

The Prevalence of Viral Pathogens among Bats in Kazakhstan
 , , , , ,
, , , , , {kind=link}
{kind=link}
{kind=link}
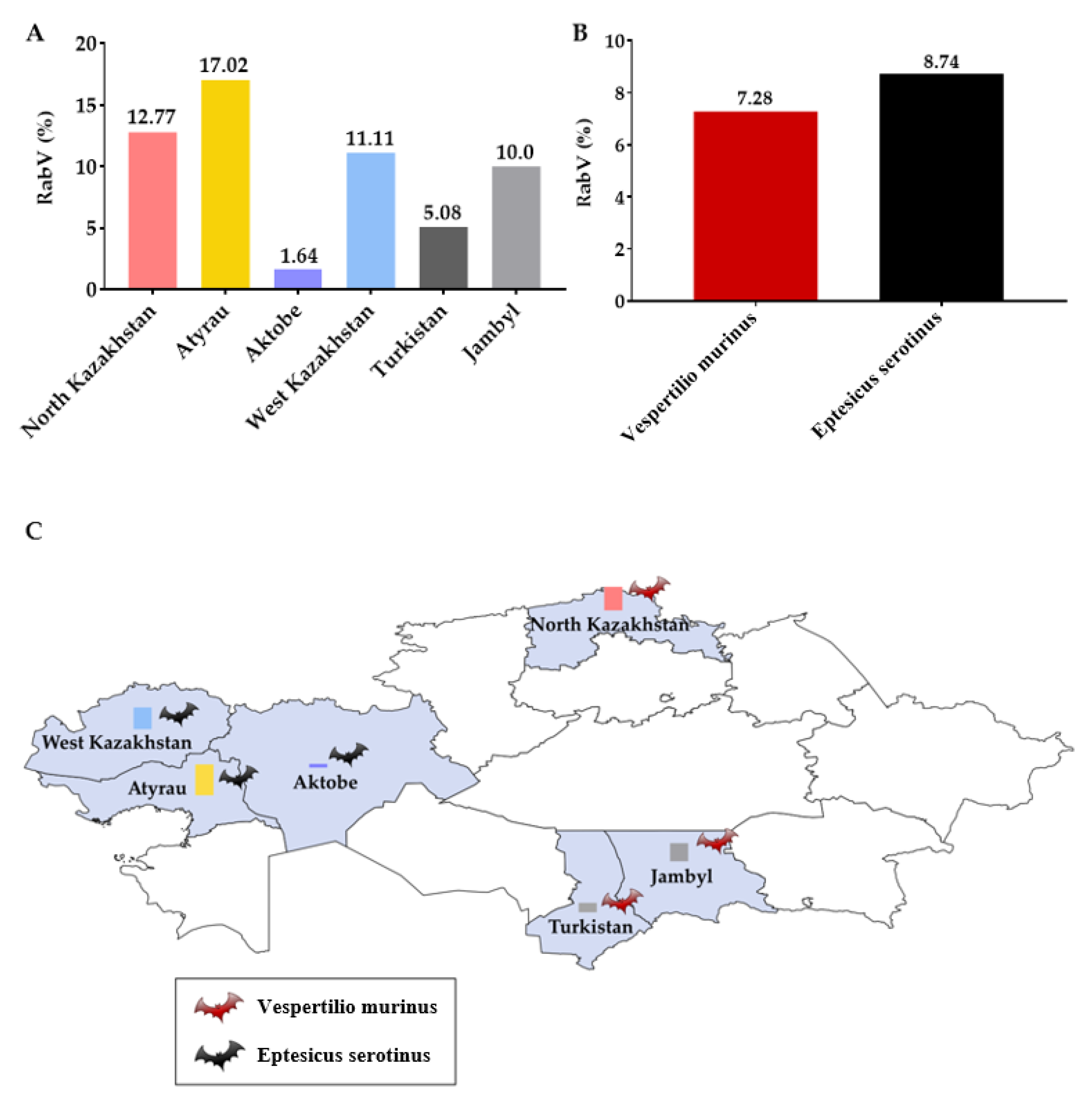{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Bat Sampling
2.2. RNA Extraction
2.3. PCR Study of the Samples from Bats
2.3.1. PCR for Influenza Virus Type A
2.3.2. RT-PCR for Detecting Coronaviruses
2.3.3. Real-Time RT-PCR for Detecting the Middle East Respiratory Syndrome Coronavirus (MERS-CoV)
2.3.4. Real-Time RT-PCR Confirmatory Assay (MERS-CoV)
2.3.5. Real-Time RT-PCR for Detecting the Crimean Congo Hemorrhagic Fever (CCHF) Virus and Coxiella Burnetii DNA
2.3.6. Real-Time RT-PCR for Detecting West the Nile fever (WNF) Virus and Rift Valley Fever (RVF) Virus
2.3.7. Real-Time RT-PCR for Detecting the Tick-Borne Encephalitis (TBE) Virus
2.3.8. Real-Time RT-PCR for Detecting Hantaviruses
2.3.9. Real-Time RT-PCR for Detecting the Rabies Virus
2.3.10. RT-PCR for the Rabies Virus Nucleoprotein Gene Product
2.4. Sequencing Assays, BLASTn Analysis and Phylogenetic Analyses
3. Results
3.1. Detection of RNA Viruses in Bats
3.2. Coronaviruses Sequencing and Phylogenetic Analysis
3.3. Rabies Virus Sequencing and Phylogenetic Analysis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Calisher, C.H.; Childs, J.E.; Field, H.E.; Holmes, K.V.; Schountz, T. Bats: Important Reservoir Hosts of Emerging Viruses. Clin. Microbiol. Rev. 2006, 19, 531–545. [Google Scholar] [CrossRef] [PubMed]
- Freuling, C. Novel Lyssavirus in Natterer’s Bat, Germany. Emerg. Infect. Dis. 2011, 17, 1519–1522. [Google Scholar] [CrossRef] [PubMed]
- Lau, S.K.; Woo, P.C.; Li, K.S.; Huang, Y.; Tsoi, H.W.; Wong, B.H.; Wong, S.S.; Leung, S.Y.; Chan, K.H.; Yuen, K.Y. Severe acute respiratory syndrome coronavirus-like virus in Chinese horseshoe bats. Proc. Natl. Acad. Sci. USA 2005, 102, 14040–14045. [Google Scholar] [CrossRef] [PubMed]
- Leroy, E.M.; Kumulungui, B.; Pourrut, X.; Rouquet, P.; Hassanin, A.; Yaba, P.; Délicat, A.; Paweska, J.T.; Gonzalez, J.P.; Swanepoel, R. Fruit bats as reservoirs of Ebola virus. Nature 2005, 438, 575–576. [Google Scholar] [CrossRef] [PubMed]
- Smith, I.; Broos, A.; De Jong, C.; Zeddeman, A.; Smith, C.; Smith, G.; Moore, F.; Barr, J.; Crameri, G.; Marsh, G.; et al. Identifying Hendra Virus Diversity in Pteropid Bats. PLoS ONE 2011, 6, e25275. [Google Scholar] [CrossRef] [PubMed]
- Swanepoel, R.; Smit, S.B.; Rollin, P.E.; Formenty, P.; Leman, P.A.; Kemp, A.; Burt, F.J.; Grobbelaar, A.A.; Croft, J.; Bausch, D.G.; et al. Studies of Reservoir Hosts for Marburg Virus. Emerg. Infect. Dis. 2007, 13, 1847–1851. [Google Scholar] [CrossRef] [PubMed]
- Letko, M.; Seifert, S.N.; Olival, K.J.; Plowright, R.K.; Munster, V.J. Bat-borne virus diversity, spillover and emergence. Nat. Rev. Genet. 2020, 18, 461–471. [Google Scholar] [CrossRef] [PubMed]
- Olival, K.J.; Hosseini, P.R.; Zambrana-Torrelio, C.; Ross, N.; Bogich, T.L.; Daszak, P. Erratum: Host and viral traits predict zoonotic spillover from mammals. Nature 2017, 548, 612. [Google Scholar] [CrossRef] [PubMed]
- Ben Hu, B.; Ge, X.; Wang, L.-F.; Shi, Z. Bat origin of human coronaviruses. Virol. J. 2015, 12, 221. [Google Scholar] [CrossRef]
- Bekenov, A.; Butovsky, P.M.; Kasabekov, B.B.; Lankin, P.M.; Strelkov, P.P.; Stogov, I.I.; Fedosenko, A.K.; Shaimardanov, R.T.; Shubin, G.I. Available online: https://www.acbk.kz/article/default/view?id=38 (accessed on 17 June 2021).
- Mendenhall, I.H.; Kerimbayev, A.A.; Strochkov, V.M.; Sultankulova, K.T.; Kopeyev, S.K.; Su, Y.C.; Smith, G.J.; Orynbayev, M.B. Discovery and Characterization of Novel Bat Coronavirus Lineages from Kazakhstan. Viruses 2019, 11, 356. [Google Scholar] [CrossRef] [PubMed]
- Humane Euthanasia of Bats for Public Health Rabies Testing of Michigan Rabies Working Group. Available online: https://www.michigan.gov/emergingdiseases/-/media/Project/Websites/emergingdiseases/Folder1/Humane_Euthanasia_of_Bats-Final.pdf?rev=ea62cbba8f884145b832d1c042d2e518&hash=9CE7B3697902E41AF346888F4FD9C645 (accessed on 1 September 2018).
- Spackman, E.; Senne, D.A.; Myers, T.J.; Bulaga, L.L.; Garber, L.P.; Perdue, M.L.; Lohman, K.; Daum, L.T.; Suarez, D.L. Development of a Real-Time Reverse Transcriptase PCR Assay for Type A Influenza Virus and the Avian H5 and H7 Hemagglutinin Subtypes. J. Clin. Microbiol. 2002, 40, 3256–3260. [Google Scholar] [CrossRef] [PubMed]
- Corman, V.M.; Eckerle, I.; Bleicker, T.; Zaki, A.; Landt, O.; Eschbach-Bludau, M.; van Boheemen, S.; Gopal, R.; Ballhause, M.; Bestebroer, T.M.; et al. Detection of a novel human coronavirus by real-time reverse-transcription polymerase chain reaction. Eurosurveillance 2012, 17, 20285. [Google Scholar] [CrossRef] [PubMed]
- Sultankulova, K.T.; Orynbaev, M.B.; Strochkov, V.M.; Shoraeva, K.A.; Chervyakova, O.V.; Sandybaev, N.T.; Sansyzbay, A.R. Phylogenetic analysis of rabies virus isolates isolated in Kazakhstan. Bull. PSU Chem. Biol. Ser. 2015, 2, 43–49. [Google Scholar]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.-S.; Noni, V.; Seelan, J.S.S.; Denel, A.; Khan, F.A.A. Ecological surveillance of bat coronaviruses in Sarawak, Malaysian Borneo. BMC Res. Notes 2021, 14, 461. [Google Scholar] [CrossRef] [PubMed]
- Anthony, S.J.; Johnson, C.K.; Greig, D.J.; Kramer, S.; Che, X.; Wells, H.; Hicks, A.L.; Joly, D.O.; Wolfe, N.D.; Daszak, P.; et al. Global patterns in coronavirus diversity. Virus Evol. 2017, 3, vex012. [Google Scholar] [CrossRef] [PubMed]
- Colombi, D.; Serra-Cobo, J.; Métras, R.; Apolloni, A.; Poletto, C.; López-Roig, M.; Bourhy, H.; Colizza, V. Mechanisms for lyssavirus persistence in non-synanthropic bats in Europe: Insights from a modeling study. Sci. Rep. 2019, 9, 537. [Google Scholar] [CrossRef] [PubMed]
- Sultanov, A.A.; Abdrakhmanov, S.K.; Abdybekova, A.M.; Karatayev, B.S.; Torgerson, P.R. Rabies in Kazakhstan. PLOS Neglected Trop. Dis. 2016, 10, e0004889. [Google Scholar] [CrossRef]
- Botvinkin, A.D. Fatal human cases of rabies in Eurasia after contacts with bats. 2011. [Google Scholar]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karagulov, A.I.; Argimbayeva, T.U.; Omarova, Z.D.; Tulendibayev, A.B.; Dushayeva, L.Z.; Svotina, M.A.; Melisbek, A.M.; Aubakir, N.A.; Barmak, S.M.; Sultankulova, K.T.; et al. The Prevalence of Viral Pathogens among Bats in Kazakhstan. Viruses 2022, 14, 2743. https://doi.org/10.3390/v14122743
Karagulov AI, Argimbayeva TU, Omarova ZD, Tulendibayev AB, Dushayeva LZ, Svotina MA, Melisbek AM, Aubakir NA, Barmak SM, Sultankulova KT, et al. The Prevalence of Viral Pathogens among Bats in Kazakhstan. Viruses. 2022; 14(12):2743. https://doi.org/10.3390/v14122743
Chicago/Turabian StyleKaragulov, Adilbay I., Takhmina U. Argimbayeva, Zamira D. Omarova, Ali B. Tulendibayev, Laura Zh. Dushayeva, Marina A. Svotina, Aibarys M. Melisbek, Nurdos A. Aubakir, Sabyrkhan M. Barmak, Kulyaisan T. Sultankulova, and et al. 2022. "The Prevalence of Viral Pathogens among Bats in Kazakhstan" Viruses 14, no. 12: 2743. https://doi.org/10.3390/v14122743
APA StyleKaragulov, A. I., Argimbayeva, T. U., Omarova, Z. D., Tulendibayev, A. B., Dushayeva, L. Z., Svotina, M. A., Melisbek, A. M., Aubakir, N. A., Barmak, S. M., Sultankulova, K. T., Alibekova, D. A., Yermekbai, T. T., Nametov, A. M., Lozovoy, D. A., Abeuov, K. B., & Orynbayev, M. B. (2022). The Prevalence of Viral Pathogens among Bats in Kazakhstan. Viruses, 14(12), 2743. https://doi.org/10.3390/v14122743