
Spatio-Temporal Epidemiology of the Spread of African Swine Fever in Wild Boar and the Role of Environmental Factors in South Korea

 ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
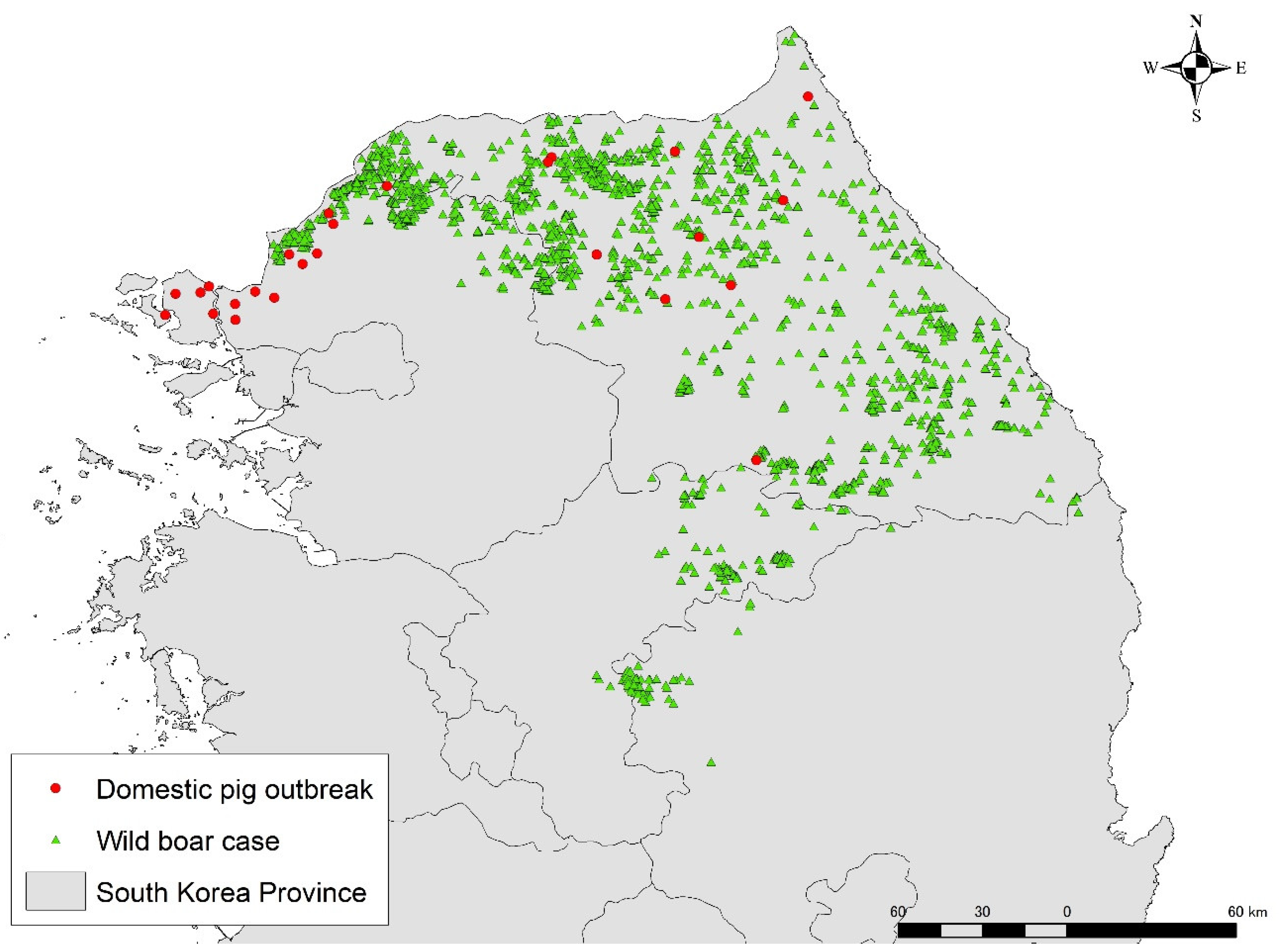
2.1. Epidemiological Data Preparation
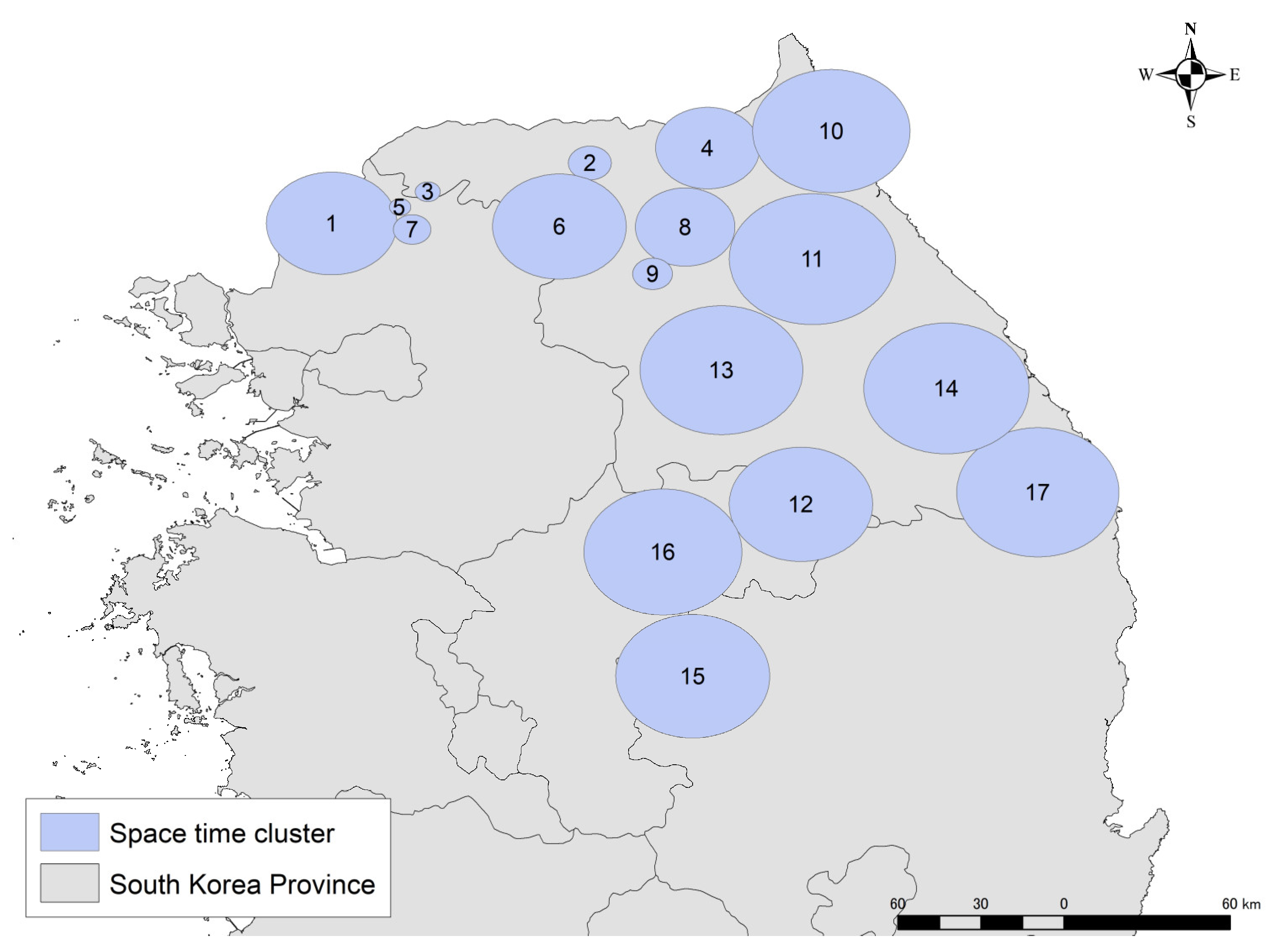
2.2. Identification of Spatio-Temporal Clusters
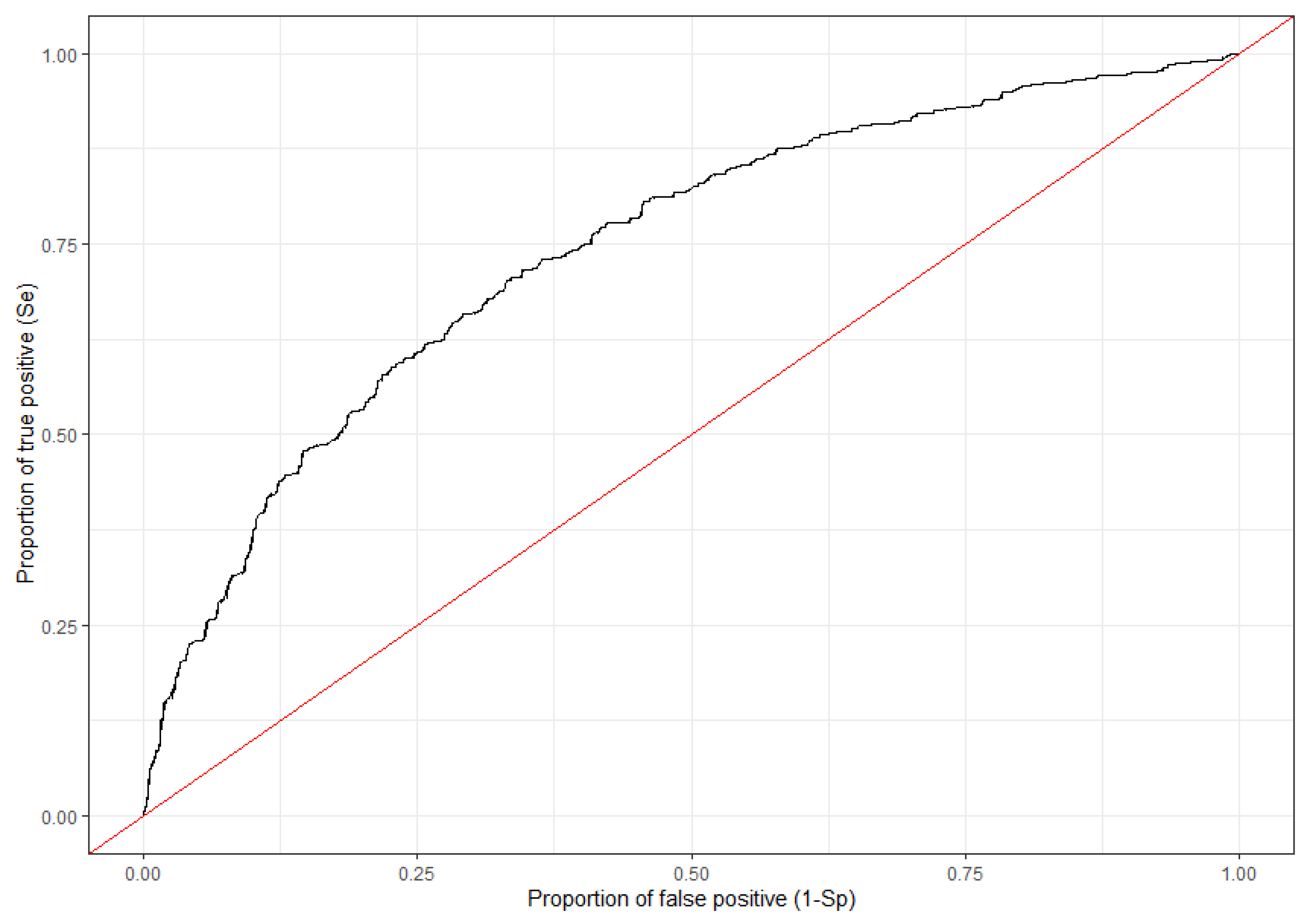
2.3. Identification of Environmental Factors Contributing to Spatio-Temporal Cluster Formation Based on GLLR Model
2.4. Estimation of Basic Reproduction Number (R0) in the Spatio-Temporal Cluster
3. Results
3.1. Identification of Spatio-Temporal Clusters
3.2. Identification of Environmental Factors Contributing to Spatio-Temporal Cluster Formation Based on a GLLR Model
3.3. Estimation of Basic Reproduction Number (R0) in the Spatio-Temporal Cluster
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sanchez-Vizcaino, J.M.; Mur, L.; Gomez-Villamandos, J.C.; Carrasco, L. An update on the epidemiology and pathology of African swine fever. J. Comp. Pathol. 2015, 152, 9–21. [Google Scholar] [CrossRef] [PubMed]
- De la Torre, A.; Bosch, J.; Iglesias, I.; Munoz, M.J.; Mur, L.; Martinez-Lopez, B.; Martinez, M.; Sanchez-Vizcaino, J.M. Assessing the risk of African swine fever introduction into the European Union by wild boar. Transbound. Emerg. Dis. 2015, 62, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organisation of the United Nations Statistics [FAO]. African Swine Fever: Detection and Diagnostic. A Manual for Veterinarians. 2017. Available online: http://www.fao.org/3/a-i7228e.pdf (accessed on 10 October 2022).
- Chenais, E.; Depner, K.; Guberti, V.; Dietze, K.; Viltrop, A.; Stahl, K. Epidemiological considerations on African swine fever in Europe 2014–2018. Porc. Health Manag. 2019, 5, 6. [Google Scholar] [CrossRef]
- Guberti, V.; Khomenko, S.; Masiulis, M.; Kerba, S. African Swine Fever in Wild Boar Ecology and Biosecurity; Rome. FAO Animal Production and Health Manual; FAO: Rome, Italy; OIE: Paris, France; EC: Ispra, Italy, 2019; No. 22. [Google Scholar]
- Adkin, A.; Coburn, H.; England, T.; Hall, S.; Hartnett, E.; Marooney, C.; Wooldridge, M.; Watson, E.; Cooper, J.; Cox, T. Risk Assessment for the Illegal Import of Contaminated Meat and Meat Products into Great Britain and the Subsequent Exposure of GB Livestock (IIRA): Foot and Mouth Disease (FMD), Classical Swine Fever (CSF), African Swine Fever (ASF), Swine Vesicular Disease (SVD); Veterinary Laboratories Agency: New Haw, UK, 2004. [Google Scholar]
- De Carvalho Ferreira, H.C.; Weesendorp, E.; Quak, S.; Stegeman, J.A.; Loeffen, W.L. Suitability of faeces and tissue samples as a basis for non-invasive sampling for African swine fever in wild boar. Vet. Microbiol. 2014, 172, 449–454. [Google Scholar] [CrossRef]
- World Organisation for Animal Health. African Swine Fever. 2022. Available online: https://www.woah.org/en/disease/african-swine-fever/ (accessed on 10 October 2022).
- Kedkovid, R.; Sirisereewan, C.; Thanawongnuwech, R. Major swine viral diseases: An Asian perspective after the African swine fever introduction. Porc. Health Manag. 2020, 6, 20. [Google Scholar] [CrossRef] [PubMed]
- OIE-WAHIS: World Animal Health Information System. Animal Disease Events. 2022. Available online: https://wahis.oie.int/#/events?viewAll=true (accessed on 1 February 2022).
- Ministry of Agriculture, Food and Rural Affairs, South Korea. Release of Information on Outbreaks of Livestock Infectious Diseases (ASF). 2022. Available online: https://mafra.go.kr/FMD-AI2/2241/subview.do (accessed on 27 January 2022).
- Ministry of Agriculture, Food and Rural Affairs, South Korea. Press Release: African Swine Fever. 2022. Available online: http://www.me.go.kr/search/totalSearch/search.jsp (accessed on 27 January 2022).
- Jo, Y.S.; Gortázar, C. African swine fever in wild boar: Assessing interventions in South Korea. Transbound. Emerg. Dis. 2021, 68, 2878–2889. [Google Scholar] [CrossRef] [PubMed]
- Segye. Detecting Dogs Are Deployed to Search for Dead Wild Boar Bodies. 2022. Available online: https://www.segye.com/newsView/20221016506928 (accessed on 17 November 2022).
- Ministry of Environment, South Korea. Detecting the Dead Body of a Wild Boar. 2022. Available online: https://me.go.kr/niwdc/web/board/read.do?menuId=24&boardId=1555430&boardMasterId=794&condition.hideCate=1 (accessed on 17 November 2022).
- PIGPEOPLE. Real-Time Status Board of African Swine Fever (ASF). 2022. Available online: http://www.pigpeople.net/mobile/article.html?no=7260 (accessed on 1 February 2022).
- Ministry of Environment, South Korea. The Habitat Density of Wild Boars Is Mainly Determined by the Capacity of the Habitat Environment. 2019. Available online: https://me.go.kr/home/web/board/read.do?boardMasterId=1&boardId=1038690&menuId=286 (accessed on 17 November 2022).
- Handonnews. ASF-Mediated, the Habitat Density of Wild Boars is Lowered. 2021. Available online: http://handonnews.kr/mobile/article.html?no=23592 (accessed on 17 November 2022).
- PIGPEOPLE. Domestic Wild Boar Expert: “It Is Impossible to Estimate the Number of Wild Boars in the Country”. 2019. Available online: http://www.pigpeople.net/news/article.html?no=7626 (accessed on 17 November 2022).
- Dodram Pig Farmers Cooperative. General Information. 2022. Available online: http://home.dodram.com/ (accessed on 5 May 2021).
- FAO. African Swine Fever in the Russian Federation: Risk Factors for Europe and Beyond Empres Watch 2013. Available online: https://www.fao.org/3/aq240e/aq240e.pdf (accessed on 21 December 2021).
- FAO. EMPRES-i Epidemiology (Frequency of African Swine Fever in Republic of Korea). Available online: https://empres-i.apps.fao.org/ (accessed on 5 May 2021).
- Iglesias, I.; Rodriguez, A.; Feliziani, F.; Rolesu, S.; de la Torre, A. Spatio-temporal Analysis of African Swine Fever in Sardinia (2012–2014): Trends in Domestic Pigs and Wild Boar. Transbound. Emerg. Dis. 2017, 64, 656–662. [Google Scholar] [CrossRef]
- ESRI. How Multi-Distance Spatial Cluster Analysis (Ripley’s K-function) Works. 2022. Available online: https://desktop.arcgis.com/en/arcmap/10.3/tools/spatial-statistics-toolbox/h-how-multi-distance-spatial-cluster-analysis-ripl.htm (accessed on 12 January 2011).
- Kulldorff, M. SaTScan; Information Management Services Inc.: Boston, MA, USA, 2010. [Google Scholar]
- Bosch, J.; Barasona, J.A.; Cadenas-Fernandez, E.; Jurado, C.; Pintore, A.; Denurra, D.; Cherchi, M.; Vicente, J.; Sanchez-Vizcaino, J.M. Retrospective spatial analysis for African swine fever in endemic areas to assess interactions between susceptible host populations. PLoS ONE 2020, 15, e0233473. [Google Scholar] [CrossRef]
- CGIAR-CSI. SRTM 90m DEM Digital Elevation Database. 2004. Available online: https://srtm.csi.cgiar.org/ (accessed on 15 May 2021).
- South Korea National Spatial Data Infrastructure Portal. Data Catalog. 2022. Available online: http://www.nsdi.go.kr/lxportal/?menuno=2679 (accessed on 17 May 2022).
- Esri. Near (Analysis). Available online: https://pro.arcgis.com/ja/pro-app/latest/tool-reference/analysis/near.htm (accessed on 29 July 2022).
- Tucker, C.J.; Pinzon, J.E.; Brown, M.E.; Slayback, D.A.; Pak, E.W.; Mahoney, R.; Vermote, E.F.; El Saleous, N. An extended AVHRR 8-km NDVI dataset compatible with MODIS and SPOT vegetation NDVI data. Int. J. Remote Sens. 2005, 26, 4485–4498. [Google Scholar] [CrossRef]
- Bosch, J.; Mardones, F.; Pérez, A.; De la Torre, A.; Muñoz, M.J. A maximum entropy model for predicting wild boar distribution in Spain. Span. J. Agric. Res. 2014, 12, 984–999. [Google Scholar] [CrossRef]
- Bosch, J.; Iglesias, I.; Munoz, M.J.; de la Torre, A. A Cartographic Tool for Managing African Swine Fever in Eurasia: Mapping Wild Boar Distribution Based on the Quality of Available Habitats. Transbound. Emerg. Dis. 2017, 64, 1720–1733. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.-T.; Pak, S.-I. Species distribution modeling for wild boar (Sus scropa) in the Republic of Korea using MODIS data. J. Prev. Vet. Med. 2020, 44, 89–95. [Google Scholar] [CrossRef]
- IFORCE. Global Accessibility Map. 2022. Available online: https://forobs.jrc.ec.europa.eu/products/gam/ (accessed on 12 January 2018).
- Unninayar, S.; Olsen, L.M. Monitoring, Observations, and Remote Sensing—Global Dimensions. In Reference Module in Earth Systems and Environmental Sciences; Elsevier: Amsterdam, The Netherlands, 2015. [Google Scholar]
- Willmott, C.J.; Kenji, M. Terrestrial Water Budget Data Archive: Monthly Time Series (1950–1999). 2001. Available online: https://sage.nelson.wisc.edu/data-and-models/atlas-of-the-biosphere/mapping-the-biosphere/ecosystems/soil-moisture/ (accessed on 29 September 2022).
- Bergmann, H.; Schulz, K.; Conraths, F.J.; Sauter-Louis, C. A review of environmental risk factors for African Swine Fever in European wild boar. Animals 2021, 11, 2692. [Google Scholar] [CrossRef] [PubMed]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. J. R. Meteorol. Soc. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Hidano, A. Introduction to Regression Analysis for Epidemiological Data (1). J. Vet. Epidemiol. 2020, 24, 29–35. [Google Scholar] [CrossRef]
- Hothorn, T.; Zeileis, A.; Farebrother, R.W.; Cummins, C.; Millo, G.; Mitchell, D.; Zeileis, M.A. Package ‘Lmtest’. Testing Linear Regression Models. Available online: https://cran.r-project.org/web/packages/lmtest/lmtest.pdf (accessed on 2 October 2022).
- Bates, D.M. Lme4: Mixed-Effects Modeling with R; Springer: New York, NY, USA, 2010. [Google Scholar]
- Fox, J.; Weisberg, S.; Adler, D.; Bates, D.; Baud-Bovy, G.; Ellison, S.; Firth, D.; Friendly, M.; Gorjanc, G.; Graves, S. Package ‘Car’; R Foundation for Statistical Computing: Vienna, Austria, 2012; Volume 16. [Google Scholar]
- Cessie, L.; Houwelingen, J. A Goodness-of-fit test for binary regression models, based on smoothing methods. Biometrics 1991, 47, 1267–1282. [Google Scholar] [CrossRef]
- Le Cessie, S.; van Houwelingen, J. Goodness of fit tests for generalized linear models based on random effect models. Biometrics 1995, 51, 600–614. [Google Scholar] [CrossRef]
- Jennie Pearce, S.F. Evaluating the predictive performance of habitat models developed using logistic regression. Ecol. Model. 2000, 133, 225–245. [Google Scholar] [CrossRef]
- Delamater, P.L.; Street, E.J.; Leslie, T.F.; Yang, Y.T.; Jacobsen, K.H. Complexity of the Basic Reproduction Number (R0). Emerg. Infect. Dis. 2019, 25, 1–4. [Google Scholar] [CrossRef]
- Anderson, R.M.; May, R.M. Infectious Diseases of Humans: Dynamics and Control; Oxford University Press: New York, NY, USA, 1992. [Google Scholar]
- Iglesias, I.; Perez, A.; Sánchez-Vizcaíno, J.; Muñoz, M.; Martínez, M.; De La Torre, A. Reproductive ratio for the local spread of highly pathogenic avian influenza in wild bird populations of Europe, 2005–2008. Epidemiol. Infect. 2011, 139, 99–104. [Google Scholar] [CrossRef]
- Iglesias, I.; Munoz, M.J.; Montes, F.; Perez, A.; Gogin, A.; Kolbasov, D.; de la Torre, A. Reproductive Ratio for the Local Spread of African Swine Fever in Wild Boars in the Russian Federation. Transbound. Emerg. Dis. 2016, 63, e237–e245. [Google Scholar] [CrossRef] [PubMed]
- Marcon, A.; Linden, A.; Satran, P.; Gervasi, V.; Licoppe, A.; Guberti, V. R0 estimation for the African swine fever epidemics in wild boar of Czech Republic and Belgium. Vet. Sci. 2019, 7, 2. [Google Scholar] [CrossRef]
- Lim, J.-S.; Kim, E.; Ryu, P.-D.; Pak, S.-I. Basic reproduction number of African swine fever in wild boars (Sus scrofa) and its spatiotemporal heterogeneity in South Korea. J. Vet. Sci. 2021, 22, e71. [Google Scholar] [CrossRef]
- Chenais, E.; Ståhl, K.; Guberti, V.; Depner, K. Identification of wild boar–habitat epidemiologic cycle in African swine fever epizootic. Emerg. Infect. Dis. 2018, 24, 810. [Google Scholar] [CrossRef] [PubMed]
- Pietschmann, J.; Guinat, C.; Beer, M.; Pronin, V.; Tauscher, K.; Petrov, A.; Keil, G.; Blome, S. Course and transmission characteristics of oral low-dose infection of domestic pigs and European wild boar with a Caucasian African swine fever virus isolate. Arch. Virol. 2015, 160, 1657–1667. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-J.; Cho, K.-H.; Ryu, J.-H.; Jang, M.-K.; Chae, H.-G.; Choi, J.-D.; Nah, J.-J.; Kim, Y.-J.; Kang, H.-E. Isolation and Genetic Characterization of African Swine Fever Virus from Domestic Pig Farms in South Korea, 2019. Viruses 2020, 12, 1237. [Google Scholar] [CrossRef] [PubMed]
- Depner, K.; Gortazar, C.; Guberti, V.; Masiulis, M.; More, S.; Oļševskis, E.; Thulke, H.H.; Viltrop, A.; Woźniakowski, G. Epidemiological analyses of African swine fever in the Baltic States and Poland: (Update September 2016–September 2017). EFSA J. 2017, 15, e05068. [Google Scholar]
- Fischer, M.; Hühr, J.; Blome, S.; Conraths, F.J.; Probst, C. Stability of African swine fever virus in carcasses of domestic pigs and wild boar experimentally infected with the ASFV “Estonia 2014” isolate. Viruses 2020, 12, 1118. [Google Scholar] [CrossRef]
- Probst, C.; Gethmann, J.; Amendt, J.; Lutz, L.; Teifke, J.P.; Conraths, F.J. Estimating the postmortem interval of wild boar carcasses. Vet. Sci. 2020, 7, 6. [Google Scholar] [CrossRef]
- Lim, S.J.; Han, S.H.; Park, J.Y.; Kim, N.H.; Namgung, H.; Oh, Y.; Park, Y.C. Wildlife as Potential Vectors of African Swine Fever Virus. J. For. Environ. Sci. 2022, 38, 55–63. [Google Scholar]
- Amendolia, S.; Lombardini, M.; Pierucci, P.; Meriggi, A. Seasonal spatial ecology of the wild boar in a peri-urban area. Mammal Res. 2019, 64, 387–396. [Google Scholar] [CrossRef]
- Ibaraki Prefecture. The Ecology of Wild Boars. 2020. Available online: https://www.pref.ibaraki.jp/nourinsuisan/hokunourin/kikaku/kikaku/inoshishinoseitai.html (accessed on 29 July 2022).
- Cukor, J.; Linda, R.; Václavek, P.; Šatrán, P.; Mahlerová, K.; Vacek, Z.; Kunca, T.; Havránek, F. Wild boar deathbed choice in relation to ASF: Are there any differences between positive and negative carcasses? Prev. Vet. Med. 2020, 177, 104943. [Google Scholar] [CrossRef] [PubMed]
- Cukor, J.; Linda, R.; Václavek, P.; Mahlerová, K.; Šatrán, P.; Havránek, F. Confirmed cannibalism in wild boar and its possible role in African swine fever transmission. Transbound. Emerg. Dis. 2020, 67, 1068–1073. [Google Scholar] [CrossRef] [PubMed]
- Pittiglio, C.; Khomenko, S.; Beltran-Alcrudo, D. Wild boar mapping using population-density statistics: From polygons to high resolution raster maps. PLoS ONE 2018, 13, e0193295. [Google Scholar] [CrossRef]
- Thurfjell, H.; Spong, G.; Ericsson, G. Effects of weather, season, and daylight on female wild boar movement. Acta Theriol. 2014, 59, 467–472. [Google Scholar] [CrossRef]
- Hankyung.com. Yeoncheon-gun Damaged 4.5 km of Wild Boar Fences Due to Heavy Rain. 2020. Available online: https://www.hankyung.com/politics/article/202008100999Y (accessed on 18 November 2022).
- National Integrated Drought Information System. Soil Moisture. 2022. Available online: https://www.drought.gov/topics/soil-moisture (accessed on 29 September 2022).
- Ministry of Agriculture, Food and Rural Affairs, South Korea. The Government Actively Promotes Measures to Prevent the Spread of African Swine Fever in Wild Boar 2021. Available online: https://www.mafra.go.kr/FMD-AI2/2241/subview.do (accessed on 14 February 2022).
- Sun, E.; Huang, L.; Zhang, X.; Zhang, J.; Shen, D.; Zhang, Z.; Wang, Z.; Huo, H.; Wang, W.; Huangfu, H.; et al. Genotype I African swine fever viruses emerged in domestic pigs in China and caused chronic infection. Emerg. Microbes Infect. 2021, 10, 2183–2193. [Google Scholar] [CrossRef]
- Sun, E.; Zhang, Z.; Wang, Z.; He, X.; Zhang, X.; Wang, L.; Wang, W.; Huang, L.; Xi, F.; Huangfu, H.; et al. Emergence and prevalence of naturally occurring lower virulent African swine fever viruses in domestic pigs in China in 2020. Sci. China Life Sci. 2021, 64, 752–765. [Google Scholar] [CrossRef]
- Ito, S.; Bosch, J.; Martínez-Avilés, M.; Sánchez-Vizcaíno, J.M. The Evolution of African Swine Fever in China: A Global Threat? Front. Vet. Sci. 2022, 9, 828498. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Cluster No. | Number of Cases | Start Date | End Date | Season | Case Doubling Time (Day) | Adjusted R-Square | Scenario 1 R0 (Dmin, max = 2, 9) | Confidence Interval (95%) | Scenario 2 R0 | Confidence Interval (95%) | Scenario 2 D (min–max) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 165 | 2020/1/9 | 2020/4/1 | Winter | 37.5 | 0.95 | 1.10 | 1.04–1.16 | 1.66 | 1.27–2.09 | 13–60 |
| 2 | 159 | 2020/1/16 | 2020/4/8 | Winter | 35.7 | 0.93 | 1.11 | 1.04–1.17 | 1.72 | 1.28–2.14 | 13–60 |
| 3 | 43 | 2020/3/12 | 2020/5/13 | Spring | 63.6 | 0.93 | 1.06 | 1.02–1.1 | 1.28 | 1.13–1.43 | 11–40.5 |
| 4 | 32 | 2020/8/13 | 2020/11/4 | Autumn | 117.5 | 0.96 | 1.03 | 1.01–1.05 | 1.15 | 1.07–1.23 | 11–40.5 |
| 5 | 18 | 2020/12/3 | 2020/12/16 | Winter | 12.1 | 0.83 | 1.33 | 1.13–1.51 | 3.06 | 1.81–4.38 | 13–60 |
| 6 | 135 | 2021/1/7 | 2021/3/17 | Winter | 32.1 | 0.93 | 1.12 | 1.05–1.19 | 1.82 | 1.3–2.27 | 13–60 |
| 7 | 66 | 2021/1/21 | 2021/4/14 | Winter | 55.0 | 0.96 | 1.07 | 1.03–1.11 | 1.45 | 1.18–1.74 | 13–60 |
| 8 | 42 | 2021/2/11 | 2021/5/5 | Spring | 73.0 | 0.96 | 1.05 | 1.02–1.08 | 1.25 | 1.11–1.38 | 11–40.5 |
| 9 | 9 | 2021/6/17 | 2021/7/21 | Summer | 105.0 | 0.97 | 1.04 | 1.01–1.06 | 1.10 | 1.06–1.14 | 9–21 |
| 10 | 11 | 2021/7/8 | 2021/8/25 | Summer | 138.6 | 0.95 | 1.03 | 1.01–1.04 | 1.07 | 1.05–1.1 | 9–21 |
| 11 | 62 | 2021/7/22 | 2021/9/15 | Summer | 36.3 | 0.92 | 1.11 | 1.04–1.17 | 1.29 | 1.18–1.39 | 9–21 |
| 12 | 99 | 2021/11/18 | 2022/1/5 | Winter | 29.1 | 0.65 | 1.13 | 1.05–1.21 | 1.84 | 1.34–2.4 | 13–60 |
| 13 | 50 | 2021/12/23 | 2022/2/16 | Winter | 51.0 | 0.93 | 1.07 | 1.03–1.12 | 1.49 | 1.2–1.8 | 13–60 |
| 14 | 189 | 2022/1/6 | 2022/3/30 | Winter | 36.1 | 0.94 | 1.11 | 1.04–1.17 | 1.70 | 1.28–2.13 | 13–60 |
| 15 | 95 | 2022/2/3 | 2022/4/27 | Spring | 56.4 | 0.95 | 1.07 | 1.03–1.11 | 1.31 | 1.14–1.49 | 11–40.5 |
| 16 | 35 | 2022/2/17 | 2022/3/9 | Winter | 19.7 | 0.80 | 1.20 | 1.08–1.31 | 2.30 | 1.5–3.07 | 13–60 |
| 17 | 5 | 2022/6/9 | 2022/7/6 | Summer | 231.0 | 0.70 | 1.02 | 1.01–1.03 | 1.05 | 1.03–1.06 | 9–21 |
| Variables | Grade (Threshold) | Total Samples (n = 2578) | Coefficients | Standard Errors | p-Value |
|---|---|---|---|---|---|
| Elevation | Grade I (232) | 648 | - | - | - |
| Grade II (352) | 649 | −0.185 | 0.079 | 0.099 | |
| Grade III (490) | 640 | 0.261 | 0.112 | 0.020 | |
| Grade IV (1157) | 641 | 0.082 | 0.112 | 0.459 | |
| Distance from road (roadway and sidewalk) | Grade I (276.2) | 645 | - | - | - |
| Grade II (602) | 644 | −0.009 | 0.111 | 0.933 | |
| Grade III (1171.5) | 644 | −0.072 | 0.112 | 0.521 | |
| Grade IV (6887.1) | 645 | −0.295 | 0.112 | 0.009 | |
| Wild boar distribution index | Grade I (0.415) | 516 | - | - | - |
| Grade II (0.504) | 517 | 0.216 | 0.125 | 0.086 | |
| Grade III (0.628) | 514 | −0.070 | 0.125 | 0.576 | |
| Grade IV (0.761) | 517 | −0.675 | 0.127 | >0.001 | |
| Grade V (1) | 514 | 0.794 | 0.128 | >0.001 | |
| Travel time to major cities | Grade I (57) | 647 | - | - | - |
| Grade II (107.5) | 642 | −0.048 | 0.112 | 0.670 | |
| Grade III (213) | 645 | −0.252 | 0.112 | 0.025 | |
| Grade IV (860) | 644 | 0.146 | 0.111 | 0.191 | |
| Soil moisture | Grade I (137.6) | 775 | - | - | - |
| Grade II (139.6) | 975 | −0.670 | 0.098 | >0.001 | |
| Grade III (141.3) | 258 | −0.280 | 0.144 | 0.053 | |
| Grade IV (143.2) | 570 | −0.965 | 0.114 | >0.001 | |
| Temperature seasonality (bioclim 4) | Grade I (9726.3) | 645 | - | - | - |
| Grade II (10,073) | 646 | −0.264 | 0.112 | 0.018 | |
| Grade III (10,226.8) | 642 | −0.221 | 0.112 | 0.048 | |
| Grade IV (10,435) | 645 | −0.311 | 0.112 | 0.005 | |
| Temperature annual range (bioclim 7) | Grade I (381) | 656 | - | - | - |
| Grade II (390) | 670 | −0.14 | 0.110 | 0.189 | |
| Grade III (396) | 650 | −0.27 | 0.111 | 0.015 | |
| Grade IV (411) | 602 | −0.36 | 0.114 | 0.002 | |
| Precipitation seasonality (bioclim 15) | Grade I (81) | 660 | - | - | - |
| Grade II (91) | 641 | 0.099 | 0.111 | 0.374 | |
| Grade III (98) | 656 | −0.062 | 0.111 | 0.573 | |
| Grade IV (105) | 621 | 0.039 | 0.112 | 0.728 | |
| Precipitation of warmest quarter (bioclim 18) | Grade I (731) | 658 | - | - | - |
| Grade II (792) | 634 | 0.190 | 0.112 | 0.089 | |
| Grade III (849) | 651 | 0.161 | 0.111 | 0.145 | |
| Grade IV (911) | 635 | −0.209 | 0.112 | 0.064 | |
| Precipitation of coldest quarter (bioclim 19) | Grade I (72) | 693 | - | - | - |
| Grade II (78) | 610 | −0.247 | 0.112 | 0.027 | |
| Grade III (94) | 640 | −0.060 | 0.110 | 0.583 | |
| Grade IV (163) | 635 | 0.093 | 0.110 | 0.397 |
| Variables | Variable | Total Samples (n = 2578) | Coefficients | Standard Errors | Odds Ratio (95% CI) | p-Value | VIF |
|---|---|---|---|---|---|---|---|
| Elevation | Grade I | 648 | - | - | - | - | 1.38 |
| Grade II | 649 | 0.530 | 0.164 | 1.70 (1.24–2.35) | 0.001 | ||
| Grade III | 640 | 1.142 | 0.195 | 3.13 (2.14–4.60) | >0.001 | ||
| Grade IV | 641 | 1.062 | 0.237 | 2.89 (1.82–4.61) | >0.001 | ||
| Distance from road (roadway and sidewalk) | Grade I | 645 | - | - | - | - | 1.04 |
| Grade II | 644 | −0.130 | 0.125 | 0.88 (0.69–1.12) | 0.300 | ||
| Grade III | 644 | −0.210 | 0.127 | 0.81 (0.63–1.04) | 0.099 | ||
| Grade IV | 645 | −0.519 | 0.132 | 0.60 (0.46–0.77) | >0.001 | ||
| Wild boar distribution index | Grade I | 516 | - | - | - | - | 1.08 |
| Grade II | 517 | 0.173 | 0.150 | 1.19 (0.89–1.60) | 0.248 | ||
| Grade III | 514 | −0.071 | 0.154 | 0.93 (0.69–1.26) | 0.646 | ||
| Grade IV | 517 | −0.378 | 0.190 | 0.69 (0.47–0.99) | 0.047 | ||
| Grade V | 514 | 0.643 | 0.261 | 1.90 (1.15–3.21) | 0.014 | ||
| Travel time to major cities | Grade I | 647 | - | - | - | - | 1.10 |
| Grade II | 642 | 0.137 | 0.132 | 1.15 (0.89–1.49) | 0.299 | ||
| Grade III | 645 | −0.004 | 0.144 | 1.00 (0.75–1.32) | 0.976 | ||
| Grade IV | 644 | 0.465 | 0.154 | 1.59 (1.18–2.15) | 0.002 | ||
| Soil moisture | Grade I | 775 | - | - | - | - | 1.31 |
| Grade II | 975 | −1.341 | 0.158 | 0.26 (0.19–0.36) | >0.001 | ||
| Grade III | 258 | −0.219 | 0.181 | 0.80 (0.56–1.15) | 0.226 | ||
| Grade IV | 570 | −1.005 | 0.155 | 0.37 (0.27–0.50) | >0.001 | ||
| Temperature annual range (bioclim 7) | Grade I | 656 | - | - | - | - | 1.36 |
| Grade II | 670 | −0.817 | 0.182 | 0.44 (0.31–0.63) | >0.001 | ||
| Grade III | 650 | −1.230 | 0.224 | 0.29 (0.19–0.45) | >0.001 | ||
| Grade IV | 602 | −1.068 | 0.227 | 0.34 (0.22–0.54) | >0.001 | ||
| Precipitation of warmest quarter (bioclim 18) | Grade I | 658 | - | - | - | - | 1.46 |
| Grade II | 634 | 0.561 | 0.162 | 1.75 (1.28–2.41) | >0.001 | ||
| Grade III | 651 | 0.420 | 0.207 | 1.52 (1.02–2.29) | 0.042 | ||
| Grade IV | 635 | −0.107 | 0.260 | 0.90 (0.54–1.50) | 0.679 | ||
| Precipitation of coldest quarter (bioclim 19) | Grade I | 693 | - | - | - | - | 1.78 |
| Grade II | 610 | −0.748 | 0.207 | 0.47 (0.31–0.71) | >0.001 | ||
| Grade III | 640 | −1.698 | 0.266 | 0.18 (0.11–0.31) | >0.001 | ||
| Grade IV | 635 | −1.834 | 0.317 | 0.16 (0.09–0.30) | >0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ito, S.; Bosch, J.; Jeong, H.; Aguilar-Vega, C.; Park, J.; Martínez-Avilés, M.; Sánchez-Vizcaíno, J.M. Spatio-Temporal Epidemiology of the Spread of African Swine Fever in Wild Boar and the Role of Environmental Factors in South Korea. Viruses 2022, 14, 2779. https://doi.org/10.3390/v14122779
Ito S, Bosch J, Jeong H, Aguilar-Vega C, Park J, Martínez-Avilés M, Sánchez-Vizcaíno JM. Spatio-Temporal Epidemiology of the Spread of African Swine Fever in Wild Boar and the Role of Environmental Factors in South Korea. Viruses. 2022; 14(12):2779. https://doi.org/10.3390/v14122779
Chicago/Turabian StyleIto, Satoshi, Jaime Bosch, Hyunkyu Jeong, Cecilia Aguilar-Vega, Jonghoon Park, Marta Martínez-Avilés, and Jose Manuel Sánchez-Vizcaíno. 2022. "Spatio-Temporal Epidemiology of the Spread of African Swine Fever in Wild Boar and the Role of Environmental Factors in South Korea" Viruses 14, no. 12: 2779. https://doi.org/10.3390/v14122779
APA StyleIto, S., Bosch, J., Jeong, H., Aguilar-Vega, C., Park, J., Martínez-Avilés, M., & Sánchez-Vizcaíno, J. M. (2022). Spatio-Temporal Epidemiology of the Spread of African Swine Fever in Wild Boar and the Role of Environmental Factors in South Korea. Viruses, 14(12), 2779. https://doi.org/10.3390/v14122779