
Flavonoids Target Human Herpesviruses That Infect the Nervous System: Mechanisms of Action and Therapeutic Insights


 ,
,  and
and 
Abstract
:1. Introduction
2. Herpesvirus Infections of the Nervous System and the Innate Immune Response
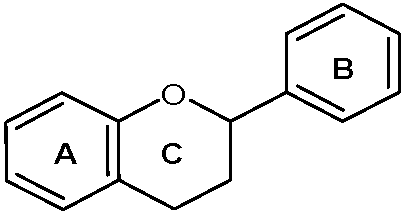
3. Flavonoids: Chemical Background, Antiviral, and Neuroprotective Activities
4. Flavonoids Target Human Herpesviruses of the Nervous System
4.1. Human Alpha-Herpesvirus Infections and Their Neurological Complications
Flavonoids Target Human Alpha-Herpesviruses
4.2. Human Beta-Herpesvirus Infections and Their Neurological Complications
Flavonoids Target Human Cytomegalovirus
4.3. Human Gamma-Herpesvirus Infections and Their Neurological Complications
Flavonoids Target Human Gamma-Herpesviruses
5. Strategies Involving Flavonoids for Enhancing Herpesvirus Treatment
6. Conclusions and Future Outlook
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stempel, M.; Chan, B.; Brinkmann, M.M. Coevolution Pays off: Herpesviruses Have the License to Escape the DNA Sensing Pathway. Med. Microbiol. Immunol. 2019, 208, 495–512. [Google Scholar] [CrossRef] [PubMed]
- Bhowmik, D.; Zhu, F. Evasion of Intracellular DNA Sensing by Human Herpesviruses. Front. Cell Infect. Microbiol. 2021, 11, 647992. [Google Scholar] [CrossRef] [PubMed]
- Adler, B.; Sattler, C.; Adler, H. Herpesviruses and Their Host Cells: A Successful Liaison. Trends Microbiol. 2017, 25, 229–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Šudomová, M.; Hassan, S.T.S. Nutraceutical Curcumin with Promising Protection against Herpesvirus Infections and Their Associated Inflammation: Mechanisms and Pathways. Microorganisms 2021, 9, 292. [Google Scholar] [CrossRef] [PubMed]
- Hassan, S.T.S.; Šudomová, M.; Masarčíková, R. Herpes simplex virus infection: An overview of the problem, pharmacologic therapy and dietary measures. Ceska Slov. Farm. 2017, 66, 95–102. [Google Scholar] [PubMed]
- Hassan, S.T.S. Shedding Light on the Effect of Natural Anti-Herpesvirus Alkaloids on SARS-CoV-2: A Treatment Option for COVID-19. Viruses 2020, 12, E476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sausen, D.G.; Reed, K.M.; Bhutta, M.S.; Gallo, E.S.; Borenstein, R. Evasion of the Host Immune Response by Betaherpesviruses. Int. J. Mol. Sci. 2021, 22, 7503. [Google Scholar] [CrossRef] [PubMed]
- Wen, K.W.; Wang, L.; Menke, J.R.; Damania, B. Cancers Associated with Human Gammaherpesviruses. FEBS J. 2021. [Google Scholar] [CrossRef] [PubMed]
- Pei, Y.; Robertson, E.S. The Crosstalk of Epigenetics and Metabolism in Herpesvirus Infection. Viruses 2020, 12, 1377. [Google Scholar] [CrossRef] [PubMed]
- Weidner-Glunde, M.; Kruminis-Kaszkiel, E.; Savanagouder, M. Herpesviral Latency-Common Themes. Pathogens 2020, 9, 125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Šudomová, M.; Berchová-Bímová, K.; Marzocco, S.; Liskova, A.; Kubatka, P.; Hassan, S.T.S. Berberine in Human Oncogenic Herpesvirus Infections and Their Linked Cancers. Viruses 2021, 13, 1014. [Google Scholar] [CrossRef] [PubMed]
- Hassan, S.T.S.; Berchová-Bímová, K.; Petráš, J.; Hassan, K.T.S. Cucurbitacin B Interacts Synergistically with Antibiotics against Staphylococcus Aureus Clinical Isolates and Exhibits Antiviral Activity against HSV-1. S. Afr. J. Bot. 2017, 108, 90–94. [Google Scholar] [CrossRef]
- Hassan, S.T.S.; Šudomová, M.; Berchová-Bímová, K.; Šmejkal, K.; Echeverría, J. Psoromic Acid, a Lichen-Derived Molecule, Inhibits the Replication of HSV-1 and HSV-2, and Inactivates HSV-1 DNA Polymerase: Shedding Light on Antiherpetic Properties. Molecules 2019, 24, 2912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poole, C.L.; James, S.H. Antiviral Therapies for Herpesviruses: Current Agents and New Directions. Clin. Ther. 2018, 40, 1282–1298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Treml, J.; Gazdová, M.; Šmejkal, K.; Šudomová, M.; Kubatka, P.; Hassan, S.T.S. Natural Products-Derived Chemicals: Breaking Barriers to Novel Anti-HSV Drug Development. Viruses 2020, 12, 154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brezáni, V.; Leláková, V.; Hassan, S.T.S.; Berchová-Bímová, K.; Nový, P.; Klouček, P.; Maršík, P.; Dall’Acqua, S.; Hošek, J.; Šmejkal, K. Anti-Infectivity against Herpes Simplex Virus and Selected Microbes and Anti-Inflammatory Activities of Compounds Isolated from Eucalyptus Globulus Labill. Viruses 2018, 10, 360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Čulenová, M.; Sychrová, A.; Hassan, S.T.S.; Berchová-Bímová, K.; Svobodová, P.; Helclová, A.; Michnová, H.; Hošek, J.; Vasilev, H.; Suchý, P.; et al. Multiple In Vitro Biological Effects of Phenolic Compounds from Morus Alba Root Bark. J. Ethnopharmacol. 2020, 248, 112296. [Google Scholar] [CrossRef] [PubMed]
- Hassan, S.T.S.; Švajdlenka, E. Biological Evaluation and Molecular Docking of Protocatechuic Acid from Hibiscus Sabdariffa, L. as a Potent Urease Inhibitor by an ESI-MS Based Method. Molecules 2017, 22, 1696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassan, S.T.S.; Švajdlenka, E.; Berchová-Bímová, K. Hibiscus Sabdariffa, L. and Its Bioactive Constituents Exhibit Antiviral Activity against HSV-2 and Anti-Enzymatic Properties against Urease by an ESI-MS Based Assay. Molecules 2017, 22, 722. [Google Scholar] [CrossRef] [PubMed]
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; International Natural Product Sciences Taskforce; Supuran, C.T. Natural Products in Drug Discovery: Advances and Opportunities. Nat. Rev. Drug. Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef] [PubMed]
- Liskova, A.; Samec, M.; Koklesova, L.; Brockmueller, A.; Zhai, K.; Abdellatif, B.; Siddiqui, M.; Biringer, K.; Kudela, E.; Pec, M.; et al. Flavonoids as an Effective Sensitizer for Anti-Cancer Therapy: Insights into Multi-Faceted Mechanisms and Applicability towards Individualized Patient Profiles. EPMA J. 2021, 12, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Bharucha, T.; Houlihan, C.F.; Breuer, J. Herpesvirus Infections of the Central Nervous System. Semin. Neurol. 2019, 39, 369–382. [Google Scholar] [CrossRef] [PubMed]
- Kawada, J.-I. Neurological Disorders Associated with Human Alphaherpesviruses. Adv. Exp. Med. Biol. 2018, 1045, 85–102. [Google Scholar] [CrossRef] [PubMed]
- Eliassen, E.; Hemond, C.C.; Santoro, J.D. HHV-6-Associated Neurological Disease in Children: Epidemiologic, Clinical, Diagnostic, and Treatment Considerations. Pediatr. Neurol. 2020, 105, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Jakhmola, S.; Upadhyay, A.; Jain, K.; Mishra, A.; Jha, H.C. Herpesviruses and the Hidden Links to Multiple Sclerosis Neuropathology. J. Neuroimmunol. 2021, 358, 577636. [Google Scholar] [CrossRef] [PubMed]
- Francis, S.S.; Wiemels, J.L.; Yang, W.; Shaw, G.M. Herpesvirus Infection in Infants with Gastroschisis. Epidemiology 2018, 29, 571–573. [Google Scholar] [CrossRef] [PubMed]
- Jarosinski, K.W. Interindividual Spread of Herpesviruses. Adv. Anat. Embryol. Cell Biol. 2017, 223, 195–224. [Google Scholar] [CrossRef] [PubMed]
- Azab, W.; Osterrieder, K. Initial Contact: The First Steps in Herpesvirus Entry. Adv. Anat. Embryol. Cell Biol. 2017, 223, 1–27. [Google Scholar] [CrossRef]
- Connolly, S.A.; Jardetzky, T.S.; Longnecker, R. The Structural Basis of Herpesvirus Entry. Nat. Rev. Microbiol. 2021, 19, 110–121. [Google Scholar] [CrossRef]
- Gilden, D.H.; Mahalingam, R.; Cohrs, R.J.; Tyler, K.L. Herpesvirus Infections of the Nervous System. Nat. Clin. Pr. Neurol. 2007, 3, 82–94. [Google Scholar] [CrossRef]
- Baldwin, K.J.; Cummings, C.L. Herpesvirus Infections of the Nervous System. Continuum: Lifelong Learn. Neurol. 2018, 24, 1349–1369. [Google Scholar] [CrossRef] [PubMed]
- Reese, T.A. Coinfections: Another Variable in the Herpesvirus Latency-Reactivation Dynamic. J. Virol. 2016, 90, 5534–5537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lomonte, P. Herpesvirus Latency: On the Importance of Positioning Oneself. Adv. Anat. Embryol. Cell Biol. 2017, 223, 95–117. [Google Scholar] [CrossRef] [PubMed]
- Sadeghipour, S.; Mathias, R.A. Herpesviruses Hijack Host Exosomes for Viral Pathogenesis. Semin. Cell Dev. Biol. 2017, 67, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Ostler, J.B.; Sawant, L.; Harrison, K.; Jones, C. Regulation of Neurotropic Herpesvirus Productive Infection and Latency-Reactivation Cycle by Glucocorticoid Receptor and Stress-Induced Transcription Factors. Vitam. Horm. 2021, 117, 101–132. [Google Scholar] [CrossRef] [PubMed]
- Soares, B.P.; Provenzale, J.M. Imaging of Herpesvirus Infections of the CNS. AJR Am. J. Roentgenol. 2016, 206, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Chijioke, O.; Azzi, T.; Nadal, D.; Münz, C. Innate Immune Responses against Epstein Barr Virus Infection. J. Leukoc. Biol. 2013, 94, 1185–1190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paludan, S.R.; Bowie, A.G.; Horan, K.A.; Fitzgerald, K.A. Recognition of Herpesviruses by the Innate Immune System. Nat. Rev. Immunol. 2011, 11, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Jondle, C.N.; Tarakanova, V.L. Innate Immunity and Alpha/Gammaherpesviruses: First Impressions Last a Lifetime. Curr. Opin. Virol. 2020, 44, 81–89. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, C.M.; Sen, G.C. Innate Immune Responses to Herpesvirus Infection. Cells 2021, 10, 2122. [Google Scholar] [CrossRef] [PubMed]
- Glaunsinger, B.A.; Ganem, D.E. Messenger RNA Turnover and Its Regulation in Herpesviral Infection. Adv. Virus Res. 2006, 66, 337–394. [Google Scholar] [CrossRef] [PubMed]
- Glaunsinger, B.A. Modulation of the Translational Landscape During Herpesvirus Infection. Annu. Rev. Virol. 2015, 2, 311–333. [Google Scholar] [CrossRef] [PubMed]
- Tucker, J.M.; Glaunsinger, B.A. Host Noncoding Retrotransposons Induced by DNA Viruses: A SINE of Infection? J. Virol. 2017, 91, e00982-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Covarrubias, S.; Richner, J.M.; Clyde, K.; Lee, Y.J.; Glaunsinger, B.A. Host Shutoff Is a Conserved Phenotype of Gammaherpesvirus Infection and Is Orchestrated Exclusively from the Cytoplasm. J. Virol. 2009, 83, 9554–9566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abernathy, E.; Clyde, K.; Yeasmin, R.; Krug, L.T.; Burlingame, A.; Coscoy, L.; Glaunsinger, B. Gammaherpesviral Gene Expression and Virion Composition Are Broadly Controlled by Accelerated MRNA Degradation. PLoS Pathog. 2014, 10, e1003882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asha, K.; Sharma-Walia, N. Targeting Host Cellular Factors as a Strategy of Therapeutic Intervention for Herpesvirus Infections. Front. Cell. Infect. Microbiol. 2021, 11, 603309. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Zhang, Y.; Hu, J.; Wu, W.; Chen, X.; Lu, Z.; Yang, R.; Huang, Y.; Fan, J. Specific T-Cell Receptor Gene Transfer Enhances Immune Response: A Potential Therapeutic Strategy for the Control of Human Cytomegalovirus Infection in Immunocompromised Patients. Cell Immunol. 2019, 336, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Martínez, G.; Mijares, M.R.; De Sanctis, J.B. Effects of Flavonoids and Its Derivatives on Immune Cell Responses. Recent Pat. Inflamm. Allergy Drug Discov. 2019, 13, 84–104. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Conlon, M.; Ren, W.; Chen, B.B.; Bączek, T. Natural Products as Targeted Modulators of the Immune System. J. Immunol. Res. 2018, 2018, 7862782. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.; Jiang, H.; Fang, J. Regulation of Immune Function by Polyphenols. J. Immunol. Res. 2018, 2018, 1264074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burkard, M.; Leischner, C.; Lauer, U.M.; Busch, C.; Venturelli, S.; Frank, J. Dietary Flavonoids and Modulation of Natural Killer Cells: Implications in Malignant and Viral Diseases. J. Nutr. Biochem. 2017, 46, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Rengasamy, K.R.R.; Khan, H.; Gowrishankar, S.; Lagoa, R.J.L.; Mahomoodally, F.M.; Khan, Z.; Suroowan, S.; Tewari, D.; Zengin, G.; Hassan, S.T.S.; et al. The Role of Flavonoids in Autoimmune Diseases: Therapeutic Updates. Pharmacol. Ther. 2019, 194, 107–131. [Google Scholar] [CrossRef] [PubMed]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An Overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Safe, S.; Jayaraman, A.; Chapkin, R.S.; Howard, M.; Mohankumar, K.; Shrestha, R. Flavonoids: Structure-Function and Mechanisms of Action and Opportunities for Drug Development. Toxicol. Res. 2021, 37, 147–162. [Google Scholar] [CrossRef] [PubMed]
- Khodzhaieva, R.S.; Gladkov, E.S.; Kyrychenko, A.; Roshal, A.D. Progress and Achievements in Glycosylation of Flavonoids. Front. Chem. 2021, 9, 637994. [Google Scholar] [CrossRef] [PubMed]
- Durazzo, A.; Lucarini, M.; Souto, E.B.; Cicala, C.; Caiazzo, E.; Izzo, A.A.; Novellino, E.; Santini, A. Polyphenols: A Concise Overview on the Chemistry, Occurrence, and Human Health. Phytother. Res. 2019, 33, 2221–2243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singla, R.K.; Dubey, A.K.; Garg, A.; Sharma, R.K.; Fiorino, M.; Ameen, S.M.; Haddad, M.A.; Al-Hiary, M. Natural Polyphenols: Chemical Classification, Definition of Classes, Subcategories, and Structures. J. AOAC Int. 2019, 102, 1397–1400. [Google Scholar] [CrossRef] [PubMed]
- Dias, M.C.; Pinto, D.C.G.A.; Silva, A.M.S. Plant Flavonoids: Chemical Characteristics and Biological Activity. Molecules 2021, 26, 5377. [Google Scholar] [CrossRef] [PubMed]
- Koklesova, L.; Liskova, A.; Samec, M.; Zhai, K.; Al-Ishaq, R.K.; Bugos, O.; Šudomová, M.; Biringer, K.; Pec, M.; Adamkov, M.; et al. Protective Effects of Flavonoids Against Mitochondriopathies and Associated Pathologies: Focus on the Predictive Approach and Personalized Prevention. Int. J. Mol. Sci. 2021, 22, 8649. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Pandey, A.K. Chemistry and Biological Activities of Flavonoids: An Overview. Sci. World J. 2013, 2013, 162750. [Google Scholar] [CrossRef] [Green Version]
- Zakaryan, H.; Arabyan, E.; Oo, A.; Zandi, K. Flavonoids: Promising Natural Compounds against Viral Infections. Arch. Virol. 2017, 162, 2539–2551. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.; Khan, H.; Aschner, M.; Hasan, M.M.; Hassan, S.T.S. Therapeutic Potential of Naringin in Neurological Disorders. Food Chem. Toxicol. 2019, 132, 110646. [Google Scholar] [CrossRef] [PubMed]
- Badshah, S.L.; Faisal, S.; Muhammad, A.; Poulson, B.G.; Emwas, A.H.; Jaremko, M. Antiviral Activities of Flavonoids. Biomed. Pharm. 2021, 140, 111596. [Google Scholar] [CrossRef] [PubMed]
- Sharma, V.; Sehrawat, N.; Sharma, A.; Yadav, M.; Verma, P.; Sharma, A.K. Multifaceted Antiviral Therapeutic Potential of Dietary Flavonoids: Emerging Trends and Future Perspectives. Biotechnol. Appl. Biochem. 2021. [Google Scholar] [CrossRef] [PubMed]
- Hassan, S.T.S.; Masarčíková, R.; Berchová, K. Bioactive Natural Products with Anti-Herpes Simplex Virus Properties. J. Pharm. Pharm. 2015, 67, 1325–1336. [Google Scholar] [CrossRef] [PubMed]
- Di Lorenzo, C.; Colombo, F.; Biella, S.; Stockley, C.; Restani, P. Polyphenols and Human Health: The Role of Bioavailability. Nutrients 2021, 13, 273. [Google Scholar] [CrossRef] [PubMed]
- Teng, H.; Chen, L. Polyphenols and Bioavailability: An Update. Crit. Rev. Food Sci. Nutr. 2019, 59, 2040–2051. [Google Scholar] [CrossRef] [PubMed]
- Hassan, S.T.S.; Žemlička, M. Plant-Derived Urease Inhibitors as Alternative Chemotherapeutic Agents. Arch. Pharm. 2016, 349, 507–522. [Google Scholar] [CrossRef] [PubMed]
- Herpes Simplex Virus. Available online: https://www.who.int/news-room/fact-sheets/detail/herpes-simplex-virus (accessed on 11 December 2021).
- Huang, Y.; Song, Y.; Li, J.; Lv, C.; Chen, Z.-S.; Liu, Z. Receptors and Ligands for Herpes Simplex Viruses: Novel Insights for Drug Targeting. Drug Discov. Today 2021, 27, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Viejo-Borbolla, A. Pathogenesis and Virulence of Herpes Simplex Virus. Virulence 2021, 12, 2670–2702. [Google Scholar] [CrossRef] [PubMed]
- Colombo, D.; Albore, M.; Nonno, F.D.; Bolino, G.; D’Ambrosio, M.; Salvi, A.; Cecannecchia, C.; Falasca, L. Fatal Fulminant HSV-2 Myocarditis: A Complicated Presentation. Int. J. Infect. Dis. 2021, 114, 124–127. [Google Scholar] [CrossRef] [PubMed]
- Hendrickx, D.M.; Sousa, J.D.; Libin, P.J.K.; Delva, W.; Liesenborgs, J.; Hens, N.; Müller, V.; Vandamme, A.-M. Comparison of Two Simulators for Individual Based Models in HIV Epidemiology in a Population with HSV 2 in Yaoundé (Cameroon). Sci. Rep. 2021, 11, 14696. [Google Scholar] [CrossRef] [PubMed]
- Edwards, R.J.; Dolly, N.; Musa, D.; Edwards, J.; Boyce, G. Clinical Presentation of Herpes Simplex Virus Infection Mimicking Neoplasia on the Face of Persons Living with HIV. Int. J. STD. AIDS 2022, 33, 212–214. [Google Scholar] [CrossRef] [PubMed]
- Bergström, P.; Trybala, E.; Eriksson, C.E.; Johansson, M.; Satir, T.M.; Widéhn, S.; Fruhwürth, S.; Michno, W.; Nazir, F.H.; Hanrieder, J.; et al. Herpes Simplex Virus 1 and 2 Infections during Differentiation of Human Cortical Neurons. Viruses 2021, 13, 2072. [Google Scholar] [CrossRef] [PubMed]
- DuRaine, G.; Johnson, D.C. Anterograde Transport of α-Herpesviruses in Neuronal Axons. Virology 2021, 559, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Hersh, N.; Steiner, I.; Siegal, T.; Benninger, F. Herpes Simplex Encephalitis in Patients Receiving Chemotherapy and Whole-Brain Radiation Therapy. J. Neurovirol. 2021, 27, 774–781. [Google Scholar] [CrossRef] [PubMed]
- Ordoñez, G.; Vales, O.; Pineda, B.; Rodríguez, K.; Pane, C.; Sotelo, J. The Presence of Herpes Simplex-1 and Varicella Zoster Viruses Is Not Related with Clinical Outcome of Bell’s Palsy. Virology 2020, 549, 85–88. [Google Scholar] [CrossRef] [PubMed]
- Akkaya, O. Prevalence of Herpes Simplex Virus Infections in the Central Nervous System. Clin. Lab. 2021, 67. [Google Scholar] [CrossRef] [PubMed]
- Rozenberg, F. Herpes simplex virus and central nervous system infections: Encephalitis, meningitis, myelitis. Virologie 2020, 24, 283–294. [Google Scholar] [CrossRef] [PubMed]
- Gershon, A.A.; Breuer, J.; Cohen, J.I.; Cohrs, R.J.; Gershon, M.D.; Gilden, D.; Grose, C.; Hambleton, S.; Kennedy, P.G.E.; Oxman, M.N.; et al. Varicella Zoster Virus Infection. Nat. Rev. Dis. Primers. 2015, 1, 15016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braspenning, S.E.; Sadaoka, T.; Breuer, J.; Verjans, G.M.G.M.; Ouwendijk, W.J.D.; Depledge, D.P. Decoding the Architecture of the Varicella-Zoster Virus Transcriptome. mBio 2020, 11, e01568-20. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, P.G.E.; Cohrs, R.J. Varicella-Zoster Virus Human Ganglionic Latency: A Current Summary. J. Neurovirol. 2010, 16, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, P.G.E.; Mogensen, T.H.; Cohrs, R.J. Recent Issues in Varicella-Zoster Virus Latency. Viruses 2021, 13, 2018. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, P.G.E.; Mogensen, T.H. Varicella-Zoster Virus Infection of Neurons Derived from Neural Stem Cells. Viruses 2021, 13, 485. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, P.G.E.; Gershon, A.A. Clinical Features of Varicella-Zoster Virus Infection. Viruses 2018, 10, 609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kennedy, P.G.E. Issues in the Treatment of Neurological Conditions Caused by Reactivation of Varicella Zoster Virus (VZV). Neurotherapeutics 2016, 13, 509–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corral, C.; Quereda, C.; Muriel, A.; Martínez-Ulloa, P.-L.; González-Gómez, F.-J.; Corral, Í. Clinical Spectrum and Prognosis of Neurological Complications of Reactivated Varicella-Zoster Infection: The Role of Immunosuppression. J. Neurovirol. 2020, 26, 696–703. [Google Scholar] [CrossRef] [PubMed]
- Abu-Rumeileh, S.; Mayer, B.; Still, V.; Tumani, H.; Otto, M.; Senel, M. Varicella Zoster Virus-Induced Neurological Disease after COVID-19 Vaccination: A Retrospective Monocentric Study. J. Neurol. 2021, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Xu, C.; Hao, C.; Zhang, Y.; Wang, Z.; Wang, S.; Wang, W. Inhibition of Herpes Simplex Virus by Myricetin through Targeting Viral GD Protein and Cellular EGFR/PI3K/Akt Pathway. Antivir. Res. 2020, 177, 104714. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.-Y.; Gao, S.-Q.; Gong, Y.-S.; Lin, T.; Tong, S.; Xiong, W.; Shi, C.-Y.; Wang, W.-Q.; Fang, J.-G. Anti-HSV-1 Effect of Dihydromyricetin from Ampelopsis Grossedentata via the TLR9-Dependent Anti-Inflammatory Pathway. J. Glob. Antimicrob. Resist. 2020, 23, 370–376. [Google Scholar] [CrossRef]
- Ortega, J.T.; Serrano, M.L.; Suárez, A.I.; Baptista, J.; Pujol, F.H.; Cavallaro, L.V.; Campos, H.R.; Rangel, H.R. Antiviral Activity of Flavonoids Present in Aerial Parts of Marcetia Taxifolia against Hepatitis B Virus, Poliovirus, and Herpes Simplex Virus in Vitro. EXCLI J. 2019, 18, 1037–1048. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.I.; Kwon, E.-B.; Oh, Y.-C.; Go, Y.; Choi, J.-G. Mori Ramulus and Its Major Component Morusin Inhibit Herpes Simplex Virus Type 1 Replication and the Virus-Induced Reactive Oxygen Species. Am. J. Chin. Med. 2021, 49, 163–179. [Google Scholar] [CrossRef] [PubMed]
- Chu, Y.; Lv, X.; Zhang, L.; Fu, X.; Song, S.; Su, A.; Chen, D.; Xu, L.; Wang, Y.; Wu, Z.; et al. Wogonin Inhibits in Vitro Herpes Simplex Virus Type 1 and 2 Infection by Modulating Cellular NF-ΚB and MAPK Pathways. BMC Microbiol. 2020, 20, 227. [Google Scholar] [CrossRef] [PubMed]
- Rittà, M.; Marengo, A.; Civra, A.; Lembo, D.; Cagliero, C.; Kant, K.; Lal, U.R.; Rubiolo, P.; Ghosh, M.; Donalisio, M. Antiviral Activity of a Arisaema Tortuosum Leaf Extract and Some of Its Constituents against Herpes Simplex Virus Type 2. Planta. Med. 2020, 86, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Fahmy, N.M.; Al-Sayed, E.; Moghannem, S.; Azam, F.; El-Shazly, M.; Singab, A.N. Breaking Down the Barriers to a Natural Antiviral Agent: Antiviral Activity and Molecular Docking of Erythrina Speciosa Extract, Fractions, and the Major Compound. Chem. Biodivers. 2020, 17, e1900511. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Song, X.; Su, G.; Wang, Y.; Wang, Z.; Jia, J.; Qing, S.; Huang, L.; Wang, Y.; Zheng, K.; et al. Amentoflavone Inhibits HSV-1 and ACV-Resistant Strain Infection by Suppressing Viral Early Infection. Viruses 2019, 11, 466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.; Lee, H.H.; Shin, Y.S.; Kang, H.; Cho, H. The Anti-HSV-1 Effect of Quercetin Is Dependent on the Suppression of TLR-3 in Raw 264.7 Cells. Arch. Pharm. Res. 2017, 40, 623–630. [Google Scholar] [CrossRef] [PubMed]
- Pradhan, P.; Nguyen, M.L. Herpes Simplex Virus Virucidal Activity of MST-312 and Epigallocatechin Gallate. Virus Res. 2018, 249, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.-Y.; Yu, Z.-Y.; Chen, Y.-C.; Hung, S.-L. Effects of Epigallocatechin-3-Gallate and Acyclovir on Herpes Simplex Virus Type 1 Infection in Oral Epithelial Cells. J. Med. Assoc. 2021, 120, 2136–2143. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-Q.; Cai, L.; Zhang, N.; Zhang, J.; Wang, H.-H.; Zhu, W. Protective Effect of Total Flavonoids from Ixeris Sonchifolia on Herpes Simplex Virus Keratitis in Mice. BMC Complement. Med. 2020, 20, 113. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.H.; Kim, J.-E.; Song, Y.-J. Antiviral Activities of Quercetin and Isoquercitrin Against Human Herpesviruses. Molecules 2020, 25, 2379. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Liu, L.; Wu, H.; Chen, S.; Zhu, Q.; Gao, H.; Yu, X.; Wang, Y.; Su, W.; Yao, X.; et al. Anti-Herpes Simplex Virus Type 1 Activity of Houttuynoid A, a Flavonoid from Houttuynia Cordata Thunb. Antivir. Res. 2017, 144, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Li, J.-J.; Chen, G.-D.; Fan, H.-X.; Hu, D.; Zhou, Z.-Q.; Lan, K.-H.; Zhang, H.-P.; Maeda, H.; Yao, X.-S.; Gao, H. Houttuynoid M, an Anti-HSV Active Houttuynoid from Houttuynia Cordata Featuring a Bis-Houttuynin Chain Tethered to a Flavonoid Core. J. Nat. Prod. 2017, 80, 3010–3013. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, P.; Reeves, M. Pathogenesis of Human Cytomegalovirus in the Immunocompromised Host. Nat. Rev. Microbiol. 2021, 19, 759–773. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, C.M. Cytomegalovirus (CMV) Infection and Latency. Pathogens 2021, 10, 342. [Google Scholar] [CrossRef] [PubMed]
- Prosser, J.D.; Holmes, T.W.; Seyyedi, M.; Choo, D.I. Congenital Cytomegalovirus (CMV) for the Pediatric Otolaryngologist. Int J. Pediatr. Otorhinolaryngol. 2021, 148, 110809. [Google Scholar] [CrossRef] [PubMed]
- Plosa, E.J.; Esbenshade, J.C.; Fuller, M.P.; Weitkamp, J.-H. Cytomegalovirus Infection. Pediatr. Rev. 2012, 33, 156–163. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, P.; Baraniak, I.; Reeves, M. The Pathogenesis of Human Cytomegalovirus. J. Pathol. 2015, 235, 288–297. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-Y.; Fang, F. Congenital Human Cytomegalovirus Infection and Neurologic Diseases in Newborns. Chin. Med. J. 2019, 132, 2109–2118. [Google Scholar] [CrossRef] [PubMed]
- Tselis, A.C. Cytomegalovirus Infections of the Adult Human Nervous System. Handb. Clin. Neurol. 2014, 123, 307–318. [Google Scholar] [CrossRef] [PubMed]
- Ting, P.; Camaj, A.; Bienstock, S.; Choy, A.; Mitter, S.S.; Barghash, M.; Mancini, D. Guillain-Barré Syndrome After Primary Cytomegalovirus Infection in a Patient With a Heart Transplant. JACC Case Rep. 2021, 3, 455–458. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Jiang, L. Cytomegalovirus Encephalitis in Immunocompetent Infants: A 15-Year Retrospective Study at a Single Center. Int. J. Infect. Dis. 2019, 82, 106–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nukui, M.; O’Connor, C.M.; Murphy, E.A. The Natural Flavonoid Compound Deguelin Inhibits HCMV Lytic Replication within Fibroblasts. Viruses 2018, 10, 614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akai, Y.; Sadanari, H.; Takemoto, M.; Uchide, N.; Daikoku, T.; Mukaida, N.; Murayama, T. Inhibition of Human Cytomegalovirus Replication by Tricin Is Associated with Depressed CCL2 Expression. Antivir. Res. 2017, 148, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Itoh, A.; Sadanari, H.; Takemoto, M.; Matsubara, K.; Daikoku, T.; Murayama, T. Tricin Inhibits the CCL5 Induction Required for Efficient Growth of Human Cytomegalovirus. Microbiol. Immunol. 2018, 62, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Kapasi, A.J.; Clark, C.L.; Tran, K.; Spector, D.H. Recruitment of Cdk9 to the Immediate-Early Viral Transcriptosomes during Human Cytomegalovirus Infection Requires Efficient Binding to Cyclin T1, a Threshold Level of IE2 86, and Active Transcription. J. Virol. 2009, 83, 5904–5917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadanari, H.; Fujimoto, K.J.; Sugihara, Y.; Ishida, T.; Takemoto, M.; Daikoku, T.; Murayama, T. The Anti-Human Cytomegalovirus Drug Tricin Inhibits Cyclin-Dependent Kinase 9. FEBS Open Bio. 2018, 8, 646–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujimoto, K.J.; Nema, D.; Ninomiya, M.; Koketsu, M.; Sadanari, H.; Takemoto, M.; Daikoku, T.; Murayama, T. An in Silico-Designed Flavone Derivative, 6-Fluoro-4’-Hydroxy-3’,5’-Dimetoxyflavone, Has a Greater Anti-Human Cytomegalovirus Effect than Ganciclovir in Infected Cells. Antivir. Res. 2018, 154, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Lange, P.T.; White, M.C.; Damania, B. Activation and Evasion of Innate Immunity by Gammaherpesviruses. J. Mol. Biol. 2021, 167214. [Google Scholar] [CrossRef] [PubMed]
- Möhl, B.S.; Chen, J.; Longnecker, R. Gammaherpesvirus Entry and Fusion: A Tale How Two Human Pathogenic Viruses Enter Their Host Cells. Adv. Virus. Res. 2019, 104, 313–343. [Google Scholar] [CrossRef] [Green Version]
- Epstein, M.A.; Achong, B.G.; Barr, Y.M. Virus particles in cultured lymphoblasts from burkitt’s lymphoma. Lancet 1964, 1, 702–703. [Google Scholar] [CrossRef]
- Epstein, M.A.; Henle, G.; Achong, B.G.; Barr, Y.M. Morphological and biological studies on a virus in cultured lymphoblasts from burkitt’s lymphoma. J. Exp. Med. 1965, 121, 761–770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciccarese, G.; Trave, I.; Herzum, A.; Parodi, A.; Drago, F. Dermatological Manifestations of Epstein-Barr Virus Systemic Infection: A Case Report and Literature Review. Int. J. Derm. 2020, 59, 1202–1209. [Google Scholar] [CrossRef] [PubMed]
- Nowalk, A.; Green, M. Epstein-Barr Virus. Microbiol. Spectr. 2016, 4, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujimoto, H.; Asaoka, K.; Imaizumi, T.; Ayabe, M.; Shoji, H.; Kaji, M. Epstein-Barr Virus Infections of the Central Nervous System. Intern. Med. 2003, 42, 33–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ridha, M.; Jones, D.G.; Lerner, D.P.; Vytopil, M.; Voetsch, B.; Burns, J.D.; Ramineni, A.; Raibagkar, P. The Spectrum of Epstein-Barr Virus Infections of the Central Nervous System after Organ Transplantation. Virol. J. 2021, 18, 162. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S.; Angelov, L.; Ahluwalia, M.S.; Yeaney, G.A. Epstein-Barr Virus-Associated Primary Central Nervous System Lymphoma in a Patient with Diffuse Cutaneous Systemic Sclerosis on Long-Term Mycophenolate Mofetil. Jt. Bone Spine 2020, 87, 163–166. [Google Scholar] [CrossRef] [PubMed]
- Houen, G.; Trier, N.H.; Frederiksen, J.L. Epstein-Barr Virus and Multiple Sclerosis. Front. Immunol. 2020, 11, 587078. [Google Scholar] [CrossRef] [PubMed]
- Soldan, S.S.; Lieberman, P.M. Epstein-Barr Virus Infection in the Development of Neurological Disorders. Drug Discov. Today Dis. Models 2020, 32, 35–52. [Google Scholar] [CrossRef] [PubMed]
- Weinberg, A.; Li, S.; Palmer, M.; Tyler, K.L. Quantitative CSF PCR in Epstein-Barr Virus Infections of the Central Nervous System. Ann. Neurol. 2002, 52, 543–548. [Google Scholar] [CrossRef]
- Cheng, H.; Chen, D.; Peng, X.; Wu, P.; Jiang, L.; Hu, Y. Clinical Characteristics of Epstein–Barr Virus Infection in the Pediatric Nervous System. BMC Infect. Dis. 2020, 20, 886. [Google Scholar] [CrossRef] [PubMed]
- Goncalves, P.H.; Ziegelbauer, J.; Uldrick, T.S.; Yarchoan, R. Kaposi Sarcoma Herpesvirus-Associated Cancers and Related Diseases. Curr. Opin. HIV AIDS 2017, 12, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Bai, L.; Dong, J.; Sun, R.; Lan, K. Kaposi’s Sarcoma-Associated Herpesvirus: Epidemiology and Molecular Biology. Adv. Exp. Med. Biol. 2017, 1018, 91–127. [Google Scholar] [CrossRef] [PubMed]
- Tso, F.Y.; Sawyer, A.; Kwon, E.H.; Mudenda, V.; Langford, D.; Zhou, Y.; West, J.; Wood, C. Kaposi’s Sarcoma-Associated Herpesvirus Infection of Neurons in HIV Positive Patients. J. Infect. Dis. 2016, 215, jiw545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jha, H.C.; Mehta, D.; Lu, J.; El-Naccache, D.; Shukla, S.K.; Kovacsics, C.; Kolson, D.; Robertson, E.S. Gammaherpesvirus Infection of Human Neuronal Cells. mBio 2015, 6, e01844-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldini, F.; Baiocchini, A.; Schininà, V.; Agrati, C.; Giancola, M.L.; Alba, L.; Grisetti, S.; Del Nonno, F.; Capobianchi, M.R.; Antinori, A. Brain Localization of Kaposi’s Sarcoma in a Patient Treated by Combination Antiretroviral Therapy. BMC Infect. Dis. 2013, 13, 600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
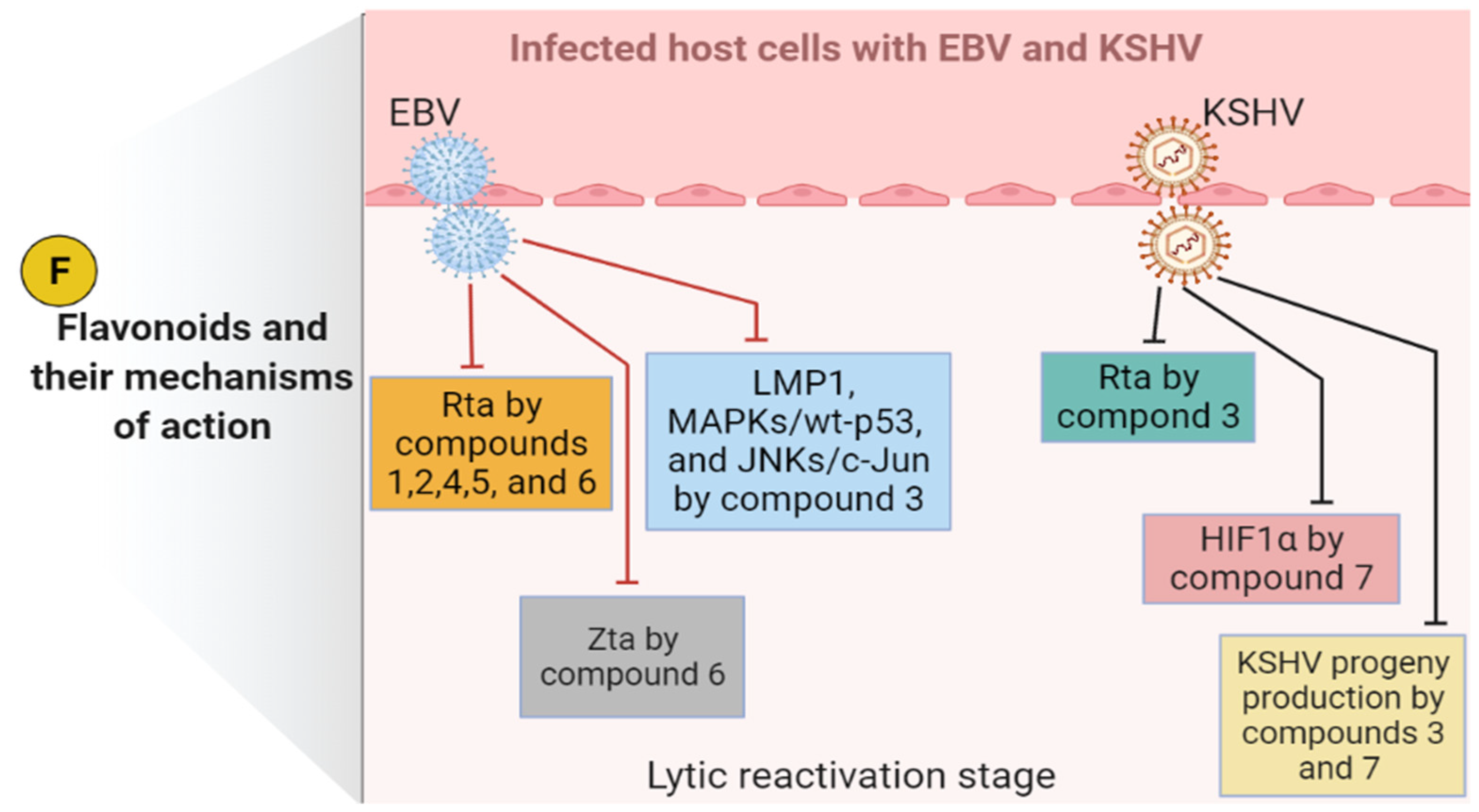
- Tsai, Y.-C.; Hohmann, J.; El-Shazly, M.; Chang, L.-K.; Dankó, B.; Kúsz, N.; Hsieh, C.-T.; Hunyadi, A.; Chang, F.-R. Bioactive Constituents of Lindernia Crustacea and Its Anti-EBV Effect via Rta Expression Inhibition in the Viral Lytic Cycle. J. Ethnopharmacol. 2020, 250, 112493. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.-H.; Chen, C.-S.; Wang, W.-H.; Hsu, S.-W.; Tsai, H.-H.; Liu, S.-T.; Chang, L.-K. TRIM5α Promotes Ubiquitination of Rta from Epstein–Barr Virus to Attenuate Lytic Progression. Front. Microbiol. 2017, 7, 1407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Li, Y.; Hu, J.; Liu, S.; Luo, X.; Tang, M.; Bode, A.M.; Dong, Z.; Liu, X.; Liao, W.; et al. ()-Epigallocatechin-3-Gallate Inhibits EBV Lytic Replication via Targeting LMP1-Mediated MAPK Signal Axes. Oncol. Res. 2021, 28, 763–778. [Google Scholar] [CrossRef] [PubMed]
- Vágvölgyi, M.; Girst, G.; Kúsz, N.; Ötvös, S.B.; Fülöp, F.; Hohmann, J.; Servais, J.-Y.; Seguin-Devaux, C.; Chang, F.-R.; Chen, M.S.; et al. Less Cytotoxic Protoflavones as Antiviral Agents: Protoapigenone 1’-O-Isopropyl Ether Shows Improved Selectivity Against the Epstein-Barr Virus Lytic Cycle. Int. J. Mol. Sci. 2019, 20, 6269. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.-C.; Fang, C.-Y.; Cheng, Y.-J.; Hsu, H.-Y.; Chou, S.-P.; Huang, S.-Y.; Tsai, C.-H.; Chen, J.-Y. Inhibition of Epstein-Barr Virus Reactivation by the Flavonoid Apigenin. J. Biomed. Sci. 2017, 24, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, C.-Y.; Chen, C.-Y.; Chiou, Y.-H.; Shyu, H.-W.; Lin, K.-H.; Chou, M.-C.; Huang, M.-H.; Wang, Y.-F. Epigallocatechin-3-Gallate Suppresses Human Herpesvirus 8 Replication and Induces ROS Leading to Apoptosis and Autophagy in Primary Effusion Lymphoma Cells. Int. J. Mol. Sci. 2017, 19, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, W.-Y.; Zhao, G.-H.; Wu, Y. Hesperetin Inhibits KSHV Reactivation and Is Reversed by HIF1α Overexpression. J. Gen. Virol. 2021, 102, 001686. [Google Scholar] [CrossRef] [PubMed]
- Polansky, H.; Javaherian, A.; Itzkovitz, E. Clinical Trial of Herbal Treatment Gene-Eden-VIR/Novirin in Oral Herpes. J. Evid. Based Integr. Med. 2018, 23, 2515690X18806269. [Google Scholar] [CrossRef] [PubMed]
- Caldas Dos Santos, T.; Rescignano, N.; Boff, L.; Reginatto, F.H.; Simões, C.M.O.; de Campos, A.M.; Mijangos, C. In Vitro Antiherpes Effect of C-Glycosyl Flavonoid Enriched Fraction of Cecropia Glaziovii Encapsulated in PLGA Nanoparticles. Mater. Sci. Eng. C Mater. Biol. Appl. 2017, 75, 1214–1220. [Google Scholar] [CrossRef] [PubMed]
- Brandariz-Nuñez, A.; Liu, T.; Du, T.; Evilevitch, A. Pressure-Driven Release of Viral Genome into a Host Nucleus Is a Mechanism Leading to Herpes Infection. eLife 2019, 8, e47212. [Google Scholar] [CrossRef] [PubMed]
- Brandariz-Nuñez, A.; Robinson, S.J.; Evilevitch, A. Pressurized DNA State inside Herpes Capsids-A Novel Antiviral Target. PLoS Pathog. 2020, 16, e1008604. [Google Scholar] [CrossRef] [PubMed]
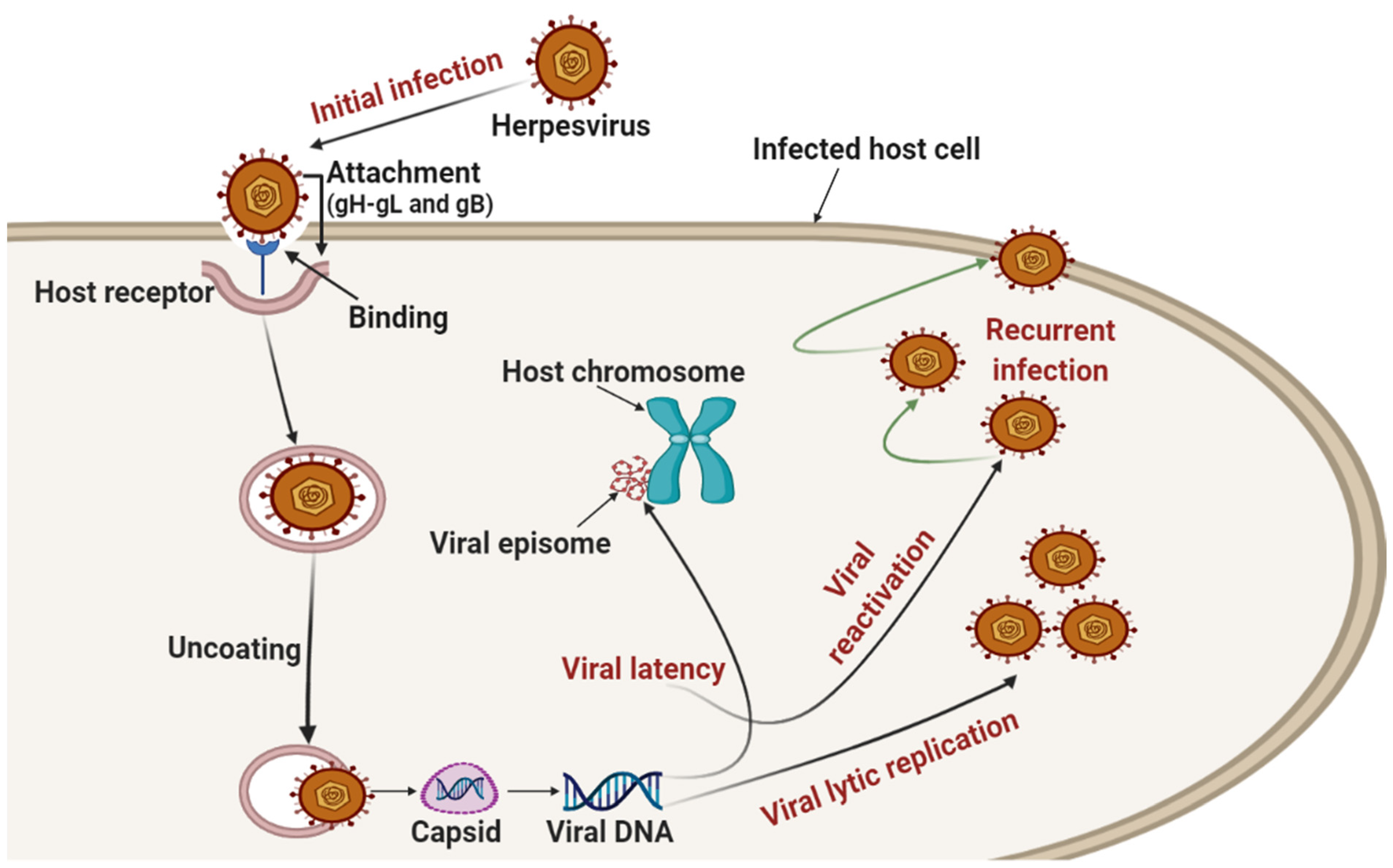
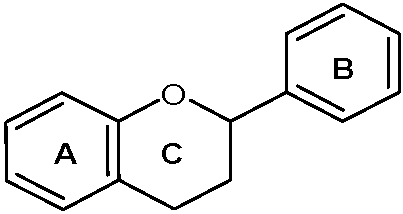
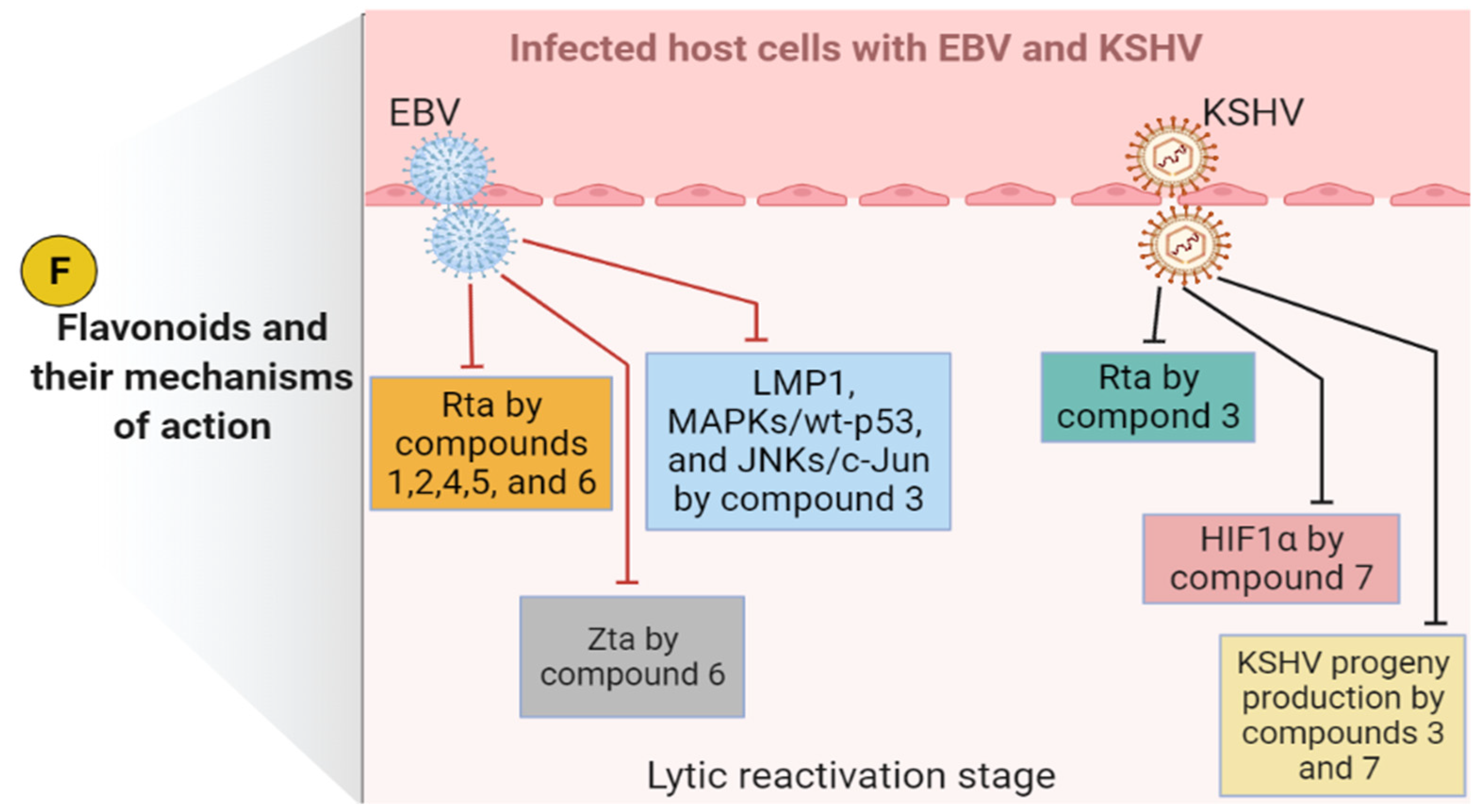

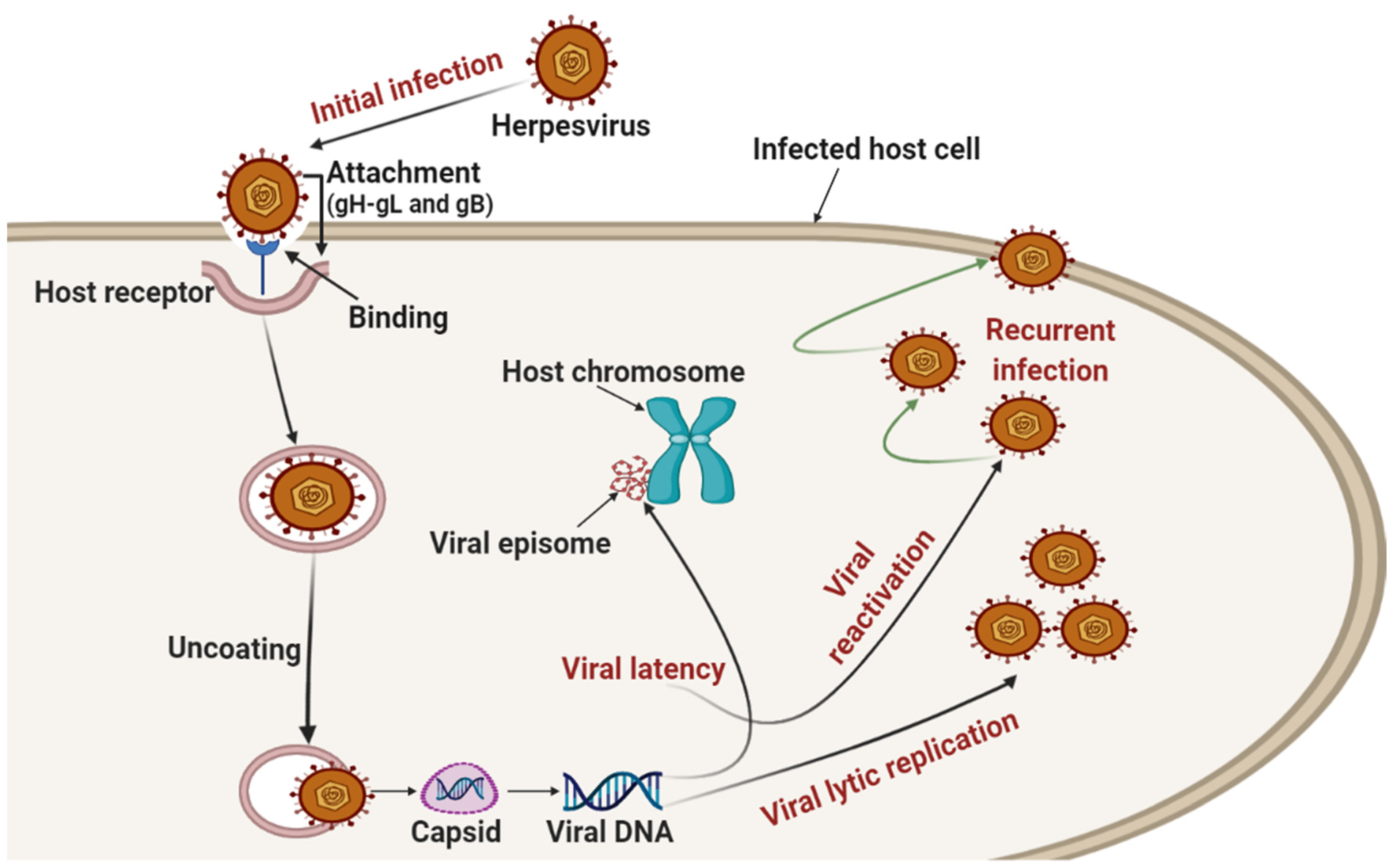{kind=link}
{kind=link}
{kind=link}
| Type of Study, Method, Virus, and Cells/Animal Model | Results (Compound, Concentration, or Dose) | Mechanisms | Reference |
|---|---|---|---|
| In vitro, in vivo, and in silico. Plaque-reduction assay, CPE-inhibition assay, time-of-addition assay, various biochemical methods, and molecular-docking studies. HSV-1 and HSV-2. Vero, HeLa, and Hep-2 cells. Three-week-old female BALB/c mice. | At concentrations ranging from 2.5 to 40 µM, myricetin in vitro effectively blocked HSV-1 and HSV-2 infections by interfering with virus adsorption and membrane fusion. In vivo, treatment with myricetin at 2.5 and 5 mg/kg inhibited the infection with HSV-1 in infected mice. In silico, myricetin was found to successfully bind to HSV-2 gD protein. | Interaction with HSV-2 gD protein. Downregulation of cellular EGFR/PI3K/Akt signaling pathway. | [90] |
| In vitro. Plaque-reduction assay, virus yield reduction assay combined with several biochemical investigations. HSV-1. Vero cells. | The potent anti-HSV-1 activity of dihydromyricetin was unveiled with an EC50 value of 12.56 µM. | HSV-1 plaque formation and progeny virus productions were inhibited by a mechanism via the diminishment of the expression of HSV-1 IE genes (ICP4 and ICP22), early genes (ICP8 and UL42), and late genes (gB, VP1/2) at concentrations of 16 and 32 µM. Inhibition of mRNA levels of TLR9. Suppression of NF-κB and TNFα. | [91] |
| In vitro. Plaque-reduction assay. Vero cells. | Treatment of HSV-1-infected Vero cells with PMF and PMF-OH notably suppressed the replication of HSV-1 with EC50 values of 6.8 and 5.9 µM, respectively. | No mechanism of action was revealed. | [92] |
| In vitro. Plaque-reduction assay and multi-biochemical studies. HSV-1. Vero cells. | Morusin significantly repressed the replication of HSV-1 in infected Vero cells at 20 µM. | Inhibition of HSV-1 gD expression. Suppression of HSV-1-induced ROS. | [93] |
| In vitro and in silico. Plaque-reduction assay, cytopathic end-point assay, and dye-uptake method. HSV-1 (KOS strain) and HSV-2 (clinical isolates). Vero cells. | Kuwanon C, Kuwanon T, and Kuwanon U potently inhibited HSV-1 replication with IC50 values of 0.91, 0.64, and 1.93 µg/mL, respectively, while Kuwanon E suppressed the replication of HSV-2 with an EC50 value of 1.61 µg/mL. | Targeting HSV-1 DNA polymerase and HSV-2 protease by molecular docking studies. | [17] |
| In vitro. Cytopathic effect assay associated with various biochemical analyses. Vero and HEC-1-A cells. | At various concentrations in µM, wogonin prevented the infection of HSV-1 and HSV-2 infections by inhibiting their replication. It inhibited HSV-2-induced CPE and decreased viral mRNA transcription, viral protein synthesis, and infectious virion particle. | The mechanism of action is mediated by modulating cellular NF-κB and JNK/p38 MAPK pathways. | [94] |
| In vitro. Plaque-reduction assay, time-of-addition assay, and post-entry assay. HSV-2 and acyclovir-resistant HSV-2 strain. Vero cells. | The replication of HSV-2 was suppressed by apigenin and luteolin with EC50 values of 0.05 and 0.41 µg/mL, respectively, for the HSV-2 standard strain and acyclovir-resistant HSV-2 strain. EC50 values were found to be 2.33 and 1.55 µg/mL, respectively. | The mechanism was ascertained by decreasing viral progeny production. Apigenin was recognized to prevent cell-to-cell virus spread. | [95] |
| In vitro and in silico. CPE and MTT assays. HSV-1 (clinical strain). Vero cells. | Vitexin demonstrated anti-HSV-1 activity with an EC50 value of 18 µg/mL. | Targeting HSV-1 DNA polymerase (predicted by a molecular docking analysis). | [96] |
| In vitro. Plaque-reduction assay, CPE assay, and other biochemical investigations. HSV-1 (F strain; standard strain) and ACV-resistant strains (HSV-1/106, HSV-1/153, and HSV-1/Blue). Vero cells. | The considerable antiviral activities of amentoflavone were observed against HSV-1 (F strain) and ACV-resistant strains (HSV-1/106, HSV-1/153, and HSV-1/Blue) with EC50 values of 22.13, 11.11, 28.22, and 25.71 µM, respectively. | Suppression of viral gene production (UL54, UL52, and UL27). Inhibition of IE protein ICP0 expression. Inhibition of nuclear import of HSV-1. | [97] |
| In vitro. Plaque-reduction assays, western blotting, real-time PCR, and ELISA assays. HSV-1. Raw 264.7 cells and Vero cells. | The anti-infectivity action of quercetin against infected Raw 264.7 cells with HSV-1 was identified at concentrations of 10, 20, and 30 µg/mL. | Inhibition of HSV-1 gene expressions (ICP0, UL13, and UL52). Suppression of gD expression. Inhibition of TLR-3 expression. Inhibition of NF-κB and IRF3 expressions. | [98] |
| In vitro. Plaque-reduction assay. Wild-type HSV-1 strain (KOS1.1) Vero cells. | The virucidal activity of epigallocatechin-3-gallate against HSV-1 was revealed at concentrations as low as 1–2 µM at temperatures between 25–37 °C. | Interfering with various steps in the HSV-1 life cycle. | [99] |
| In vitro. Plaque assay, MTT assay, Western blotting analysis, confocal laser scanning microscopy, and real-time PCR analyses. HSV-1 (KOS strain). Oral epithelial cells and Vero cells. | Treatment of oral epithelial cells with epigallocatechin-3-gallate (25 µg/mL) significantly impeded HSV-1-induced cell death. | Reducing the expression of viral IE and ICP0 proteins. Inhibition of viral particles and viral DNA during the viral entry phase. | [100] |
| In vivo. CPE assay coupled with biochemical and histopathological analyses. HSV-1 (SM44 standard strain). Seventy-five adult male BALB/c mice. | Treatment with ISH total flavonoids at 50, 100, and 200 mg/kg (applied orally twice a day for two weeks) significantly boosted the corneal lesion degree and enhanced mice survival rate. | ISH total flavonoids improved the levels of IL-2 and INF-γ and lowered the levels of IL-4 in the serum of mice. | [101] |
| In vitro. Plaque-reduction assay with several biochemical assessments. Recombinant laboratory pOka strain of VZV (VZV–pOka). HFF cells. | Quercetin and isoquercitrin exhibited strong anti-VZV properties with IC50 values of 3.8 and 14.4 µg/mL, respectively. | Inhibition of VZV lytic-genes expressions. Reduction of VZV ORF62 (IE), ORF28 (E), and gB (L) transcripts levels. | [102] |
| In vitro and in vivo. β-galactosidase-activity assay, luciferase-activity assay, plaque-reduction assay, progeny-HSV-1-yield assay, time-of-addition assay, fusion-inhibition assay, and other biochemical methods. HSV-1 (ACV-resistant strain), HSV-2, and VZV. Vero cells and MeWo cells. Six-week-old BALB/c mice. | Houttuynoid A strongly repressed the infectivity of HSV-1, HSV-2, and VZV, with IC50 values of 23.50, 36.38, and 23.48 µM, respectively. | Blocking HSV-1 membrane fusion induced by viral glycoproteins (in vitro). Inhibition of HSV-1 multiplication by preventing lesion formation in a HSV-1 infection mouse model (in vivo). | [103] |
| In vitro. Plaque-formation assay and CellTiter-Glo Luminescent cell viability assay. HSV-1 (F strain). Vero cells. | houttuynoid M and houttuynoid A exposed anti-HSV-1 actions, with IC50 values of 17.72 and 12.42 µM, respectively. | No mechanism of action was defined. | [104] |
| Type of Study, Method, Virus, and Cells | Results (Compound, Concentration, or Dose) | Mechanisms | Reference |
|---|---|---|---|
| In vitro. Plaque-reduction assay and multiple biochemical analyses. HCMV–Towne. HFF cells. | Quercetin and isoquercitrin potently hindered the replication of HCMV with IC50 values of 5.9 and 1.9 µg/mL, respectively. | Inhibition of HCMV-IE gene expression. Suppression of the transcript levels of HCMV UL122 (IE), UL44 (E) and UL83 (L). Inhibition of MIEP activation by interfering with the JNK pathway. | [102] |
| In vitro. mCherry (a marker of infection), eGFP (a marker of late viral replication) fluorescence assays, and various biochemical analyses. Recombinant HCMV (ganciclovir-resistant strain) TB40/EmCherry-UL99eGFP. NuFF-1 cells. | Treatment of HCMV-infected NuFF-1 cells with deguelin at high (moi = 1.0) or low (moi = 0.01) multiplicities potently suppressed the HCMV lytic replication, with IC50 values of 55.8 and 23.4 nM, respectively. | Deguelin (250 nM) effectively repressed E and L viral gene transcriptions and reduced the expressions of IE2-86 and IE2-60 proteins. | [114] |
| In vitro. Plaque-reduction assay coupled with multiple biochemical tests. HCMV–Towne. HEL fibroblast cells. | Tricin suppressed the replication and infection of HCMV at a concentration of 10 µM. | Reduction of IE1 and UL54 (encoding DNA polymerase) genes expression. Inhibition of CCL2-CCR2 axis expressions in the HCMV replication cycle. | [115] |
| In vitro. Plaque assay combined with various biochemical analyses. HCMV–Towne. HEL fibroblast cells. | Treatment with tricin (10 µM) showed considerable inhibition of HCMV replication. | Inhibition of IE1 and UL54 gene expressions. Suppression of CCL5 protein expression. | [116] |
| In vitro and in silico. Plaque-reduction assay and various biochemical and molecular docking analyses. HCMV–Towne. HEL fibroblast cells. | Tricin and flavopiridol (synthetic flavonoid and standard inhibitor of CDK) exhibited notable anti-HCMV properties, with EC50 values of 2.09 µM and 15.8 nM, respectively. | In vitro (tricin and flavopiridol repressed the activity of CDK9, with IC50 values of 1.38 µM and 8.20 nM, respectively). The anti-CDK9 activity of tricin is related to the phosphorylation of the carboxy-terminal domain of RNA polymerase II. In silico (tricin was found to bind to the ATP-binding site of CDK9). | [118] |
| In vitro and in silico. Plaque-reduction assay and multiple biochemical and molecular docking simulations assays. HCMV–Towne. HEL fibroblast cells. | The anti-HCMV activities of tricin and 6F-tricin were determined, with EC50 values of 54.3 and 0.13 nM, respectively. | In silico (tricin and 6F-tricin were detected to bind to the ATP-binding site of CDK9, and significant binding affinity was observed with 6F-tricin). | [119] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Šudomová, M.; Berchová-Bímová, K.; Mazurakova, A.; Šamec, D.; Kubatka, P.; Hassan, S.T.S. Flavonoids Target Human Herpesviruses That Infect the Nervous System: Mechanisms of Action and Therapeutic Insights. Viruses 2022, 14, 592. https://doi.org/10.3390/v14030592
Šudomová M, Berchová-Bímová K, Mazurakova A, Šamec D, Kubatka P, Hassan STS. Flavonoids Target Human Herpesviruses That Infect the Nervous System: Mechanisms of Action and Therapeutic Insights. Viruses. 2022; 14(3):592. https://doi.org/10.3390/v14030592
Chicago/Turabian StyleŠudomová, Miroslava, Kateřina Berchová-Bímová, Alena Mazurakova, Dunja Šamec, Peter Kubatka, and Sherif T. S. Hassan. 2022. "Flavonoids Target Human Herpesviruses That Infect the Nervous System: Mechanisms of Action and Therapeutic Insights" Viruses 14, no. 3: 592. https://doi.org/10.3390/v14030592
APA StyleŠudomová, M., Berchová-Bímová, K., Mazurakova, A., Šamec, D., Kubatka, P., & Hassan, S. T. S. (2022). Flavonoids Target Human Herpesviruses That Infect the Nervous System: Mechanisms of Action and Therapeutic Insights. Viruses, 14(3), 592. https://doi.org/10.3390/v14030592