistical Analysis
2.8. Flow Cytometry
2.9. Spectrofluorometry
2.10. Nanoparticle Tracking Analysis
2.11. Super-Resolution Confocal Microscopy
2.12. Electron Microscopy
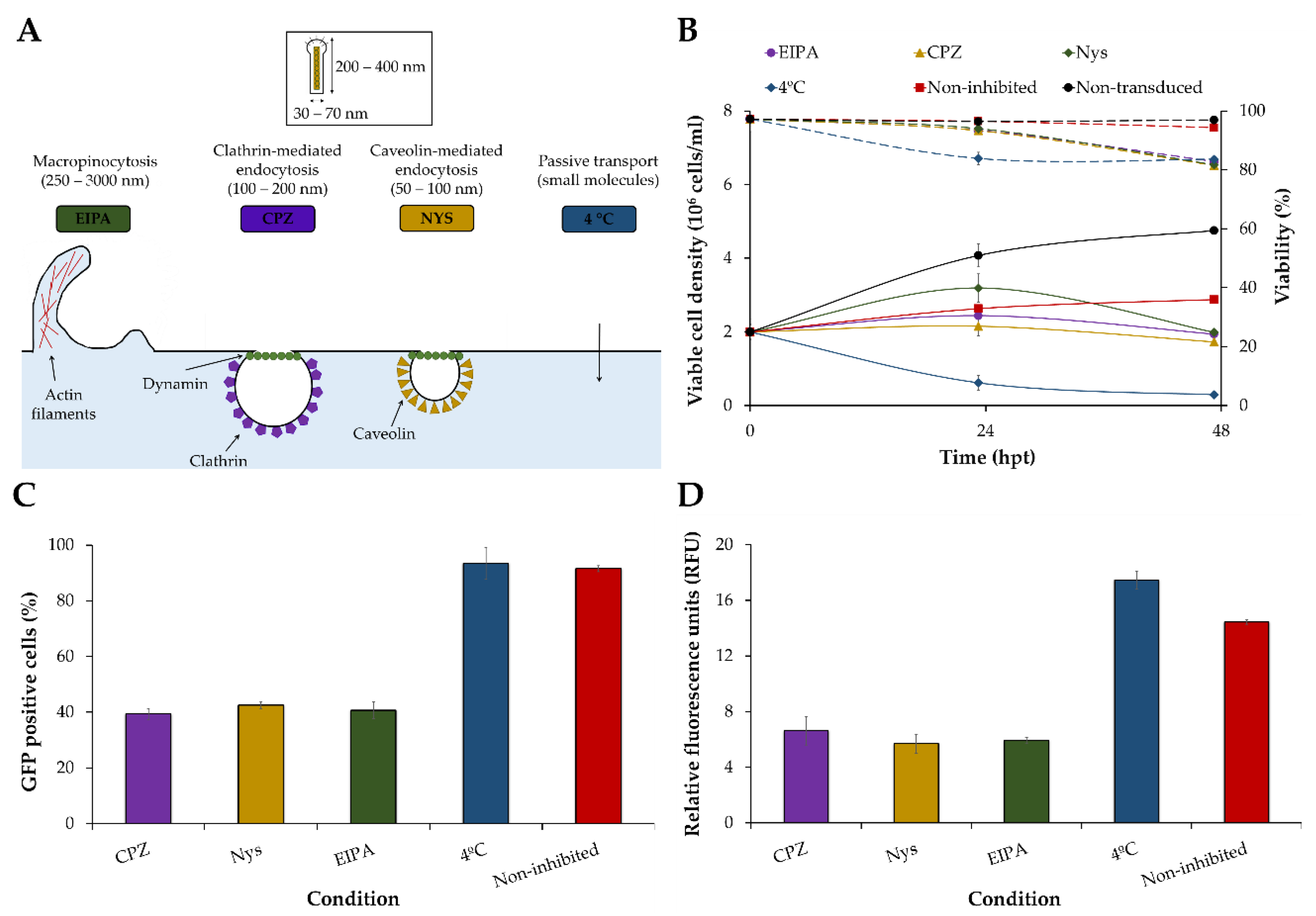
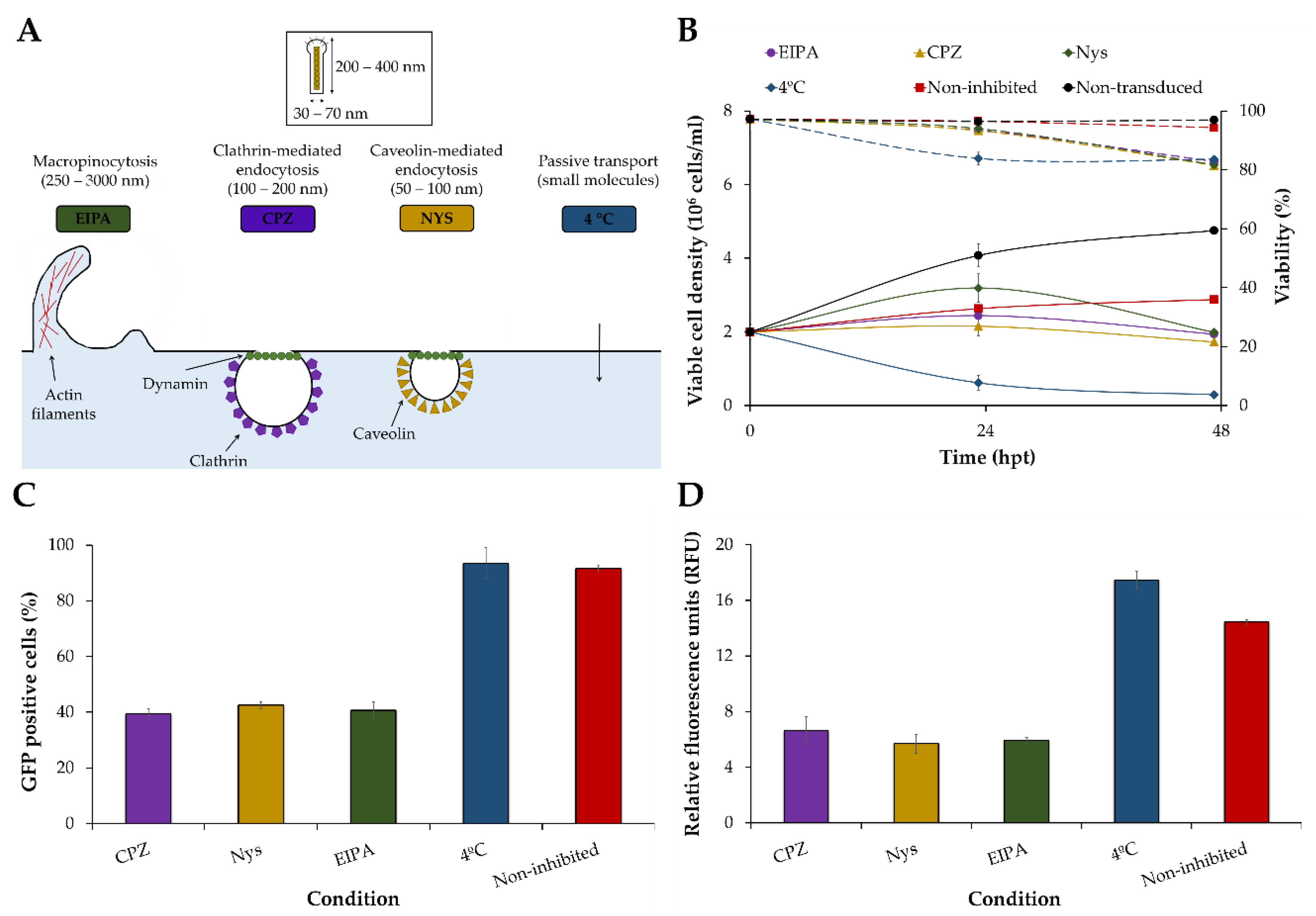
2.13. Inhibition of BacMam Baculovirus Uptake Pathways
3. Results
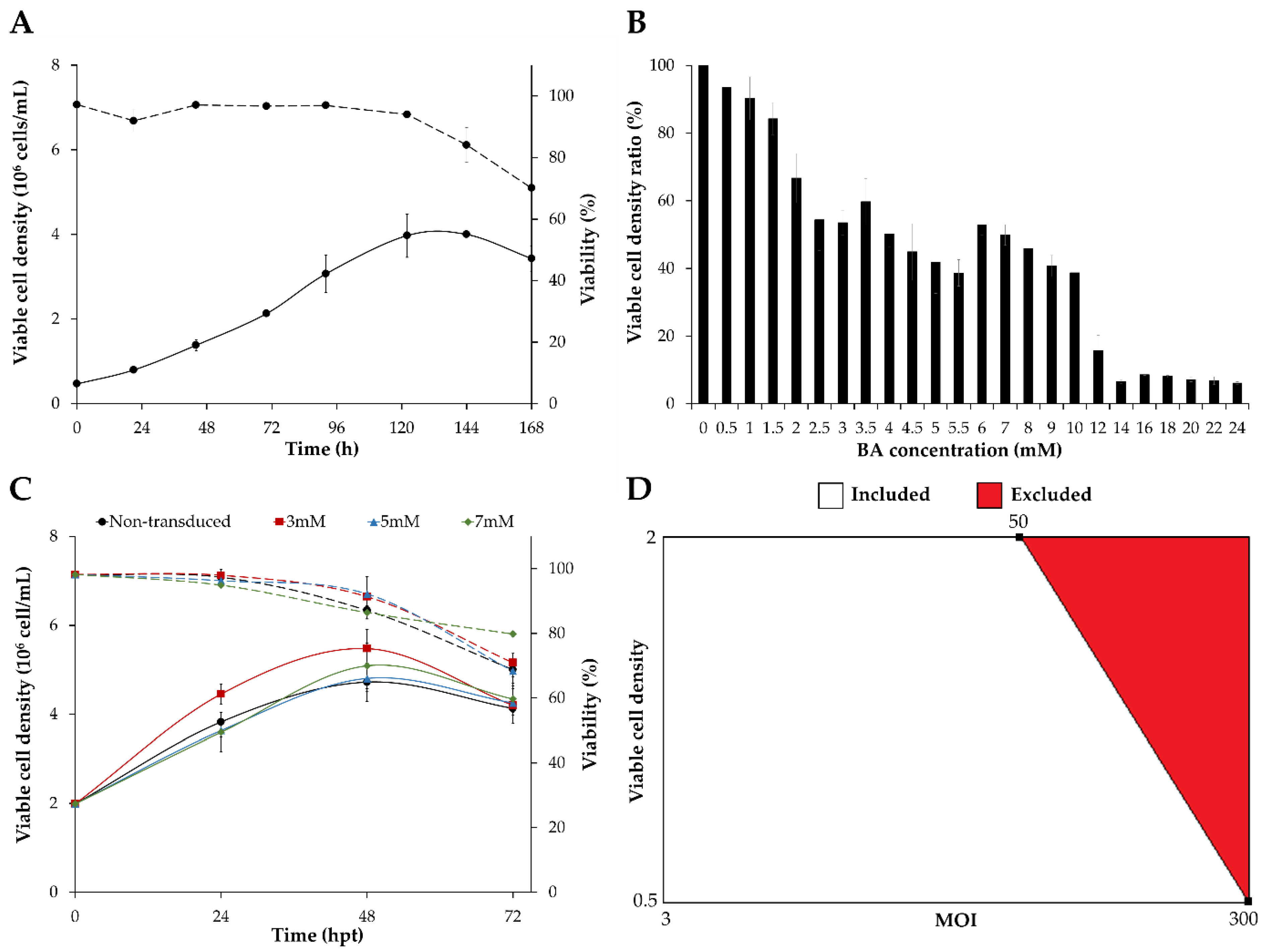
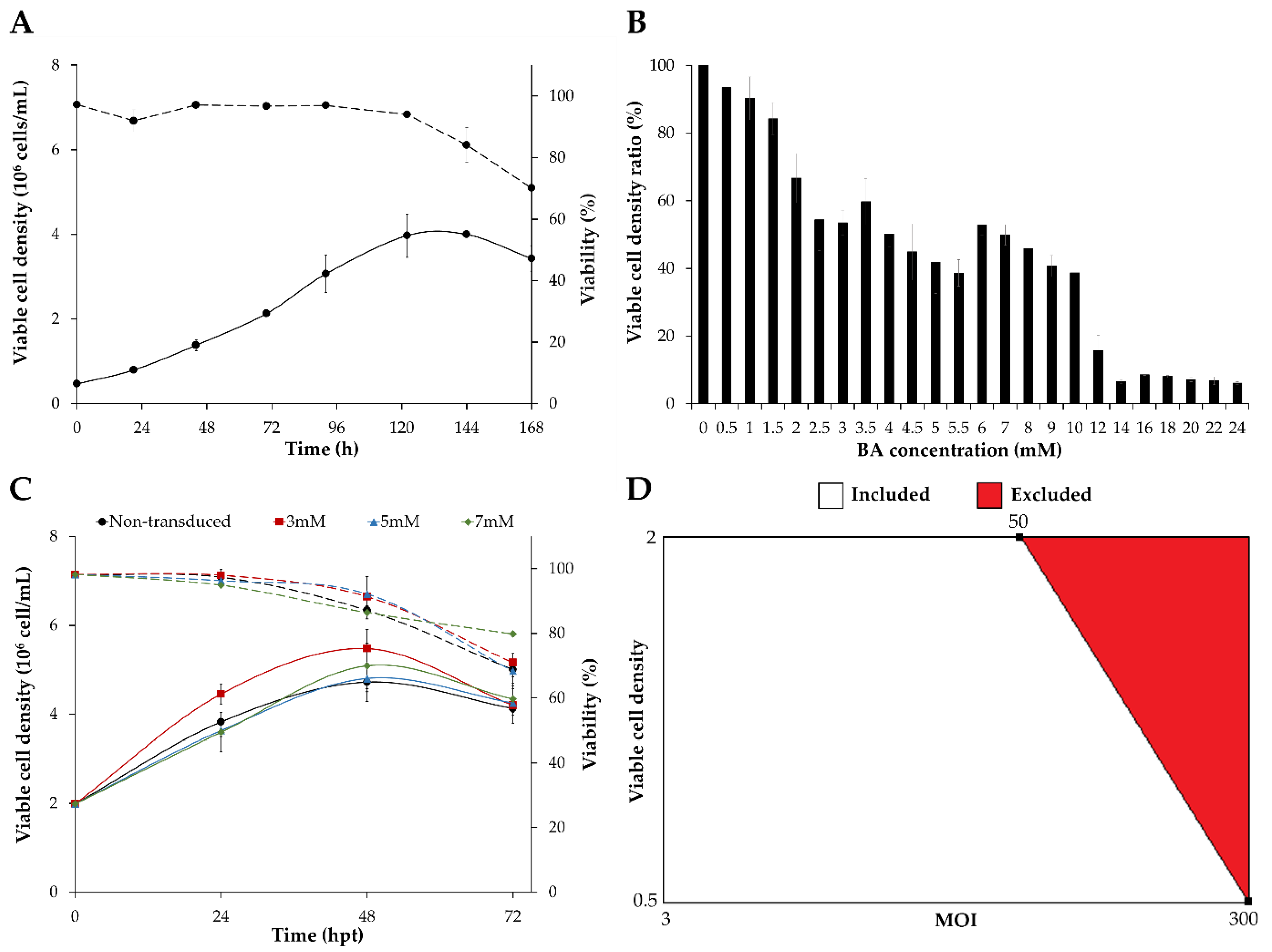
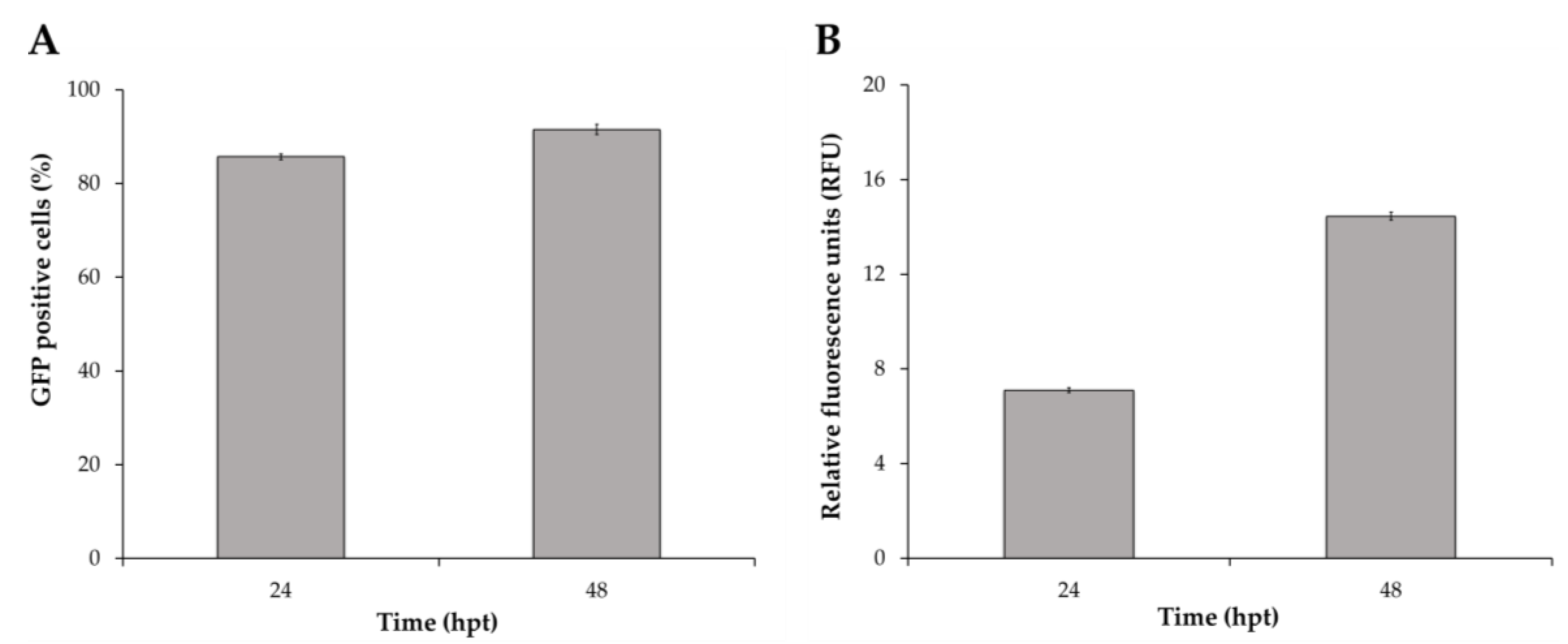
3.1. Screening of Transduction Conditions
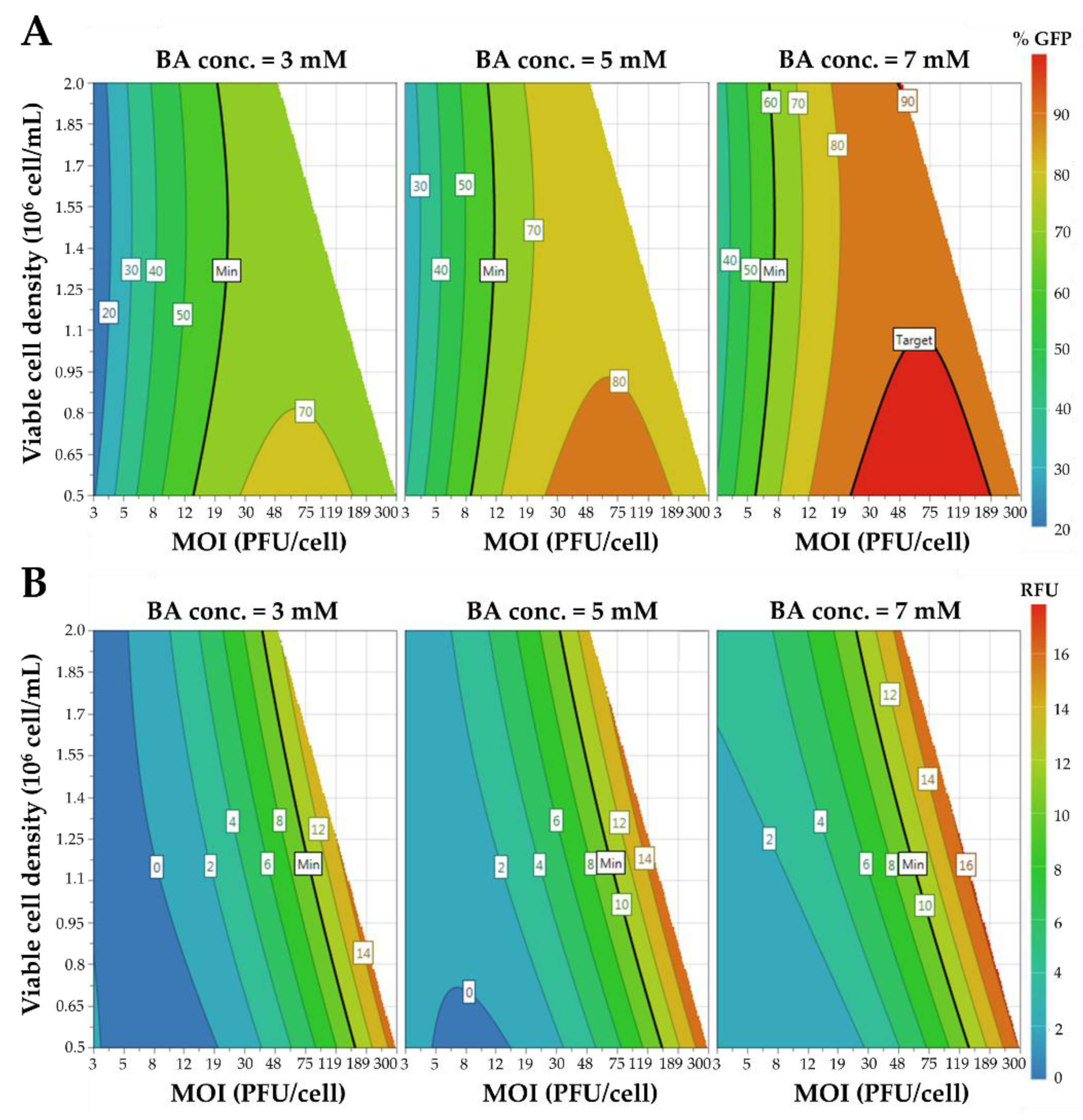
3.2. Optimization of HEK293 Cell Transduction by Design of Experiments (DoE)
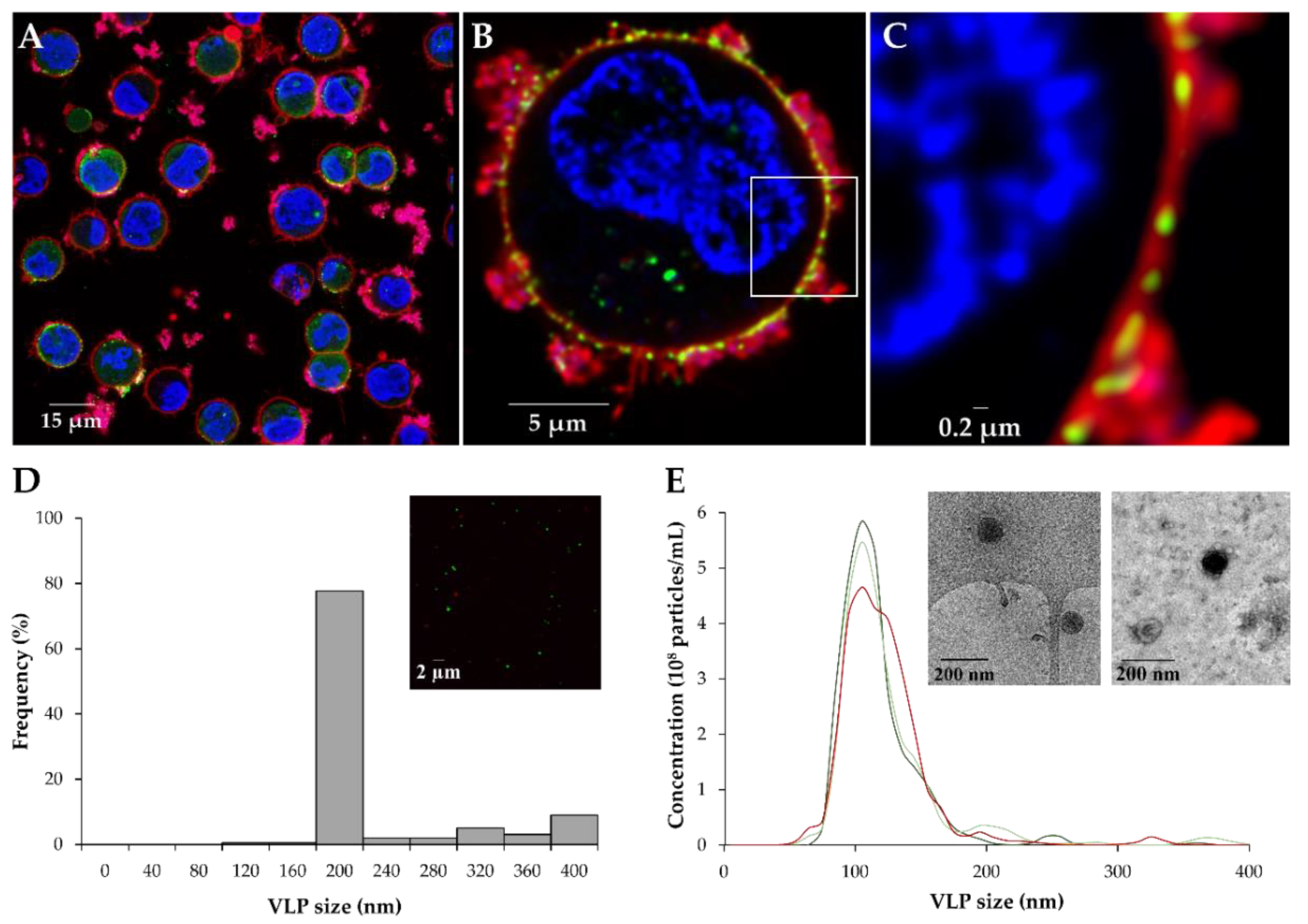
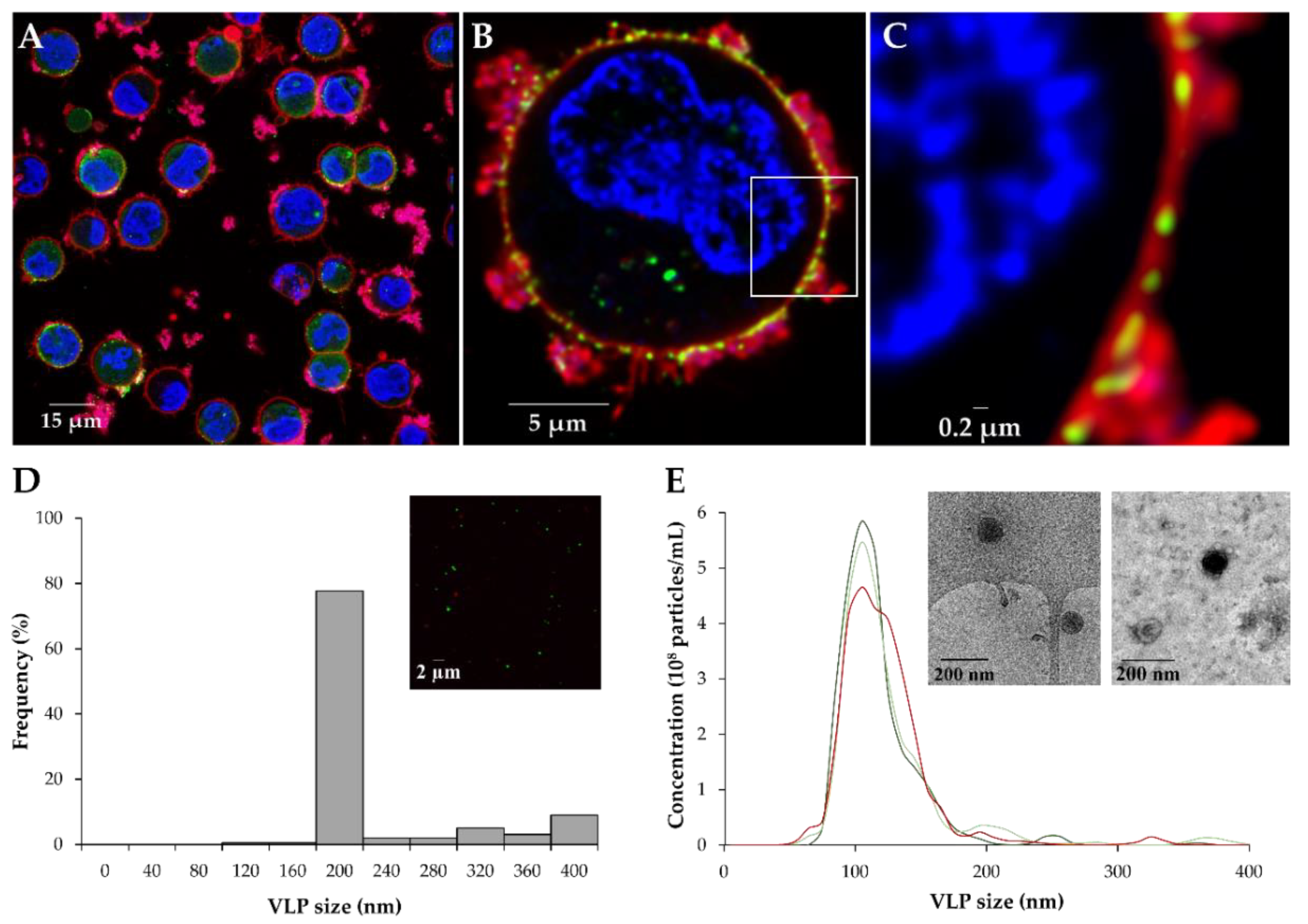
3.3. Characterization of VLP Production with the BacMam Expression System
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Nooraei, S.; Bahrulolum, H.; Hoseini, Z.S.; Katalani, C.; Hajizade, A.; Easton, A.J.; Ahmadian, G. Virus-like particles: Preparation, immunogenicity and their roles as nanovaccines and drug nanocarriers. J. Nanobiotechnol. 2021, 19, 59. [Google Scholar] [CrossRef] [PubMed]
- Qian, C.; Liu, X.; Xu, Q.; Wang, Z.; Chen, J.; Li, T.; Zheng, Q.; Yu, H.; Gu, Y.; Li, S.; et al. Recent progress on the versatility of virus-like particles. Vaccines 2020, 8, 139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, P.; Narayanan, E.; Liu, Q.; Tsybovsky, Y.; Boswell, K.; Ding, S.; Hu, Z.; Follmann, D.; Lin, Y.; Miao, H.; et al. A multiclade env-gag VLP mRNA vaccine elicits tier-2 HIV-1-neutralizing antibodies and reduces the risk of heterologous SHIV infection in macaques. Nat. Med. 2021, 27, 2234–2245. [Google Scholar] [CrossRef] [PubMed]
- Walpita, P.; Cong, Y.; Jahrling, P.B.; Rojas, O.; Postnikova, E.; Yu, S.; Johns, L.; Holbrook, M.R. A VLP-based vaccine provides complete protection against Nipah virus challenge following multiple-dose or single-dose vaccination schedules in a hamster model. npj Vaccines 2017, 2, 21. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, I.C.; Ipekoglu, E.M.; Bulbul, A.; Turay, N.; Yildirim, M.; Evcili, I.; Yilmaz, N.S.; Guvencli, N.; Aydin, Y.; Gungor, B.; et al. Development and preclinical evaluation of virus-like particle vaccine against COVID-19 infection. Allergy Eur. J. Allergy Clin. Immunol. 2021, 77, 258–270. [Google Scholar] [CrossRef] [PubMed]
- Deo, V.K.; Kato, T.; Park, E.Y. Chimeric Virus-Like Particles Made Using GAG and M1 Capsid Proteins Providing Dual Drug Delivery and Vaccination Platform. Mol. Pharm. 2015, 12, 839–845. [Google Scholar] [CrossRef]
- Fontana, D.; Garay, E.; Cevera, L.; Kratje, R.; Prieto, C.; Gòdia, F. Chimeric vlps based on hiv-1 gag and a fusion rabies glycoprotein induce specific antibodies against rabies and foot-and-mouth disease virus. Vaccines 2021, 9, 251. [Google Scholar] [CrossRef]
- Fernandes, F.; Teixeira, A.P.; Carinhas, N.; Carrondo, M.J.T.; Alves, P.M. Insect cells as a production platform of complex virus-like particles. Expert Rev. Vaccines 2013, 12, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Puente-Massaguer, E.; Lecina, M.; Gòdia, F. Application of advanced quantification techniques in nanoparticle-based vaccine development with the Sf9 cell baculovirus expression system. Vaccine 2020, 38, 1849–1859. [Google Scholar] [CrossRef] [PubMed]
- Puente-Massaguer, E.; Gòdia, F.; Lecina, M. Development of a non-viral platform for rapid virus-like particle production in Sf9 cells. J. Biotechnol. 2020, 322, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Puente-Massaguer, E.; Grau-Garcia, P.; Strobl, F.; Grabherr, R.; Striedner, G.; Lecina, M.; Gòdia, F. Accelerating HIV-1 VLP production using stable High Five insect cell pools. Biotechnol. J. 2021, 16, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Puente-Massaguer, E.; Grau-Garcia, P.; Strobl, F.; Grabherr, R.; Striedner, G.; Lecina, M.; Gòdia, F. Stable Sf9 cell pools as a system for rapid HIV-1 virus-like particle production. J. Chem. Technol. Biotechnol. 2021, 96, 3388–3397. [Google Scholar] [CrossRef]
- Lavado-García, J.; Cervera, L.; Gòdia, F. An Alternative Perfusion Approach for the Intensification of Virus-Like Particle Production in HEK293 Cultures. Front. Bioeng. Biotechnol. 2020, 8, 617. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.C.; Lu, H.R.; Ross, T.M. Baculovirus-produced influenza virus-like particles in mammalian cells protect mice from lethal influenza challenge. Viral Immunol. 2011, 24, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Boyce, F.M.; Bucher, N.L.R. Baculovirus-mediated gene transfer into mammalian cells. Proc. Natl. Acad. Sci. USA 1996, 93, 2348–2352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Targovnik, A.M.; Simonin, J.A.; Mc Callum, G.J.; Smith, I.; Cuccovia Warlet, F.U.; Nugnes, M.V.; Miranda, M.V.; Belaich, M.N. Solutions against emerging infectious and noninfectious human diseases through the application of baculovirus technologies. Appl. Microbiol. Biotechnol. 2021, 105, 8195–8226. [Google Scholar] [CrossRef]
- Cervera, L.; Gutiérrez-Granados, S.; Martínez, M.; Blanco, J.; Gòdia, F.; Segura, M.M. Generation of HIV-1 Gag VLPs by transient transfection of HEK 293 suspension cell cultures using an optimized animal-derived component free medium. J. Biotechnol. 2013, 166, 152–165. [Google Scholar] [CrossRef]
- Hermida-Matsumoto, L.; Resh, M.D. Localization of Human Immunodeficiency Virus Type 1 Gag and Env at the Plasma Membrane by Confocal Imaging. J. Virol. 2000, 74, 8670–8679. [Google Scholar] [CrossRef] [Green Version]
- Puente-Massaguer, E.; Lecina, M.; Gòdia, F. Integrating nanoparticle quantification and statistical design of experiments for efficient HIV-1 virus-like particle production in High Five cells. Appl. Microbiol. Biotechnol. 2020, 104, 1569–1582. [Google Scholar] [CrossRef]
- Puente-Massaguer, E.; Strobl, F.; Grabherr, R.; Striedner, G.; Lecina, M.; Gòdia, F. PEI-mediated transient transfection of high five cells at bioreactor scale for HIV-1 VLP production. Nanomaterials 2020, 10, 1580. [Google Scholar] [CrossRef]
- González-Domínguez, I.; Puente-Massaguer, E.; Cervera, L.; Gòdia, F. Quantification of the HIV-1 virus-like particle production process by super-resolution imaging: From VLP budding to nanoparticle analysis. Biotechnol. Bioeng. 2020, 117, 1929–1945. [Google Scholar] [CrossRef] [PubMed]
- Puente-Massaguer, E.; Saccardo, P.; Ferrer-Miralles, N.; Lecina, M.; Gòdia, F. Coupling Microscopy and Flow Cytometry for a Comprehensive Characterization of Nanoparticle Production in Insect Cells. Cytom. Part A 2020, 97, 921–932. [Google Scholar] [CrossRef] [PubMed]
- González-Domínguez, I.; Grimaldi, N.; Cervera, L.; Ventosa, N.; Gòdia, F. Impact of physicochemical properties of DNA/PEI complexes on transient transfection of mammalian cells. N. Biotechnol. 2019, 49, 88–97. [Google Scholar] [CrossRef] [Green Version]
- Kost, T.A.; Condreay, J.P.; Jarvis, D.L. Baculovirus as versatile vectors for protein expression in insect and mammalian cells. Nat. Biotechnol. 2005, 23, 567–575. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.S.; Ho, Y.C.; Wang, K.C.; Hu, Y.C. Investigation of optimal transduction conditions for baculovirus-mediated gene delivery into mammalian cells. Biotechnol. Bioeng. 2004, 88, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Li, Y.; Ning, Y.J.; Deng, F.; Vlak, J.M.; Hu, Z.; Wang, H.; Wang, M. The Major Hurdle for Effective Baculovirus Transduction into Mammalian Cells Is Passing Early Endosomes. J. Virol. 2019, 93, e00709-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volkman, L.E.; Goldsmith, P.A. In vitro survey of Autographa californica nuclear polyhedrosis virus interaction with nontarget vertebrate host cells. Appl. Environ. Microbiol. 1983, 45, 1085–1093. [Google Scholar] [CrossRef] [Green Version]
- Argilaguet, J.M.; Pérez-Martín, E.; López, S.; Goethe, M.; Escribano, J.M.; Giesow, K.; Keil, G.M.; Rodríguez, F. BacMam immunization partially protects pigs against sublethal challenge with African swine fever virus. Antiviral Res. 2013, 98, 61–65. [Google Scholar] [CrossRef]
- Dukkipati, A.; Park, H.H.; Waghray, D.; Fischer, S.; Garcia, K.C. BacMam system for high-level expression of recombinant soluble and membrane glycoproteins for structural studies. Protein Expr. Purif. 2008, 62, 160–170. [Google Scholar] [CrossRef] [Green Version]
- Jardin, B.A.; Zhao, Y.; Selvaraj, M.; Montes, J.; Tran, R.; Prakash, S.; Elias, C.B. Expression of SEAP (secreted alkaline phosphatase) by baculovirus mediated transduction of HEK 293 cells in a hollow fiber bioreactor system. J. Biotechnol. 2008, 135, 272–280. [Google Scholar] [CrossRef]
- Scott, M.J.; Modha, S.S.; Rhodes, A.D.; Broadway, N.M.; Hardwicke, P.I.; Zhao, H.J.; Kennedy-Wilson, K.M.; Sweitzer, S.M.; Martin, S.L. Efficient expression of secreted proteases via recombinant BacMam virus. Protein Expr. Purif. 2007, 52, 104–116. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Chen, P.H.; He, X. Expression and purification of class 7 semaphorin and its Plexinc1 receptor using baculovirus-mediated mammalian cell gene transduction. In Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2017; Volume 1493, pp. 41–56. [Google Scholar]
- Thompson, C.M.; Petiot, E.; Mullick, A.; Aucoin, M.G.; Henry, O.; Kamen, A. Critical assessment of influenza VLP production in Sf9 and HEK293 expression systems. BMC Biotechnol. 2015, 15, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Lavado-García, J.; Jorge, I.; Boix-Besora, A.; Vázquez, J.; Gòdia, F.; Cervera, L. Characterization of HIV-1 virus-like particles and determination of Gag stoichiometry for different production platforms. Biotechnol. Bioeng. 2021, 118, 2660–2675. [Google Scholar] [CrossRef] [PubMed]
- González-Domínguez, I.; Puente-Massaguer, E.; Cervera, L.; Gòdia, F. Quality Assessment of Virus-Like Particles at Single Particle Level: A Comparative Study. Viruses 2020, 12, 223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altmann, F.; Staudacher, E.; Wilson, I.B.H.; März, L. Insect cells as hosts for the expression of recombinant glycoproteins. Glycoconj. J. 1999, 16, 109–123. [Google Scholar] [CrossRef]
- Chang, G.R.L.; Lai, S.Y.; Chang, P.C.; Wang, M.Y. Production of immunogenic one-component avian H7-subtype influenza virus-like particles. Process Biochem. 2011, 46, 1292–1298. [Google Scholar] [CrossRef]
- Blissard, G.W.; Theilmann, D.A. Baculovirus Entry and Egress from Insect Cells. Annu. Rev. Virol. 2018, 5, 113–139. [Google Scholar] [CrossRef]
- Long, G.; Pan, X.; Kormelink, R.; Vlak, J.M. Functional Entry of Baculovirus into Insect and Mammalian Cells Is Dependent on Clathrin-Mediated Endocytosis. J. Virol. 2006, 80, 8830–8833. [Google Scholar] [CrossRef] [Green Version]
- Kataoka, C.; Kaname, Y.; Taguwa, S.; Abe, T.; Fukuhara, T.; Tani, H.; Moriishi, K.; Matsuura, Y. Baculovirus GP64-Mediated Entry into Mammalian Cells. J. Virol. 2012, 86, 2610–2620. [Google Scholar] [CrossRef] [Green Version]
- Lavado-García, J.; Díaz-Maneh, A.; Canal-Paulí, N.; Pérez-Rubio, P.; Gòdia, F.; Cervera, L. Metabolic engineering of HEK293 cells to improve transient transfection and cell budding of HIV-1 virus-like particles. Biotechnol. Bioeng. 2021, 118, 1649–1663. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Independent Variables | Coding Levels | ||
|---|---|---|---|
| −1 | 0 | 1 | |
| MOI (PFU/cell) | 3 | 30 | 300 |
| Viable cell density (106 cell/mL) | 0.5 | 1.25 | 2 |
| Butyric acid concentration (mM) | 3 | 5 | 7 |
| Exp. No. | MOI | VCD (106 cell/mL) | Butyric Acid (mM) | Cell Transduction Efficiency (%) | VLP Production (RFU) |
|---|---|---|---|---|---|
| 1 | 3 | 0.5 | 3 | 14.5 | 0.4 |
| 2 | 300 | 0.5 | 3 | 58.7 | 14.6 |
| 3 | 3 | 0.5 | 7 | 43.7 | 0.9 |
| 4 | 300 | 0.5 | 7 | 80.4 | 15.6 |
| 5 | 3 | 2 | 7 | 40.0 | 1.6 |
| 6 | 50 | 2 | 3 | 65.1 | 10.6 |
| 7 | 50 | 2 | 7 | 88.6 | 15.4 |
| 8 | 3 | 0.5 | 5.7 | 32.2 | 0.8 |
| 9 | 3 | 2 | 4.3 | 24.5 | 0.7 |
| 10 | 3 | 1 | 3 | 12.9 | 0.1 |
| 11 | 3 | 1.5 | 3 | 9.4 | 0.1 |
| 12 | 64.6 | 0.5 | 3 | 82.5 | 3.9 |
| 13 | 64.6 | 0.5 | 7 | 91.7 | 6.1 |
| 14 | 7.7 | 2 | 3 | 38.0 | 1.5 |
| 15 | 30 | 0.5 | 5 | 86.8 | 2.5 |
| 16 | 19.1 | 1.25 | 7 | 77.9 | 4.6 |
| 17 | 122.5 | 1.25 | 5 | 83.2 | 17.3 |
| 18 | 19.1 | 1.25 | 5 | 67.3 | 3.3 |
| 19 | 19.1 | 1.25 | 5 | 68.7 | 3.2 |
| 20 | 19.1 | 1.25 | 5 | 67.7 | 2.8 |
| Cell transduction efficiency function—Parameters | Coefficient | Standard error | p-value | ||
| Constant | 74.4 | 1.9 | <0.01 | ||
| MOI | 20.9 | 1.8 | <0.01 | ||
| Butyric acid concentration | 11.2 | 1.3 | <0.01 | ||
| MOI2 | −30.9 | 2.7 | <0.01 | ||
| VLP production function—Parameters | Coefficient | Standard error | p-value | ||
| Constant | 5.7 | 0.3 | <0.01 | ||
| MOI | 11.5 | 0.5 | <0.01 | ||
| Viable cell density | 4.0 | 0.3 | <0.01 | ||
| Butyric acid concentration | 1.1 | 0.3 | <0.01 | ||
| MOI2 | 6.6 | 0.6 | <0.01 | ||
| MOI × Viable cell concentration | 4.1 | 0.5 | <0.01 | ||
| System | Cell Line | Titer | PTMs | Scalability | Cost | Time | BV Contamination |
|---|---|---|---|---|---|---|---|
| BacMam | HEK293 | ++ | ++++ | ++ | ++ | ++ | + |
| SGE | ++ | ++++ | +++ | ++ | ++++ | − | |
| Sf9 | + | ++ | ++++ | + | +++ | − | |
| High Five | + | ++ | ++++ | + | +++ | − | |
| TGE | HEK293 | ++ | ++++ | + | ++++ | + | − |
| Sf9 | ++ | ++ | + | +++ | + | − | |
| High Five | + | ++ | + | +++ | + | − | |
| BEVS | ++++ | ++ | +++ | + | + | +++ | |
| Sf9 | ++++ | ++ | +++ | + | + | ++++ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Puente-Massaguer, E.; Cajamarca-Berrezueta, B.; Volart, A.; González-Domínguez, I.; Gòdia, F. Transduction of HEK293 Cells with BacMam Baculovirus Is an Efficient System for the Production of HIV-1 Virus-like Particles. Viruses 2022, 14, 636. https://doi.org/10.3390/v14030636
Puente-Massaguer E, Cajamarca-Berrezueta B, Volart A, González-Domínguez I, Gòdia F. Transduction of HEK293 Cells with BacMam Baculovirus Is an Efficient System for the Production of HIV-1 Virus-like Particles. Viruses. 2022; 14(3):636. https://doi.org/10.3390/v14030636
Chicago/Turabian StylePuente-Massaguer, Eduard, Byron Cajamarca-Berrezueta, Aleix Volart, Irene González-Domínguez, and Francesc Gòdia. 2022. "Transduction of HEK293 Cells with BacMam Baculovirus Is an Efficient System for the Production of HIV-1 Virus-like Particles" Viruses 14, no. 3: 636. https://doi.org/10.3390/v14030636
APA StylePuente-Massaguer, E., Cajamarca-Berrezueta, B., Volart, A., González-Domínguez, I., & Gòdia, F. (2022). Transduction of HEK293 Cells with BacMam Baculovirus Is an Efficient System for the Production of HIV-1 Virus-like Particles. Viruses, 14(3), 636. https://doi.org/10.3390/v14030636