
Isolation and Genomic Characterization of SARS-CoV-2 Omicron Variant Obtained from Human Clinical Specimens

 ,
,  , , , ,
, , , , {kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Clinical Specimens
2.2. In Vitro Virus Isolation Approach
2.3. In Vivo Virus Isolation Approach
2.4. Next-Generation Sequencing and Data Analysis
3. Results
3.1. Characterization of the Circulating SARS-CoV-2 Variants
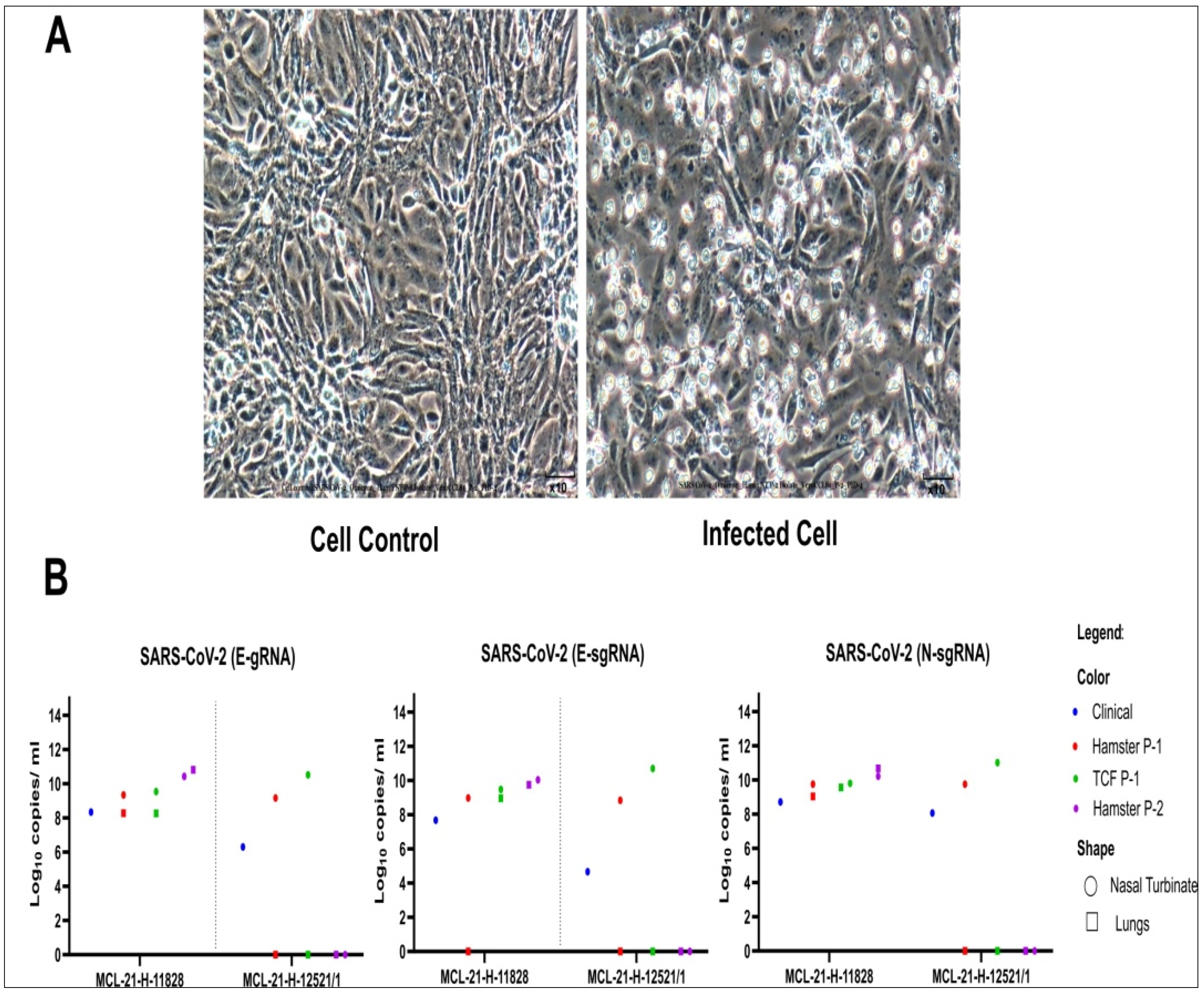
3.2. Virus Isolation with Syrian Hamster Was Successful than on Vero CCL-81 Cells
3.3. Subsequent Virus Isolation on Vero CCL-81 Cells
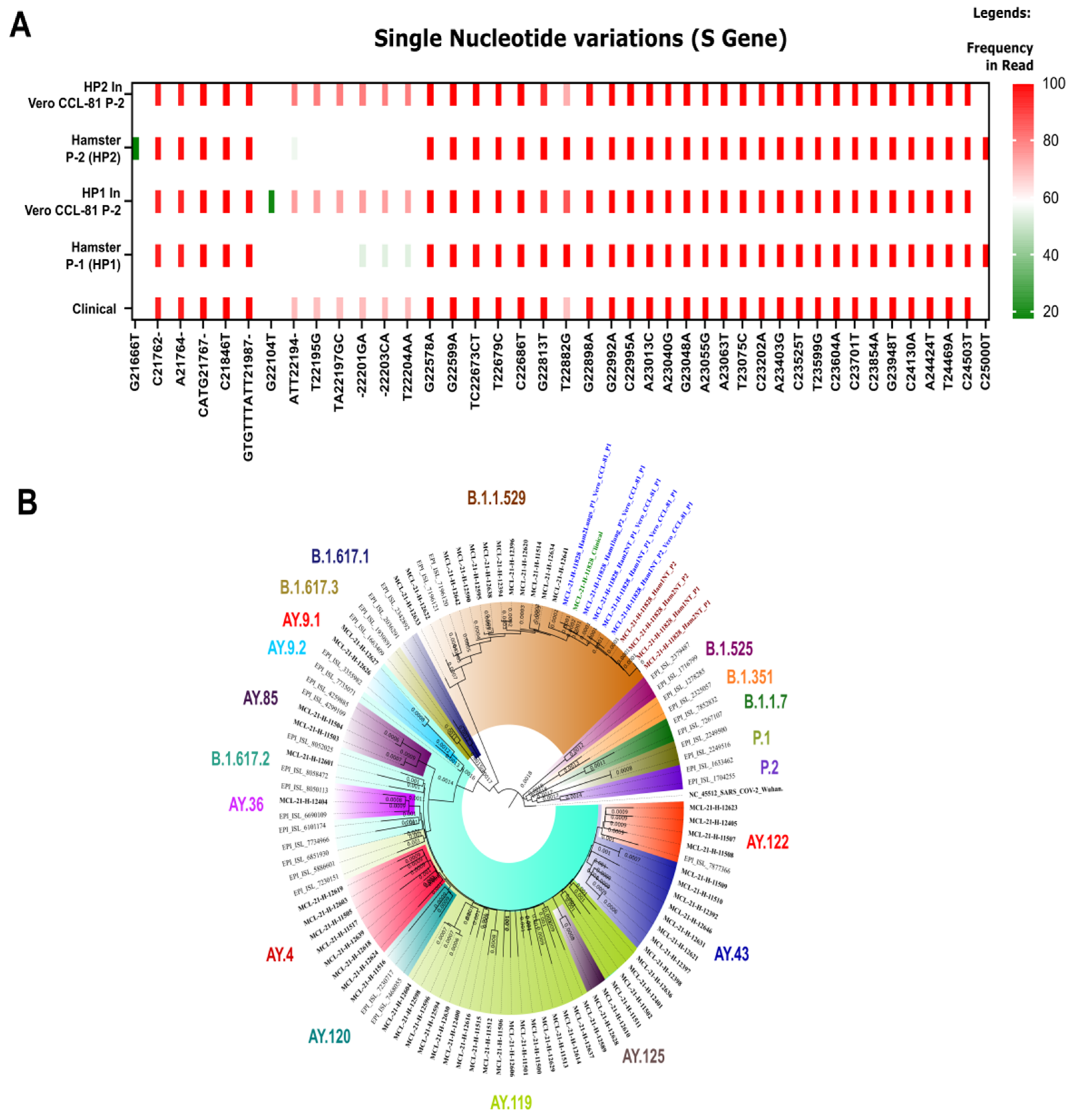
3.4. Genomic Analysis of Clinical Specimens and SARS-CoV-2 Isolates
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Classification of Omicron (B.1.1.529): SARS-CoV-2 Variant of Concern. Available online: https://www.who.int/news/item/26-11-2021-classification-of-omicron-(b.1.1.529)-sars-cov-2-variant-of-concern (accessed on 21 December 2021).
- GISAID. Tracking of Variants. Omicron. Available online: https://www.gisaid.org/hcov19-variants/ (accessed on 21 December 2021).
- Covariants. Variant: 21K (Omicron). CoVariants. Available online: https://covariants.org/variants/21K.Omicron (accessed on 21 December 2021).
- Wurtz, N.; Penant, G.; Jardot, P.; Duclos, N.; La Scola, B. Culture of SARS-CoV-2 in a panel of laboratory cell lines, permissivity, and differences in growth profile. Eur. J. Clin. Microbiol. Infect. Dis. 2021, 40, 477–484. [Google Scholar] [CrossRef] [PubMed]
- Willett, B.J.; Grove, J.; MacLean, O.; Wilkie, C.; Logan, N.; De Lorenzo, G.; Furnon, W.; Scott, S.; Manali, M.; Szemiel, A.; et al. The hyper-transmissible SARS-CoV-2 Omicron variant exhibits significant antigenic change, vaccine escape and a switch in cell entry mechanism. MedRxiv. 2022, preprint. [Google Scholar] [CrossRef]
- Peacock, T.P.; Brown, J.C.; Zhou, J.; Thakur, N.; Newman, J.; Kugathasan, R.; Sukhova, K.; Kaforou, M.; Bailey, D.; Barclay, W.S. The SARS-CoV-2 variant, Omicron, shows rapid replication in human primary nasal epithelial cultures and efficiently uses the endosomal route of entry. BioRxiv. 2022, preprint. [Google Scholar] [CrossRef]
- Shuai, H.; Chan, J.F.; Hu, B.; Chai, Y.; Yuen, T.T.; Yin, F.; Huang, X.; Yoon, C.; Hu, J.C.; Liu, H.; et al. Attenuated replication and pathogenicity of SARS-CoV-2 B. 1.1. 529 Omicron. Nature 2022, 1. [Google Scholar] [CrossRef]
- Cele, S.; Jackson, L.; Khoury, D.S.; Khan, K.; Moyo-Gwete, T.; Tegally, H.; San, J.E.; Cromer, D.; Scheepers, C.; Amoako, D.; et al. SARS-CoV-2 Omicron has extensive but incomplete escape of Pfizer BNT162b2 elicited neutralization and requires ACE2 for infection. MedRxiv. 2021, preprint. [Google Scholar] [CrossRef]
- Sarkale, P.; Patil, S.; Yadav, P.D.; Nyayanit, D.A.; Sapkal, G.; Baradkar, S.; Lakra, R.; Shete-Aich, A.; Prasad, S.; Basu, A.; et al. First isolation of SARS-CoV-2 from clinical samples in India. Indian J. Med. Res. 2020, 151, 244. [Google Scholar] [PubMed]
- Choudhary, M.L.; Vipat, V.; Jadhav, S.; Basu, A.; Cherian, S.; Abraham, P.; Potdar, V.A. Development of in vitro transcribed RNA as positive control for laboratory diagnosis of SARS-CoV-2 in India. Indian J. Med. Res. 2020, 151, 251. [Google Scholar] [PubMed]
- Yadav, P.D.; Nyayanit, D.A.; Shete, A.M.; Jain, S.; Majumdar, T.P.; Chaubal, G.Y.; Shil, P.; Kore, P.M.; Mourya, D.T. Complete genome sequencing of Kaisodi virus isolated from ticks in India belonging to Phlebovirus genus, family Phenuiviridae. Ticks Tick Borne Dis. 2019, 10, 23–33. [Google Scholar] [CrossRef]
- Tandel, D.; Gupta, D.; Sah, V.; Harshan, K.H. N440K variant of SARS-CoV-2 has Higher Infectious Fitness. bioRxiv. 2021. preprint. [Google Scholar] [CrossRef]
- Kullappan, M.; Mary, U.; Ambrose, J.M.; Veeraraghavan, V.P.; Surapaneni, K.M. Elucidating the role of N440K mutation in SARS-CoV-2 spike–ACE-2 binding affinity and COVID-19 severity by virtual screening, molecular docking and dynamics approach. J. Biomol. Struct. Dyn. 2021, 8, 1–8. [Google Scholar] [CrossRef] [PubMed]
- VanBlargan, L.A.; Errico, J.M.; Halfmann, P.J.; Zost, S.J.; Crowe, J.E.; Purcell, L.A.; Kawaoka, Y.; Corti, D.; Fremont, D.H.; Diamond, M.S. An infectious SARS-CoV-2 B. 1.1. 529 Omicron virus escapes neutralization by therapeutic monoclonal antibodies. Nat. Med. 2022, 1–6. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yadav, P.D.; Gupta, N.; Potdar, V.; Mohandas, S.; Sahay, R.R.; Sarkale, P.; Shete, A.M.; Razdan, A.; Patil, D.Y.; Nyayanit, D.A.; et al. Isolation and Genomic Characterization of SARS-CoV-2 Omicron Variant Obtained from Human Clinical Specimens. Viruses 2022, 14, 461. https://doi.org/10.3390/v14030461
Yadav PD, Gupta N, Potdar V, Mohandas S, Sahay RR, Sarkale P, Shete AM, Razdan A, Patil DY, Nyayanit DA, et al. Isolation and Genomic Characterization of SARS-CoV-2 Omicron Variant Obtained from Human Clinical Specimens. Viruses. 2022; 14(3):461. https://doi.org/10.3390/v14030461
Chicago/Turabian StyleYadav, Pragya D., Nivedita Gupta, Varsha Potdar, Sreelekshmy Mohandas, Rima R. Sahay, Prasad Sarkale, Anita M. Shete, Alpana Razdan, Deepak Y. Patil, Dimpal A. Nyayanit, and et al. 2022. "Isolation and Genomic Characterization of SARS-CoV-2 Omicron Variant Obtained from Human Clinical Specimens" Viruses 14, no. 3: 461. https://doi.org/10.3390/v14030461
APA StyleYadav, P. D., Gupta, N., Potdar, V., Mohandas, S., Sahay, R. R., Sarkale, P., Shete, A. M., Razdan, A., Patil, D. Y., Nyayanit, D. A., Joshi, Y., Patil, S., Majumdar, T., Dighe, H., Malhotra, B., Shastri, J., & Abraham, P. (2022). Isolation and Genomic Characterization of SARS-CoV-2 Omicron Variant Obtained from Human Clinical Specimens. Viruses, 14(3), 461. https://doi.org/10.3390/v14030461