
Evolution of TRIM5 and TRIM22 in Bats Reveals a Complex Duplication Process

 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
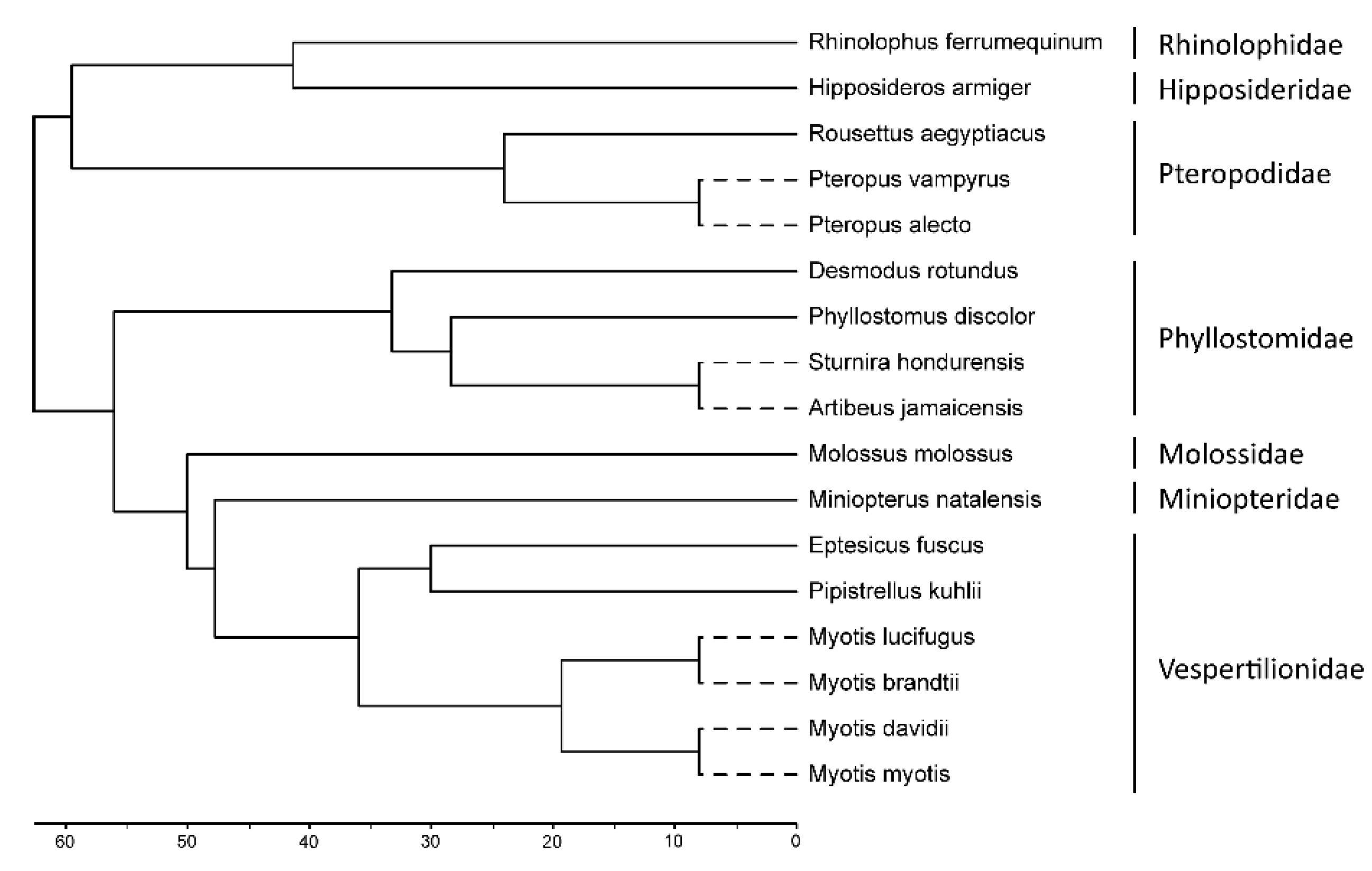
2.1. Phylogenetic Analyses
2.2. Molecular Evolutionary Analyses
3. Results
3.1. Data Retrieving
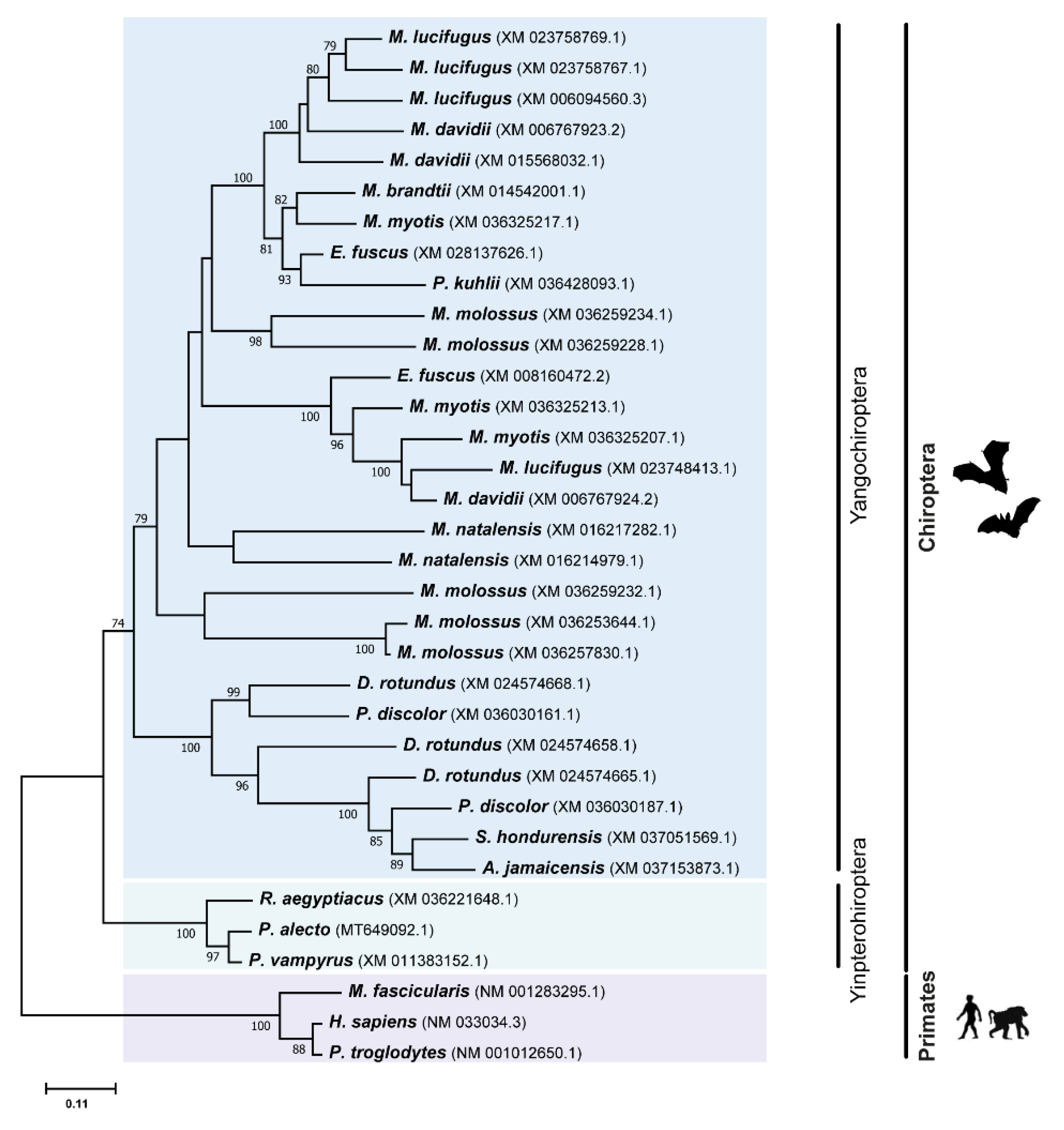
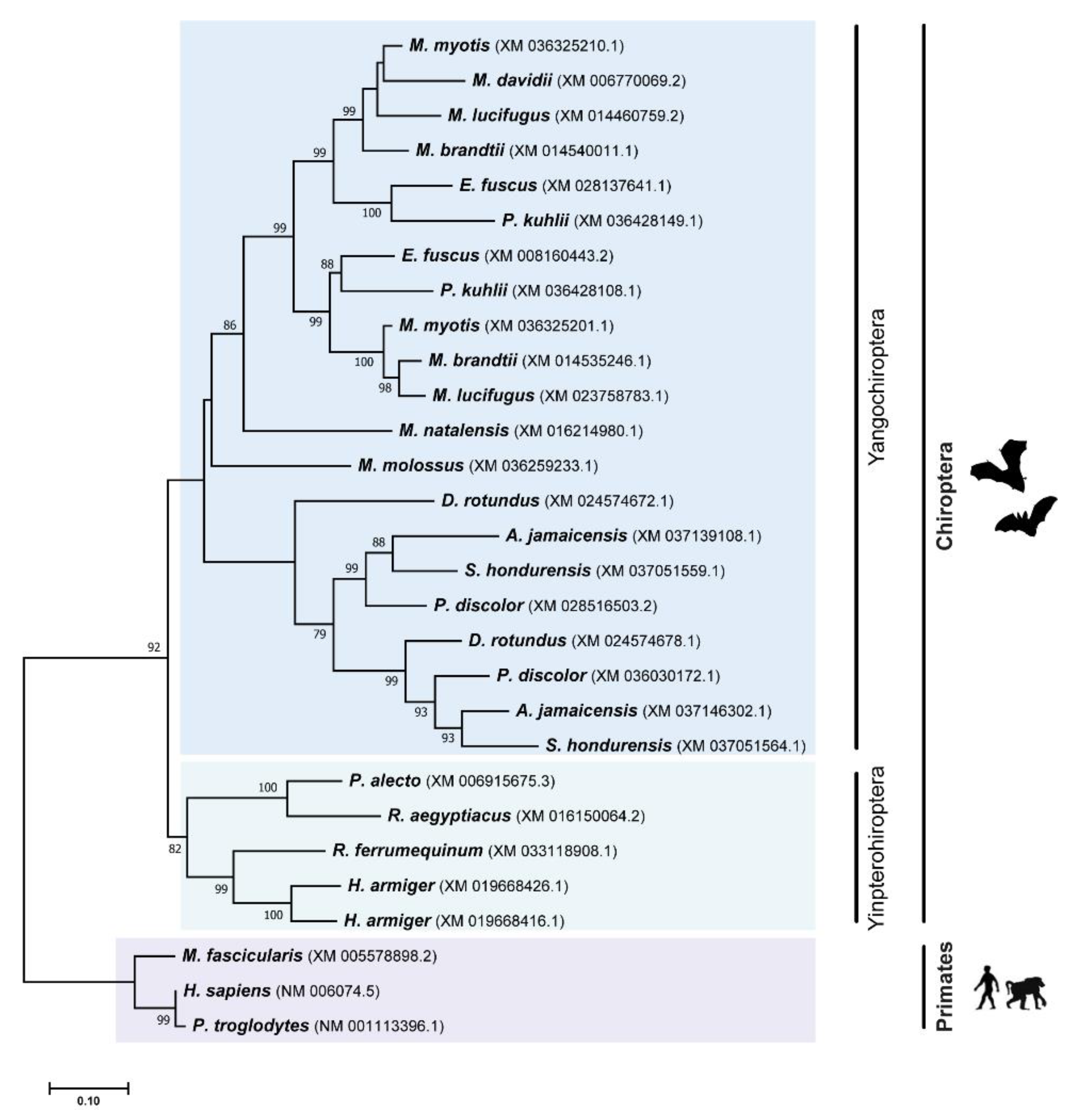
3.2. Phylogenetic Analysis Suggest Several Events of Duplication for TRIM5 and TRIM22 Genes
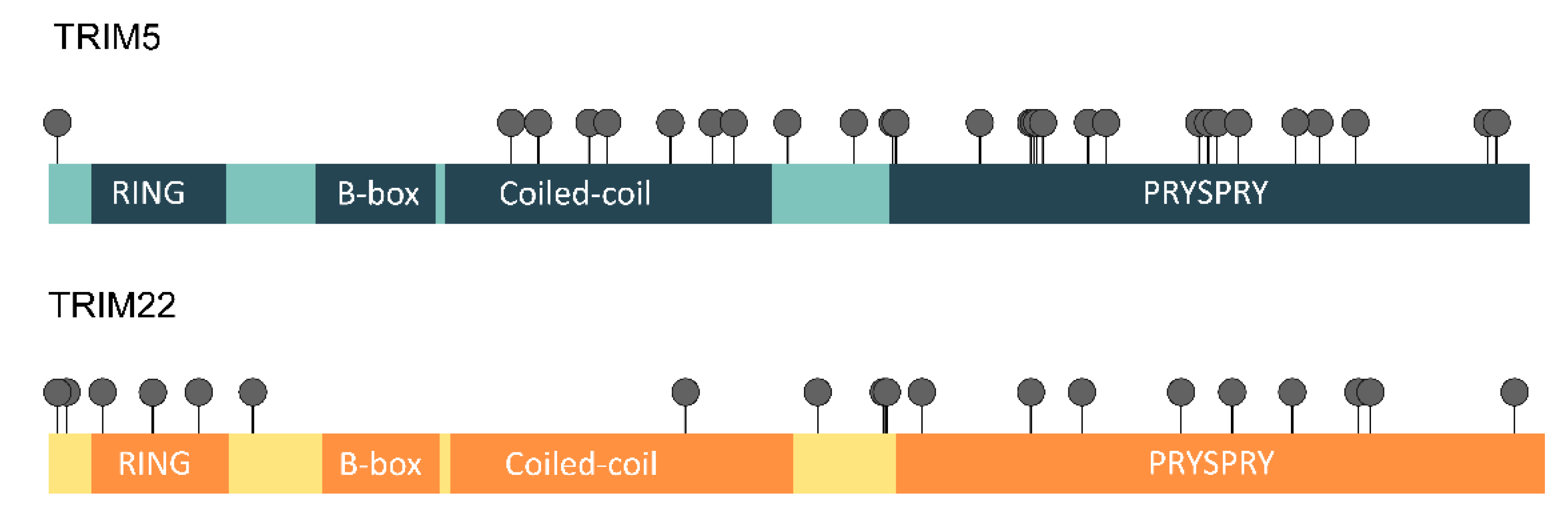
3.3. Evidence of Positive Selection Acting on TRIM5 and TRIM22 Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Duggal, N.K.; Emerman, M. Evolutionary conflicts between viruses and restriction factors shape immunity. Nat. Rev. Immunol. 2012, 12, 687–695. [Google Scholar] [CrossRef] [PubMed]
- Diehl, W.E.; Patel, N.; Halm, K.; Johnson, W.E. Tracking interspecies transmission and long-term evolution of an ancient retrovirus using the genomes of modern mammals. Elife 2016, 5, e12704. [Google Scholar] [CrossRef] [PubMed]
- Yan, N.; Chen, Z.J. Intrinsic antiviral immunity. Nat. Immunol. 2012, 13, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Morrison, J.H.; Miller, C.; Bankers, L.; Crameri, G.; Wang, L.-F.; Poeschla, E.M. A potent postentry restriction to primate lentiviruses in a yinpterochiropteran bat. MBio 2020, 11, e01854-20. [Google Scholar] [CrossRef] [PubMed]
- Jáuregui, P.; Crespo, H.; Glaria, I.; Luján, L.; Contreras, A.; Rosati, S.; de Andrés, D.; Amorena, B.; Towers, G.J.; Reina, R. Ovine TRIM5α Can restrict visna/maedi Virus. J. Virol. 2012, 86, 9504–9509. [Google Scholar] [CrossRef] [PubMed]
- Besnier, C.; Takeuchi, Y.; Towers, G. Restriction of lentivirus in monkeys. Proc. Natl. Acad. Sci. USA 2002, 99, 11920–11925. [Google Scholar] [CrossRef]
- Stremlau, M.; Owens, C.M.; Perron, M.J.; Kiessling, M.; Autissier, P.; Sodroski, J. The cytoplasmic body component TRIM5alpha Restricts HIV-1 infection in old world monkeys. Nature 2004, 427, 848–853. [Google Scholar] [CrossRef]
- Han, K.; Lou, D.I.; Sawyer, S.L. Identification of a genomic reservoir for new TRIM genes in primate genomes. PLoS Genet. 2011, 7, e1002388. [Google Scholar] [CrossRef]
- Nisole, S.; Stoye, J.P.; Saïb, A. TRIM family proteins: Retroviral restriction and antiviral defence. Nat. Rev. Microbiol. 2005, 3, 799–808. [Google Scholar] [CrossRef]
- Johnson, W.E.; Sawyer, S.L. Molecular evolution of the antiretroviral TRIM5 Gene. Immunogenetics 2009, 61, 163–176. [Google Scholar] [CrossRef]
- Song, B.; Gold, B.; O’Huigin, C.; Javanbakht, H.; Li, X.; Stremlau, M.; Winkler, C.; Dean, M.; Sodroski, J. The B30.2(SPRY) domain of the retroviral restriction factor TRIM5alpha exhibits lineage-specific length and sequence variation in primates. J. Virol. 2005, 79, 6111–6121. [Google Scholar] [CrossRef] [PubMed]
- D’Cruz, A.A.; Babon, J.J.; Norton, R.S.; Nicola, N.A.; Nicholson, S.E. Structure and function of the SPRY/B30.2 domain proteins involved in innate immunity. Protein Sci. 2013, 22, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Javanbakht, H.; Diaz-Griffero, F.; Stremlau, M.; Si, Z.; Sodroski, J. The contribution of RING and B-Box 2 domains to retroviral restriction mediated by monkey TRIM5alpha. J. Biol. Chem. 2005, 280, 26933–26940. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Griffero, F.; Perron, M.; McGee-Estrada, K.; Hanna, R.; Maillard, P.V.; Trono, D.; Sodroski, J. A Human TRIM5alpha B30.2/SPRY domain mutant gains the ability to restrict and prematurely uncoat B-tropic murine leukemia virus. Virology 2008, 378, 233–242. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Biris, N.; Yang, Y.; Taylor, A.B.; Tomashevski, A.; Guo, M.; Hart, P.J.; Diaz-Griffero, F.; Ivanov, D.N. Structure of the rhesus monkey TRIM5α PRYSPRY domain, the HIV capsid recognition module. Proc. Natl. Acad. Sci. USA 2012, 109, 13278–13283. [Google Scholar] [CrossRef] [PubMed]
- James, L.C.; Keeble, A.H.; Khan, Z.; Rhodes, D.A.; Trowsdale, J. Structural basis for PRYSPRY-mediated tripartite motif (TRIM) protein function. Proc. Natl. Acad. Sci. USA 2007, 104, 6200–6205. [Google Scholar] [CrossRef]
- Sawyer, S.L.; Emerman, M.; Malik, H.S. Discordant evolution of the adjacent antiretroviral genes TRIM22 and TRIM5 in mammals. PLoS Pathog. 2007, 3, e197. [Google Scholar] [CrossRef]
- Ribeiro, C.M.S.; Sarrami-Forooshani, R.; Setiawan, L.C.; Zijlstra-Willems, E.M.; van Hamme, J.L.; Tigchelaar, W.; van der Wel, N.N.; Kootstra, N.A.; Gringhuis, S.I.; Geijtenbeek, T.B.H. Receptor usage dictates HIV-1 restriction by human TRIM5α in dendritic cell subsets. Nature 2016, 540, 448–452. [Google Scholar] [CrossRef]
- Kim, K.; Dauphin, A.; Komurlu, S.; McCauley, S.M.; Yurkovetskiy, L.; Carbone, C.; Diehl, W.E.; Strambio-De-Castillia, C.; Campbell, E.M.; Luban, J. Cyclophilin A protects HIV-1 from restriction by human TRIM5α. Nat. Microbiol. 2019, 4, 2044–2051. [Google Scholar] [CrossRef]
- Selyutina, A.; Persaud, M.; Simons, L.M.; Bulnes-Ramos, A.; Buffone, C.; Martinez-Lopez, A.; Scoca, V.; Di Nunzio, F.; Hiatt, J.; Marson, A.; et al. Cyclophilin a prevents HIV-1 restriction in lymphocytes by blocking human TRIM5α binding to the viral Core. Cell Rep. 2020, 30, 3766–3777.e6. [Google Scholar] [CrossRef]
- Dufour, C.; Claudel, A.; Joubarne, N.; Merindol, N.; Maisonnet, T.; Masroori, N.; Plourde, M.B.; Berthoux, L. Editing of the human TRIM5 gene to introduce mutations with the potential to inhibit HIV-1. PLoS ONE 2018, 13, e0191709. [Google Scholar] [CrossRef] [PubMed]
- Yap, M.W.; Nisole, S.; Stoye, J.P. A single amino acid change in the SPRY domain of human Trim5alpha leads to HIV-1 restriction. Curr. Biol. 2005, 15, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Di Pietro, A.; Kajaste-Rudnitski, A.; Oteiza, A.; Nicora, L.; Towers, G.J.; Mechti, N.; Vicenzi, E. TRIM22 inhibits influenza A Virus infection by targeting the viral nucleoprotein for degradation. J. Virol. 2013, 87, 4523–4533. [Google Scholar] [CrossRef]
- Gao, B.; Duan, Z.; Xu, W.; Xiong, S. Tripartite motif-containing 22 inhibits the activity of hepatitis B virus core promoter, which is dependent on nuclear-located RING domain. Hepatology 2009, 50, 424–433. [Google Scholar] [CrossRef]
- Jing, H.; Tao, R.; Dong, N.; Cao, S.; Sun, Y.; Ke, W.; Li, Y.; Wang, J.; Zhang, Y.; Huang, H.; et al. Nuclear localization signal in TRIM22 is essential for inhibition of type 2 porcine reproductive and respiratory syndrome virus replication in MARC-145 cells. Virus Genes 2019, 55, 660–672. [Google Scholar] [CrossRef]
- Hattlmann, C.J.; Kelly, J.N.; Barr, S.D. TRIM22: A diverse and dynamic antiviral protein. Mol. Biol. Int. 2012, 2012, 153415. [Google Scholar] [CrossRef] [PubMed]
- Ohainle, M.; Kim, K.; Komurlu Keceli, S.; Felton, A.; Campbell, E.; Luban, J.; Emerman, M. TRIM34 restricts HIV-1 and SIV capsids in a TRIM5α-dependent manner. PLoS Pathog. 2020, 16, e1008507. [Google Scholar] [CrossRef]
- Bharaj, P.; Wang, Y.E.; Dawes, B.E.; Yun, T.E.; Park, A.; Yen, B.; Basler, C.F.; Freiberg, A.N.; Lee, B.; Rajsbaum, R. The matrix protein of Nipah virus targets the E3-Ubiquitin ligase TRIM6 to inhibit the IKKε kinase-mediated Type-I IFN antiviral response. PLoS Pathog. 2016, 12, e1005880. [Google Scholar] [CrossRef]
- Tareen, S.U.; Sawyer, S.L.; Malik, H.S.; Emerman, M. An expanded clade of rodent Trim5 genes. Virology 2009, 385, 473–483. [Google Scholar] [CrossRef]
- Si, Z.; Vandegraaff, N.; O’huigin, C.; Song, B.; Yuan, W.; Xu, C.; Perron, M.; Li, X.; Marasco, W.A.; Engelman, A.; et al. Evolution of a cytoplasmic tripartite motif (TRIM) protein in cows that restricts retroviral infection. Proc. Natl. Acad. Sci. USA 2006, 103, 7454–7459. [Google Scholar] [CrossRef]
- Sawyer, S.L.; Wu, L.I.; Emerman, M.; Malik, H.S. Positive selection of primate TRIM5alpha identifies a critical species-specific retroviral restriction domain. Proc. Natl. Acad. Sci. USA 2005, 102, 2832–2837. [Google Scholar] [CrossRef] [PubMed]
- Águeda-Pinto, A.; Lemos de Matos, A.; Pinheiro, A.; Neves, F.; de Sousa-Pereira, P.; Esteves, P.J. Not so unique to primates: The independent adaptive evolution of TRIM5 in lagomorpha lineage. PLoS ONE 2019, 14, e0226202. [Google Scholar] [CrossRef] [PubMed]
- Boso, G.; Shaffer, E.; Liu, Q.; Cavanna, K.; Buckler-White, A.; Kozak, C.A. Evolution of the rodent Trim5 cluster is marked by divergent paralogous expansions and independent acquisitions of TrimCyp fusions. Sci. Rep. 2019, 9, 11263. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.J.; Rabosky, D.L. Speciation dynamics during the global radiation of extant bats. Evolution 2015, 69, 1528–1545. [Google Scholar] [CrossRef]
- Amador, L.I.; Leticia Moyers Arévalo, R.; Almeida, F.C.; Catalano, S.A.; Giannini, N.P. Bat systematics in the light of unconstrained analyses of a comprehensive molecular supermatrix. J. Mamm. Evol. 2018, 25, 37–70. [Google Scholar] [CrossRef]
- Agnarsson, I.; Zambrana-Torrelio, C.M.; Flores-Saldana, N.P.; May-Collado, L.J. A time-calibrated species-level phylogeny of bats (Chiroptera, Mammalia). PLoS Curr. 2011, 3, RRN1212. [Google Scholar] [CrossRef]
- Calisher, C.H.; Childs, J.E.; Field, H.E.; Holmes, K.V.; Schountz, T. Bats: Important reservoir hosts of emerging viruses. Clin. Microbiol. Rev. 2006, 19, 531–545. [Google Scholar] [CrossRef]
- Andersen, K.G.; Rambaut, A.; Lipkin, W.I.; Holmes, E.C.; Garry, R.F. The proximal origin of SARS-CoV-2. Nat. Med. 2020, 26, 450–452. [Google Scholar] [CrossRef]
- Seyran, M.; Pizzol, D.; Adadi, P.; El-Aziz, T.M.A.; Hassan, S.S.; Soares, A.; Kandimalla, R.; Lundstrom, K.; Tambuwala, M.; Aljabali, A.A.A.; et al. Questions concerning the proximal origin of SARS-CoV-2. J. Med. Virol. 2021, 93, 1204–1206. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X Version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. ProtTest 3: Fast selection of best-fit models of protein evolution. Bioinformatics 2011, 27, 1164–1165. [Google Scholar] [CrossRef] [PubMed]
- Pond, S.L.K.; Frost, S.D.W. Datamonkey: Rapid detection of selective pressure on individual sites of codon alignments. Bioinformatics 2005, 21, 2531–2533. [Google Scholar] [CrossRef]
- Yang, Z. PAML 4: Phylogenetic analysis by maximum likelihood. Mol. Biol. Evol. 2007, 24, 1586–1591. [Google Scholar] [CrossRef]
- Kosakovsky Pond, S.L.; Frost, S.D.W. Not so different after all: A comparison of methods for detecting amino acid sites under selection. Mol. Biol. Evol. 2005, 22, 1208–1222. [Google Scholar] [CrossRef]
- Murrell, B.; Wertheim, J.O.; Moola, S.; Weighill, T.; Scheffler, K.; Kosakovsky Pond, S.L. Detecting individual sites subject to episodic diversifying selection. PLoS Genet. 2012, 8, e1002764. [Google Scholar] [CrossRef] [PubMed]
- Murrell, B.; Moola, S.; Mabona, A.; Weighill, T.; Sheward, D.; Kosakovsky Pond, S.L.; Scheffler, K. FUBAR: A fast, unconstrained bayesian approximation for inferring selection. Mol. Biol. Evol. 2013, 30, 1196–1205. [Google Scholar] [CrossRef]
- Águeda-Pinto, A.; Alves, L.Q.; Neves, F.; McFadden, G.; Jacobs, B.L.; Castro, L.F.C.; Rahman, M.M.; Esteves, P.J. Convergent loss of the necroptosis pathway in disparate mammalian lineages shapes viruses countermeasures. Front. Immunol. 2021, 12, 747737. [Google Scholar] [CrossRef]
- Pinheiro, A.; Águeda-Pinto, A.; Melo-Ferreira, J.; Neves, F.; Abrantes, J.; Esteves, P.J. Analysis of substitution rates showed That TLR5 is evolving at different rates among mammalian groups. BMC Evol. Biol. 2019, 19, 221. [Google Scholar] [CrossRef]
- Miller-Butterworth, C.M.; Murphy, W.J.; O’Brien, S.J.; Jacobs, D.S.; Springer, M.S.; Teeling, E.C. A family matter: Conclusive resolution of the taxonomic position of the long-fingered bats, miniopterus. Mol. Biol. Evol. 2007, 24, 1553–1561. [Google Scholar] [CrossRef]
- Kosiol, C.; Vinař, T.; da Fonseca, R.R.; Hubisz, M.J.; Bustamante, C.D.; Nielsen, R.; Siepel, A. Patterns of positive selection in six mammalian genomes. PLoS Genet. 2008, 4, e1000144. [Google Scholar] [CrossRef] [PubMed]
- De Matos, A.L.; van der Loo, W.; Areal, H.; Lanning, D.K.; Esteves, P.J. Study of sylvilagus rabbit TRIM5α species-specific domain: How ancient endoviruses could have shaped the antiviral repertoire in lagomorpha. BMC Evol. Biol. 2011, 11, 294. [Google Scholar] [CrossRef] [PubMed]
- Tong, S.; Li, Y.; Rivailler, P.; Conrardy, C.; Castillo, D.A.A.; Chen, L.-M.; Recuenco, S.; Ellison, J.A.; Davis, C.T.; York, I.A.; et al. A distinct lineage of influenza a virus from bats. Proc. Natl. Acad. Sci. USA 2012, 109, 4269–4274. [Google Scholar] [CrossRef] [PubMed]
- Tong, S.; Zhu, X.; Li, Y.; Shi, M.; Zhang, J.; Bourgeois, M.; Yang, H.; Chen, X.; Recuenco, S.; Gomez, J.; et al. New world bats harbor diverse influenza a viruses. PLoS Pathog. 2013, 9, e1003657. [Google Scholar] [CrossRef]
- Campos, A.C.A.; Góes, L.G.B.; Moreira-Soto, A.; de Carvalho, C.; Ambar, G.; Sander, A.-L.; Fischer, C.; Ruckert da Rosa, A.; Cardoso de Oliveira, D.; Kataoka, A.P.G.; et al. Bat influenza A(HL18NL11) virus in fruit bats, Brazil. Emerg. Infect. Dis. 2019, 25, 333–337. [Google Scholar] [CrossRef]
- Ciminski, K.; Thamamongood, T.; Zimmer, G.; Schwemmle, M. Novel insights into bat influenza a viruses. J. Gen. Virol. 2017, 98, 2393–2400. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Family | Species Name | TRIM6 | TRIM34 | TRIM5 | TRIM22 |
|---|---|---|---|---|---|---|
| Yangochiroptera | Vespertilionidae | Myotis brandtii | XM_005860820.1 | XM_005860818.2 | XM_014542001.1 | XM_014540011.1 XM_014535246.1 |
| Myotis myotis | XM_036325208.1 | XM_036325206.1 | XM_036325207.1 XM_036325213.1 XM_036325217.1 | XM_036325201.1 XM_036325210.1 | ||
| Myotis lucifugus | XM_006094555.2 | XM_023758764.1 | XM_023758769.1 XM_023758767.1 XM_023748413.1 | XM_014460759.2 XM_023758783.1 | ||
| Myotis davidii | XM_015560087.1 | XM_015560147.1 | XM_006767923.2 XM_015568032.1 XM_006767924.2 | XM_006770069.2 | ||
| Pipistrellus kuhlii | XM_036428111.1 | XM_036428091.1 | XM_036428093.1 | XM_036428149.1 XM_036428108.1 | ||
| Eptesicus fuscus | XM_008160441.2:134-1606 | XM_008160439.2:174-1766 | XM_028137626.1 XM_008160472.2 | XM_028137641.1 XM_008160443.2 | ||
| Miniopteridae | Miniopterus natalensis | XM_016215031.1:626-2098 | XM_016215034.1:166-1623 | XM_016214979.1 XM_016217282.1 | XM_016214980.1 | |
| Molossidae | Molossus molossus | XM_036259561.1 | XM_036258280.1 | XM_036259234.1 XM_036259232.1 XM_036257830.1 XM_036253644.1 XM_036259228.1 | XM_036259233.1 | |
| Phyllostomidae | Sturnira hondurensis | XM_037037747.1 | XM_037051569.1 | XM_037051564.1 XM_037051559.1 | ||
| Desmodus rotundus | XM_024574677.1 | XM_024574667.1 | XM_024574668.1 XM_024574658.1 | XM_024574672.1 XM_024574678.1 | ||
| Phyllostomus discolor | XM_028515503.2 | XM_036030161.1 XM_036030187.1 | XM_036030172.1 XM_028516503.2 | |||
| Artibeus jamaicensis | XM_037146300.1 | XM_037146301.1 | XM_037153873.1 | XM_037146302.1 XM_037139108.1 | ||
| Yinpterochiroptera | Hipposideridae | Hipposideros armiger | XM_019668446.1 | XM_019668445.1 | XM_019668416.1 XM_019668426.1 | |
| Rhinolophidae | Rhinolophus ferrumequinum | XM_033120756.1 | XM_033120754.1 | XM_033118908.1 | ||
| Pteropodidae | Pteropus alecto | XM_006915678.1 | XM_015592357.2 | MT649092.1 | XM_006915675.3 | |
| Pteropus vampyrus | XM_011383131.1 | XM_011383134.2 | XM_011383152.1 | |||
| Rousettus aegyptiacus | XM_016126836.2 | XM_036221642.1 | XM_036221648.1 | XM_016150064.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernandes, A.P.; Águeda-Pinto, A.; Pinheiro, A.; Rebelo, H.; Esteves, P.J. Evolution of TRIM5 and TRIM22 in Bats Reveals a Complex Duplication Process. Viruses 2022, 14, 345. https://doi.org/10.3390/v14020345
Fernandes AP, Águeda-Pinto A, Pinheiro A, Rebelo H, Esteves PJ. Evolution of TRIM5 and TRIM22 in Bats Reveals a Complex Duplication Process. Viruses. 2022; 14(2):345. https://doi.org/10.3390/v14020345
Chicago/Turabian StyleFernandes, Alexandre P., Ana Águeda-Pinto, Ana Pinheiro, Hugo Rebelo, and Pedro J. Esteves. 2022. "Evolution of TRIM5 and TRIM22 in Bats Reveals a Complex Duplication Process" Viruses 14, no. 2: 345. https://doi.org/10.3390/v14020345
APA StyleFernandes, A. P., Águeda-Pinto, A., Pinheiro, A., Rebelo, H., & Esteves, P. J. (2022). Evolution of TRIM5 and TRIM22 in Bats Reveals a Complex Duplication Process. Viruses, 14(2), 345. https://doi.org/10.3390/v14020345