
Multiple Neuraminidase Containing Influenza Virus-like Particle Vaccines Protect Mice from Avian and Human Influenza Virus Infection

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice, Cells, and Viruses
2.2. Generation of Recombinant Baculovirus and Virus-Like Particles
2.3. Characterization of VLPs
2.4. Immunization and Challenge Infection
2.5. Antibody Response, Virus Titer, and Cytokine Response
2.6. Microneutralization Activity
2.7. Neuraminidase Inhibition (NAI) Analysis
2.8. Flow Cytometry
2.9. Statistical Analysis
3. Results
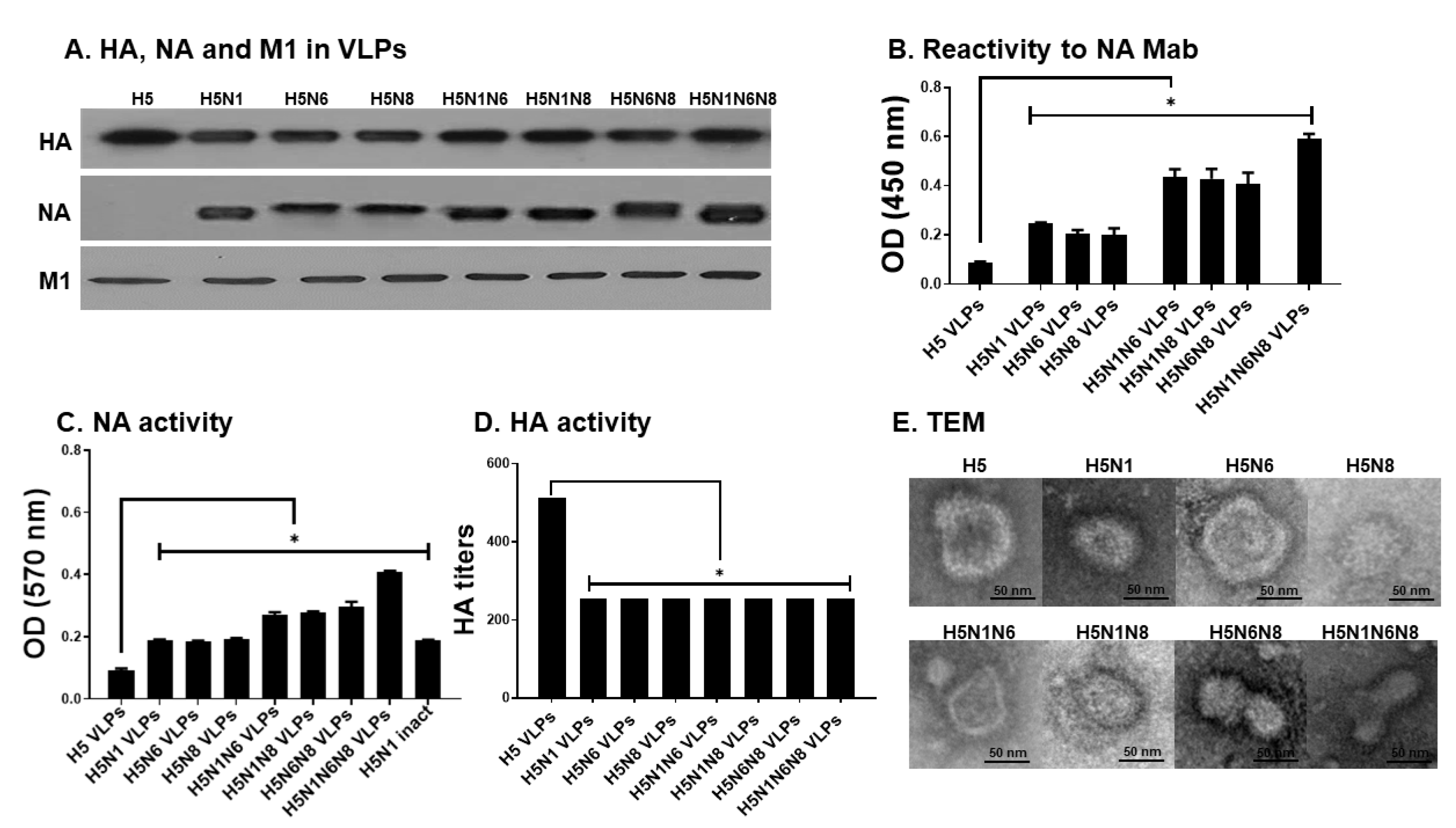
3.1. Production of Influenza VLPs Containing Influenza HA, NA, and M1
3.2. VLPs Vaccination Induced H5N1 or H3N2 Virus-Specific Antibody Responses in Sera and Lungs
3.3. VLPs Vaccination Induced H5N1 or H3N2 Virus-Specific Antibody Responses in Sera and Lungs
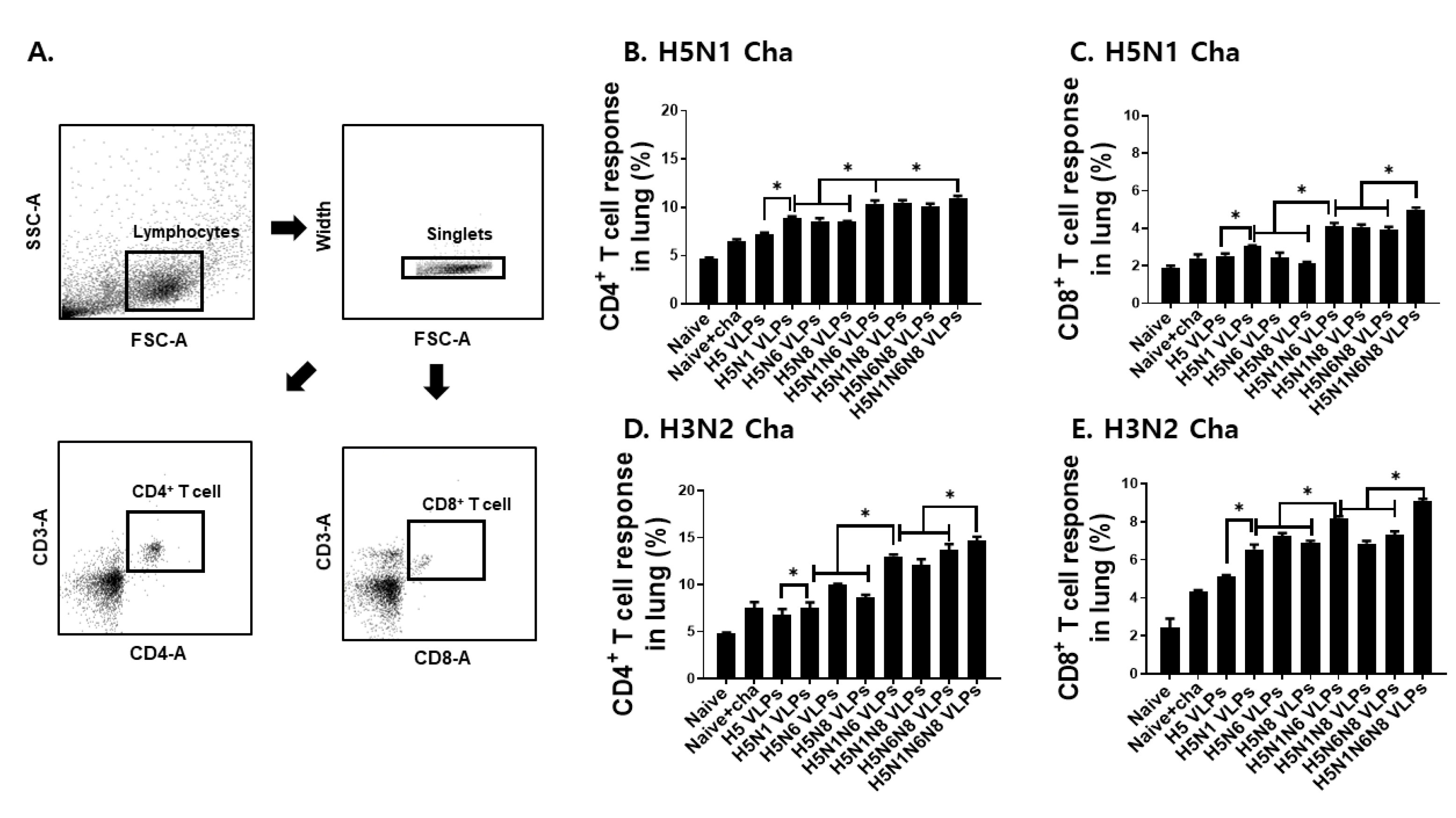
3.4. VLP Vaccines Elicited H5N1 or H3N2 Virus-Specific CD4+ and CD8+ T Cell Immune Responses in the Lung
3.5. VLPs Vaccine Elicited H5N1 or H3N2 Viruses-Specific Germinal Center-Like Cell and B Cells Immune Responses in the Lung
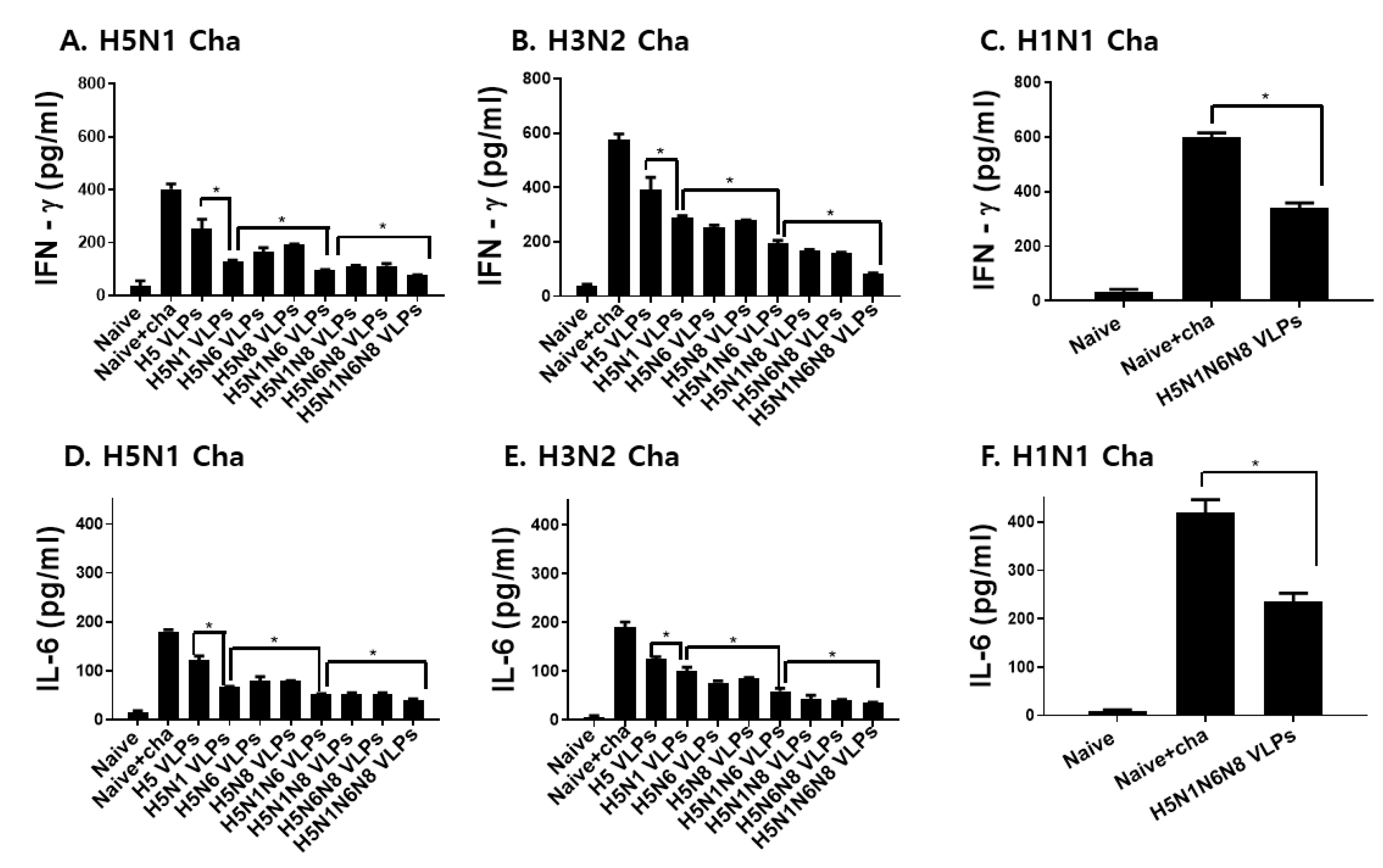
3.6. VLPs Vaccination Significantly Reduced Inflammatory Cytokine Response and Lung Virus Titers
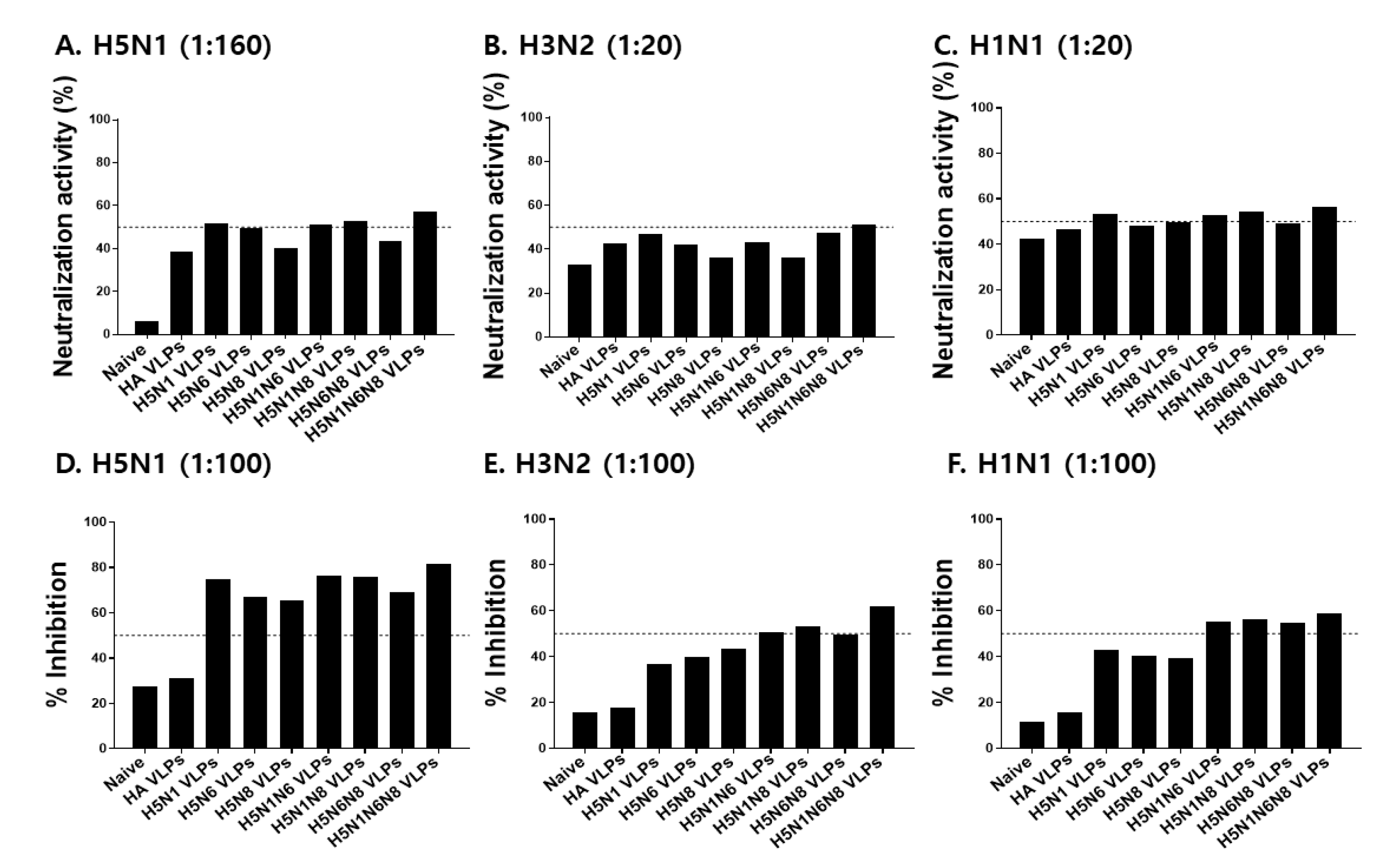
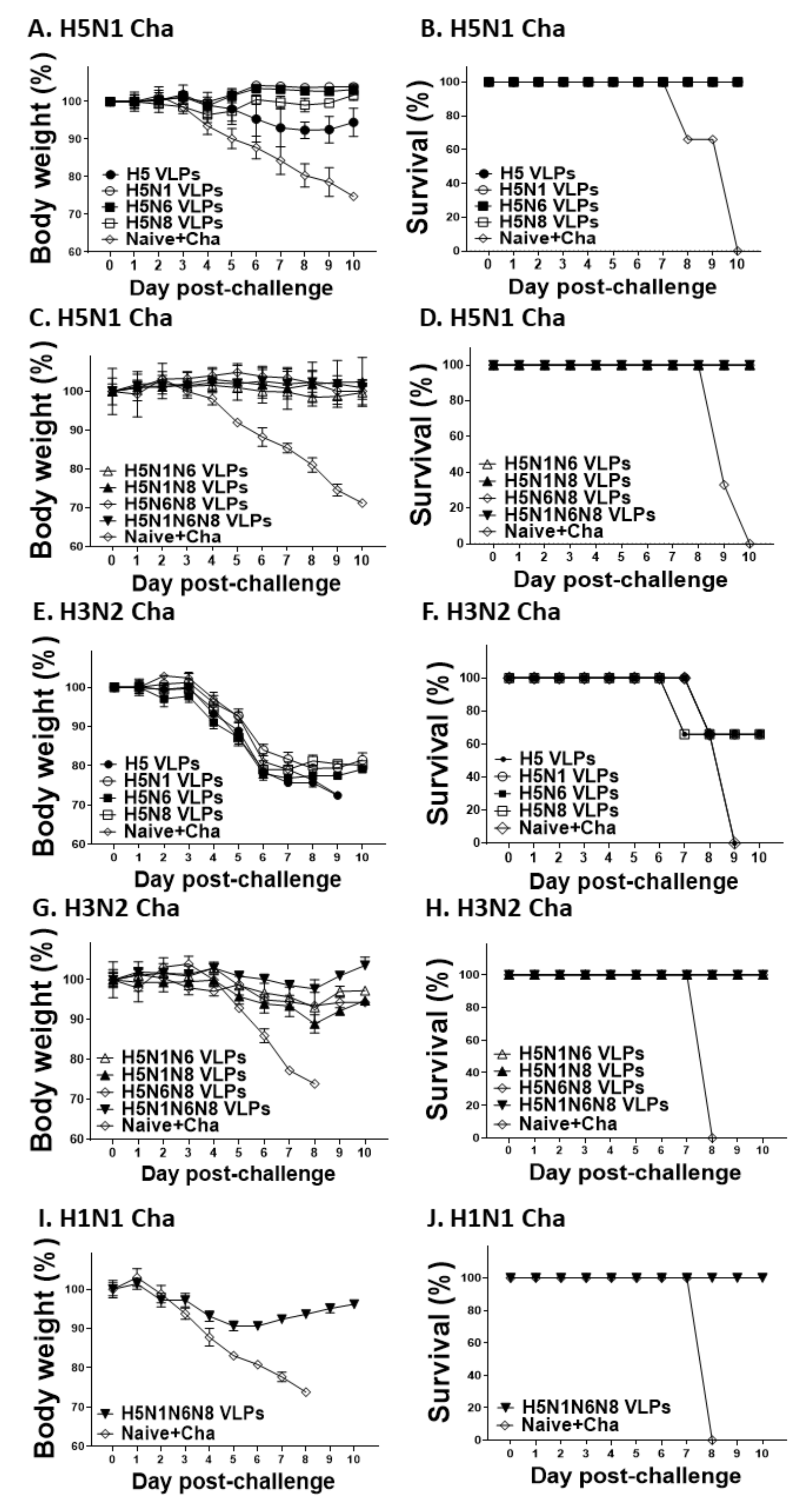
3.7. VLPs Vaccine Efficacy against H5N1, H3N2, and H1N1 Virus Challenge Infections
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Short, K.; Richard, M.; Verhagen, J.H.; van Riel, D.; Schrauwen, E.J.; Brand, J.M.V.D.; Mänz, B.; Bodewes, R.; Herfst, S. One health, multiple challenges: The inter-species transmission of influenza A virus. One Health 2015, 1, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, N.P.A.S.; Mueller, J. Updating the Accounts: Global Mortality of the 1918–1920 “Spanish” Influenza Pandemic. Bull. Hist. Med. 2002, 76, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Reid, A.; Taubenberger, J.K.; Fanning, T.G. Evidence of an absence: The genetic origins of the 1918 pandemic influenza virus. Nat. Rev. Genet. 2004, 2, 909–914. [Google Scholar] [CrossRef]
- Taubenberger, J.K.; Morens, D.M. 1918 Influenza: The mother of all pandemics. Rev. Biomed. 2006, 17, 69–79. [Google Scholar] [CrossRef]
- World Organisation for Animal Health. Available online: https://www.oie.int/en/disease/avian-influenza (accessed on 12 January 2022).
- Lai, S.; Qin, Y.; Cowling, B.J.; Ren, X.; Wardrop, N.A.; Gilbert, M.; Tsang, T.K.; Wu, P.; Feng, L.; Jiang, H.; et al. Global epidemiology of avian influenza A H5N1 virus infection in humans, 1997–2015: A systematic review of individual case data. Lancet Infect. Dis. 2016, 16, e108–e118. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.-H.; Lee, Y.-T.; Park, S.; Jung, Y.-J.; Lee, Y.; Ko, E.-J.; Kim, Y.-J.; Li, X.; Kang, S.-M. Neuraminidase expressing virus-like particle vaccine provides effective cross protection against influenza virus. Virology 2019, 535, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Stadlbauer, D.; Zhu, X.; McMahon, M.; Turner, J.S.; Wohlbold, T.J.; Schmitz, A.J.; Strohmeier, S.; Yu, W.; Nachbagauer, R.; Mudd, P.A.; et al. Broadly protective human antibodies that target the active site of influenza virus neuraminidase. Science 2019, 366, 499–504. [Google Scholar] [CrossRef]
- Heaton, N.S.; Sachs, D.; Chen, C.-J.; Hai, R.; Palese, P. Genome-wide mutagenesis of influenza virus reveals unique plasticity of the hemagglutinin and NS1 proteins. Proc. Natl. Acad. Sci. USA 2013, 110, 20248–20253. [Google Scholar] [CrossRef] [Green Version]
- Krammer, F.; Fouchier, R.A.; Eichelberger, M.C.; Webby, R.J.; Shaw-Saliba, K.; Wan, H.; Wilson, P.C.; Compans, R.W.; Skountzou, I.; Monto, A.S. NAction! How can neuraminidase-based immunity contribute to better influenza virus vaccines? MBio 2018, 9, e02332-17. [Google Scholar] [CrossRef] [Green Version]
- Sandbulte, M.; Westgeest, K.B.; Gao, J.; Xu, X.; Klimov, A.I.; Russell, C.; Burke, D.; Smith, D.; Fouchier, R.; Eichelberger, M.C. Discordant antigenic drift of neuraminidase and hemagglutinin in H1N1 and H3N2 influenza viruses. Proc. Natl. Acad. Sci. USA 2011, 108, 20748–20753. [Google Scholar] [CrossRef] [Green Version]
- Westgeest, K.; Russell, C.; Lin, X.; Spronken, M.; Bestebroer, T.; Bahl, J.; Van Beek, R.; Skepner, E.; Halpin, R.; De Jong, J.; et al. Genomewide Analysis of Reassortment and Evolution of Human Influenza A(H3N2) Viruses Circulating between 1968 and 2011. J. Virol. 2014, 88, 2844–2857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Job, E.R.; Ysenbaert, T.; Smet, A.; Christopoulou, I.; Strugnell, T.; Oloo, E.O.; Oomen, R.P.; Kleanthous, H.; Vogel, T.U.; Saelens, X. Broadened immunity against influenza by vaccination with computationally designed influenza virus N1 neuraminidase constructs. Vaccines 2018, 3, 55. [Google Scholar] [CrossRef]
- Johansson, B.E.; Brett, I.C. Changing perspective on immunization against influenza. Vaccine 2007, 25, 3062–3065. [Google Scholar] [CrossRef]
- Chackerian, B.; Peabody, D.S. Factors That Govern the Induction of Long-Lived Antibody Responses. Viruses 2020, 12, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quan, F.-S.; Kim, M.-C.; Lee, B.-J.; Song, J.-M.; Compans, R.; Kang, S.-M. Influenza M1 VLPs containing neuraminidase induce heterosubtypic cross-protection. Virology 2012, 430, 127–135. [Google Scholar] [CrossRef] [Green Version]
- Zhao, G.; Sun, S.; Du, L.; Xiao, W.; Ru, Z.; Kou, Z.; Guo, Y.; Yu, H.; Jiang, S.; Lone, Y.; et al. An H5N1 M2e-based multiple antigenic peptide vaccine confers heterosubtypic protection from lethal infection with pandemic 2009 H1N1 virus. Virol. J. 2010, 7, 151. [Google Scholar] [CrossRef] [Green Version]
- Ren, Z.; Ji, X.; Meng, L.; Wei, Y.; Wang, T.; Feng, N.; Zheng, X.; Wang, H.; Li, N.; Gao, X.; et al. H5N1 influenza virus-like particle vaccine protects mice from heterologous virus challenge better than whole inactivated virus. Virus Res. 2015, 200, 9–18. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention, CDC. Isolation of avian influenza A (H5N1) viruses from humans—Hong Kong, May–December. MMWR Morbid. Mortal. Wkly. Rep. 1997, 46, 1204–1207. [Google Scholar]
- Perrone, L.A.; Ahmad, A.; Veguilla, V.; Lu, X.; Smith, G.; Katz, J.M.; Pushko, P.; Tumpey, T.M. Intranasal Vaccination with 1918 Influenza Virus-Like Particles Protects Mice and Ferrets from Lethal 1918 and H5N1 Influenza Virus Challenge. J. Virol. 2009, 83, 5726–5734. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.; Ko, E.; Lee, Y.; Kim, K.; Kim, M.; Lee, Y.; Kang, S. Intranasal vaccination with M2e5x virus-like particles induces humoral and cellular immune responses conferring cross-protection against heterosubtypic influenza viruses. PLoS ONE 2018, 13, e0190868. [Google Scholar] [CrossRef] [Green Version]
- Quan, F.-S.; Vunnava, A.; Compans, R.W.; Kang, S.-M. Virus-Like Particle Vaccine Protects against 2009 H1N1 Pandemic Influenza Virus in Mice. PLoS ONE 2010, 5, e9161. [Google Scholar] [CrossRef] [PubMed]
- Quan, F.-S.; Huang, C.; Compans, R.W.; Kang, S.-M. Virus-Like Particle Vaccine Induces Protective Immunity against Homologous and Heterologous Strains of Influenza Virus. J. Virol. 2007, 81, 3514–3524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, H.-J.; Chu, K.-B.; Lee, D.-H.; Lee, S.-H.; Park, B.R.; Kim, M.-C.; Kang, S.-M.; Quan, F.-S. Influenza M2 virus-like particle vaccination enhances protection in combination with avian influenza HA VLPs. PLoS ONE 2019, 14, e0216871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, H.-J.; Chu, K.-B.; Yoon, K.-W.; Eom, G.-D.; Mao, J.; Kim, M.-J.; Lee, S.-H.; Moon, E.-K.; Quan, F.-S. Neuraminidase in Virus-like Particles Contributes to the Protection against High Dose of Avian Influenza Virus Challenge Infection. Pathogens 2021, 10, 1291. [Google Scholar] [CrossRef]
- Doyle, T.M.; Hashem, A.M.; Li, C.; Van Domselaar, G.; Larocque, L.; Wang, J.; Smith, D.; Cyr, T.; Farnsworth, A.; He, R.; et al. Universal anti-neuraminidase antibody inhibiting all influenza A subtypes. Antivir. Res. 2013, 100, 567–574. [Google Scholar] [CrossRef]
- Choi, H.-J.; Yoo, D.-G.; Bondy, B.J.; Quan, F.-S.; Compans, R.; Kang, S.-M.; Prausnitz, M.R. Stability of influenza vaccine coated onto microneedles. Biomaterials 2012, 33, 3756–3769. [Google Scholar] [CrossRef] [Green Version]
- Basak, S.; Kang, H.-J.; Lee, S.-H.; Chu, K.-B.; Moon, E.-K.; Quan, F.-S. Influenza vaccine efficacy induced by orally administered recombinant baculoviruses. PLoS ONE 2020, 15, e0233520. [Google Scholar] [CrossRef]
- Gross, F.L.; Bai, Y.; Jefferson, S.; Holiday, C.; Levine, M.Z. Measuring Influenza Neutralizing Antibody Responses to A(H3N2) Viruses in Human Sera by Microneutralization Assays Using MDCK-SIAT1 Cells. J. Vis. Exp. 2017, 129, 56448. [Google Scholar] [CrossRef]
- World Health Organization. Serological Diagnosis of Influenza by Microneutralization Assay; WHO Press: Geneva, Switzerland, 2015. [Google Scholar]
- Kang, H.; Chu, K.; Kim, M.; Lee, S.; Park, H.; Jin, H.; Moon, E.; Quan, F. Protective immunity induced by CpG ODN-adjuvanted virus-like particles containing Toxoplasma gondii proteins. Parasite Immunol. 2021, 43, e12799. [Google Scholar] [CrossRef]
- Zhang, R.; Chen, T.; Ou, X.; Liu, R.; Yang, Y.; Ye, W.; Chen, J.; Yao, D.; Sun, B.; Zhang, X.; et al. Clinical, epidemiological and virological characteristics of the first detected human case of avian influenza A(H5N6) virus. Infect. Genet. Evol. 2016, 40, 236–242. [Google Scholar] [CrossRef] [Green Version]
- Wei, S.-H.; Yang, J.-R.; Wu, H.-S.; Chang, M.-C.; Lin, J.-S.; Lin, C.-Y.; Liu, Y.-L.; Lo, Y.-C.; Yang, C.-H.; Chuang, J.-H.; et al. Human infection with avian influenza A H6N1 virus: An epidemiological analysis. Lancet Respir. Med. 2013, 1, 771–778. [Google Scholar] [CrossRef]
- Parry, J. H10N8 avian flu virus claims its first known human casualty. BMJ 2014, 348, g1360. [Google Scholar] [CrossRef]
- Angeletti, D.; Yewdell, J.W. Is it possible to develop a “universal” influenza virus vaccine? Outflanking antibody immunodominance on the road to universal influenza vaccination. Cold Spring Harbor Perspect. Biol. 2018, 10, a028852. [Google Scholar] [CrossRef] [PubMed]
- Schotsaert, M.; Ysenbaert, T.; Smet, A.; Schepens, B.; Vanderschaeghe, D.; Stegalkina, S.; Vogel, T.U.; Callewaert, N.; Fiers, W.; Saelens, X. Long-Lasting Cross-Protection Against Influenza A by Neuraminidase and M2e-based immunization strategies. Sci. Rep. 2016, 6, 24402. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Zhang, Z.; Liu, H.; Tian, M.; Zhu, X.; Zhang, Z.; Wang, W.; Zhou, X.; Zhang, F.; Ge, Q.; et al. B Cells Are the Dominant Antigen-Presenting Cells that Activate Naive CD4+ T Cells upon Immunization with a Virus-Derived Nanoparticle Antigen. Immunity 2018, 49, 695–708.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, H.-J.; Chu, K.-B.; Yoon, K.-W.; Eom, G.-D.; Mao, J.; Kim, M.-J.; Lee, S.-H.; Moon, E.-K.; Quan, F.-S. Multiple Neuraminidase Containing Influenza Virus-like Particle Vaccines Protect Mice from Avian and Human Influenza Virus Infection. Viruses 2022, 14, 429. https://doi.org/10.3390/v14020429
Kang H-J, Chu K-B, Yoon K-W, Eom G-D, Mao J, Kim M-J, Lee S-H, Moon E-K, Quan F-S. Multiple Neuraminidase Containing Influenza Virus-like Particle Vaccines Protect Mice from Avian and Human Influenza Virus Infection. Viruses. 2022; 14(2):429. https://doi.org/10.3390/v14020429
Chicago/Turabian StyleKang, Hae-Ji, Ki-Back Chu, Keon-Woong Yoon, Gi-Deok Eom, Jie Mao, Min-Ju Kim, Su-Hwa Lee, Eun-Kyung Moon, and Fu-Shi Quan. 2022. "Multiple Neuraminidase Containing Influenza Virus-like Particle Vaccines Protect Mice from Avian and Human Influenza Virus Infection" Viruses 14, no. 2: 429. https://doi.org/10.3390/v14020429
APA StyleKang, H.-J., Chu, K.-B., Yoon, K.-W., Eom, G.-D., Mao, J., Kim, M.-J., Lee, S.-H., Moon, E.-K., & Quan, F.-S. (2022). Multiple Neuraminidase Containing Influenza Virus-like Particle Vaccines Protect Mice from Avian and Human Influenza Virus Infection. Viruses, 14(2), 429. https://doi.org/10.3390/v14020429