
SARS-CoV-2 and West Nile Virus Prevalence Studies in Raccoons and Raccoon Dogs from Germany

 , , , , , , and
, , , , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Molecular Detection of Viral RNA
2.2.1. SARS-CoV-2
2.2.2. WNV
2.3. Surrogate Virus Neutralization Test (sVNT) for SARS-CoV-2 Antibodies
2.4. Serological Investigation for Flaviviruses
3. Results
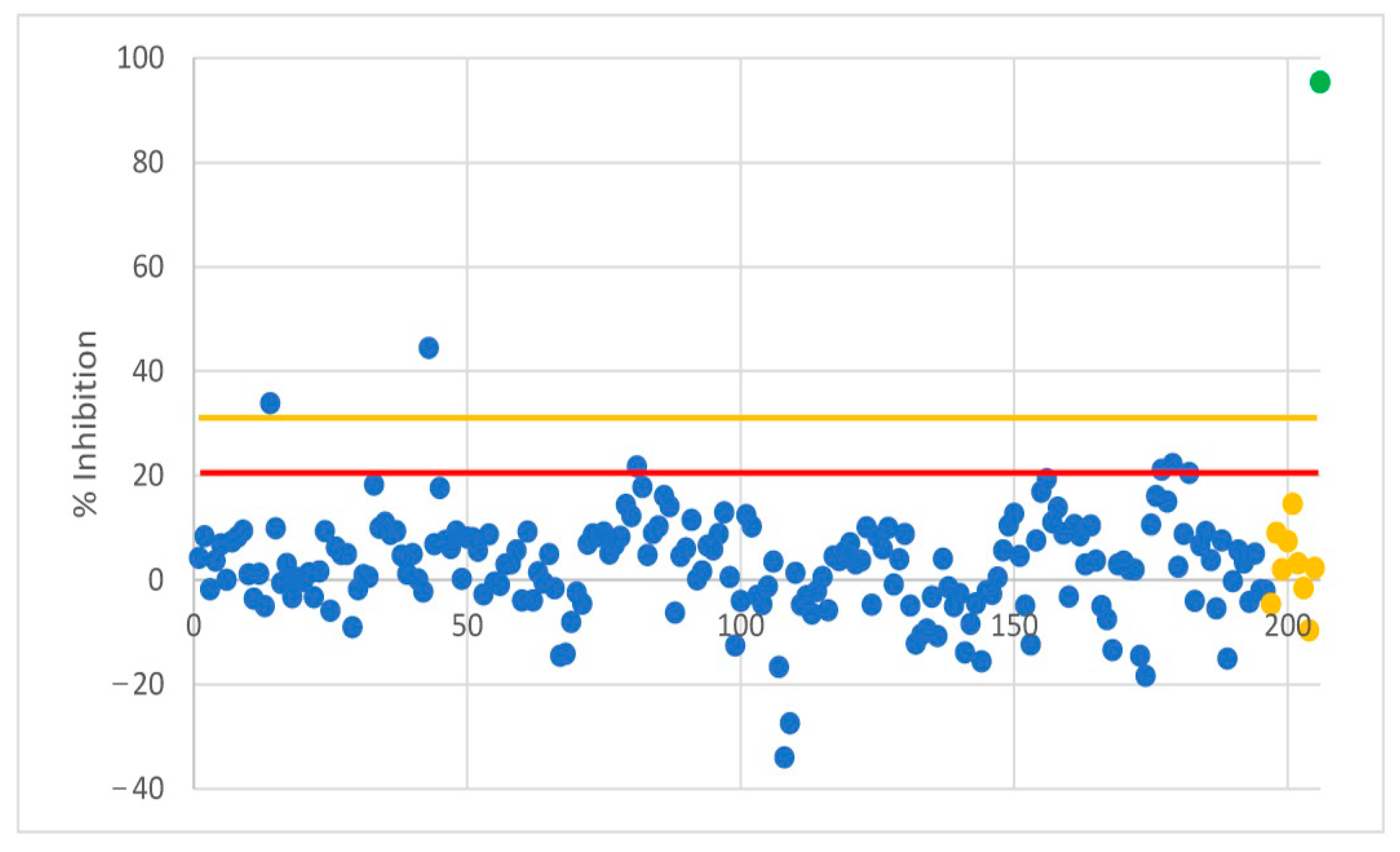
3.1. Investigation of Occurence of SARS-CoV-2 in Raccoons and Raccoon Dogs in Germany
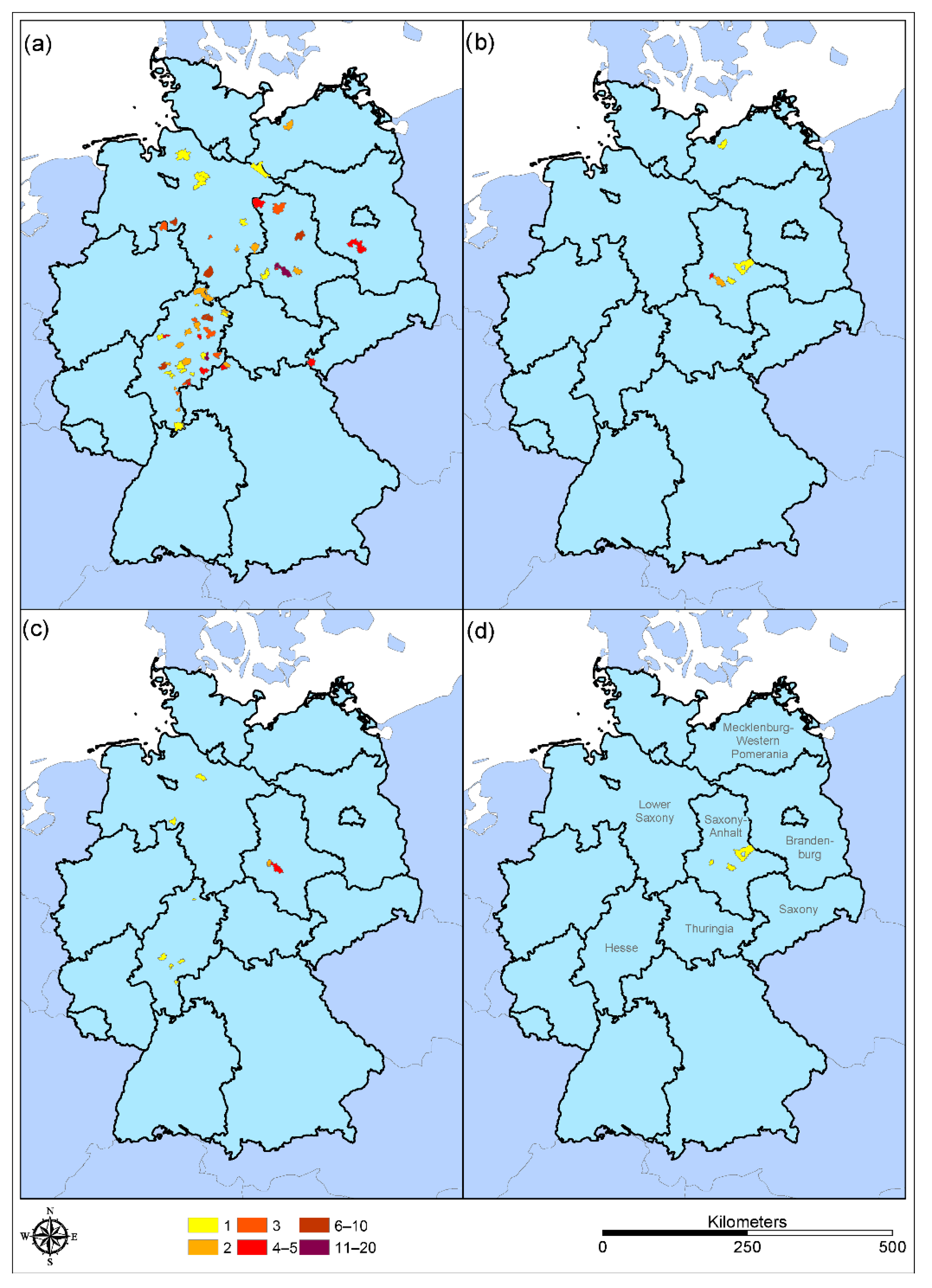
3.2. Incidence of WNV in the Domestic Raccoon Population
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lawson, B.; Neimanis, A.; Lavazza, A.; López-Olvera, J.R.; Tavernier, P.; Billinis, C.; Duff, J.P.; Mladenov, D.T.; Rijks, J.M.; Savić, S.; et al. How to Start Up a National Wildlife Health Surveillance Programme. Animals 2021, 11, 2543. [Google Scholar] [CrossRef]
- Schilling, A.-K.; Mazzamuto, M.V.; Romeo, C. A Review of Non-Invasive Sampling in Wildlife Disease and Health Research: What’s New? Animals 2022, 12, 1719. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, B.; García-Bocanegra, I.; Acevedo, P.; Cáceres, G.; Alves, P.C.; Gortázar, C. Stepping up from wildlife disease surveillance to integrated wildlife monitoring in Europe. Res. Vet. Sci. 2022, 144, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Kessler, S.; Harder, T.C.; Schwemmle, M.; Ciminski, K. Influenza A Viruses and Zoonotic Events-Are We Creating Our Own Reservoirs? Viruses 2021, 13, 2250. [Google Scholar] [CrossRef] [PubMed]
- Aznar, I.; Baldinelli, F.; Papanikolaou, A.; Stoicescu, A.; Van der Stede, Y. Annual Report on surveillance for avian influenza in poultry and wild birds in Member States of the European Union in 2020. EFSA J. 2021, 19, e06953. [Google Scholar] [PubMed]
- Verhagen, J.H.; Fouchier, R.A.M.; Lewis, N. Highly Pathogenic Avian Influenza Viruses at the Wild-Domestic Bird Interface in Europe: Future Directions for Research and Surveillance. Viruses 2021, 13, 212. [Google Scholar] [CrossRef]
- Frant, M.P.; Gal-Cisoń, A.; Bocian, Ł.; Ziętek-Barszcz, A.; Niemczuk, K.; Szczotka-Bochniarz, A. African Swine Fever (ASF) Trend Analysis in Wild Boar in Poland (2014–2020). Animals 2022, 12, 1170. [Google Scholar] [CrossRef] [PubMed]
- Sauter-Louis, C.; Conraths, F.J.; Probst, C.; Blohm, U.; Schulz, K.; Sehl, J.; Fischer, M.; Forth, J.H.; Zani, L.; Depner, K.; et al. African Swine Fever in Wild Boar in Europe-A Review. Viruses 2021, 13, 1717. [Google Scholar] [CrossRef]
- Blome, S.; Franzke, K.; Beer, M. African swine fever-A review of current knowledge. Virus Res. 2020, 287, 198099. [Google Scholar] [CrossRef]
- Garnier, R.; Graham, A.L. Insights from Parasite-Specific Serological Tools in Eco-Immunology. Am. Zool. 2014, 54, 363–376. [Google Scholar] [CrossRef] [PubMed]
- Nijman, V. Illegal and Legal Wildlife Trade Spreads Zoonotic Diseases. Trends Parasitol. 2021, 37, 359–360. [Google Scholar] [CrossRef] [PubMed]
- Bezerra-Santos, M.A.; Mendoza-Roldan, J.A.; Thompson, R.C.A.; Dantas-Torres, F.; Otranto, D. Legal versus Illegal Wildlife Trade: Zoonotic Disease Risks. Trends Parasitol. 2021, 37, 360–361. [Google Scholar] [CrossRef] [PubMed]
- Lafferty, K.D. The ecology of climate change and infectious diseases. Ecology 2009, 90, 888–900. [Google Scholar] [CrossRef] [PubMed]
- Schell, C.J.; Stanton, L.A.; Young, J.K.; Angeloni, L.M.; Lambert, J.E.; Breck, S.W.; Murray, M.H. The evolutionary consequences of human-wildlife conflict in cities. Evol. Appl. 2021, 14, 178–197. [Google Scholar] [CrossRef] [PubMed]
- König, H.J.; Kiffner, C.; Kramer-Schadt, S.; Fürst, C.; Keuling, O.; Ford, A.T. Human-wildlife coexistence in a changing world. Conserv. Biol. 2020, 34, 786–794. [Google Scholar] [CrossRef]
- Castillo-Contreras, R.; Mentaberre, G.; Fernandez Aguilar, X.; Conejero, C.; Colom-Cadena, A.; Ráez-Bravo, A.; González-Crespo, C.; Espunyes, J.; Lavín, S.; López-Olvera, J.R. Wild boar in the city: Phenotypic responses to urbanisation. Sci. Total Environ. 2021, 773, 145593. [Google Scholar] [CrossRef]
- Munnink, B.B.O.; Sikkema, R.S.; Nieuwenhuijse, D.F.; Molenaar, R.J.; Munger, E.; Molenkamp, R.; Van Der Spek, A.; Tolsma, P.; Rietveld, A.; Brouwer, M. Transmission of SARS-CoV-2 on mink farms between humans and mink and back to humans. Science 2021, 371, 172–177. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.; Mossman, K.; Baker, M.L. Zooanthroponotic potential of SARS-CoV-2 and implications of reintroduction into human populations. Cell Host Microbe 2021, 29, 160–164. [Google Scholar] [CrossRef] [PubMed]
- Kock, R.; Michel, A.L.; Yeboah-Manu, D.; Azhar, E.I.; Torrelles, J.B.; Cadmus, S.I.; Brunton, L.; Chakaya, J.M.; Marais, B.; Mboera, L.; et al. Zoonotic Tuberculosis-The Changing Landscape. Int. J. Infect. Dis. 2021, 113 (Suppl. S1), S68–S72. [Google Scholar] [CrossRef] [PubMed]
- Shivaprakash, K.N.; Sen, S.; Paul, S.; Kiesecker, J.M.; Bawa, K.S. Mammals, wildlife trade, and the next global pandemic. Curr. Biol. 2021, 31, 3671–3677.e3673. [Google Scholar] [CrossRef]
- Fagre, A.C.; Cohen, L.E.; Eskew, E.A.; Farrell, M.; Glennon, E.; Joseph, M.B.; Frank, H.K.; Ryan, S.J.; Carlson, C.J.; Albery, G.F. Assessing the risk of human-to-wildlife pathogen transmission for conservation and public health. Ecol. Lett. 2022, 25, 1534–1549. [Google Scholar] [CrossRef] [PubMed]
- Račnik, J.; Kočevar, A.; Slavec, B.; Korva, M.; Rus, K.R.; Zakotnik, S.; Zorec, T.M.; Poljak, M.; Matko, M.; Rojs, O.Z.; et al. Transmission of SARS-CoV-2 from Human to Domestic Ferret. Emerg. Infect. Dis. 2021, 27, 2450–2453. [Google Scholar] [CrossRef] [PubMed]
- Rentería-Solís, Z.M.; Hamedy, A.; Michler, F.-U.; Michler, B.A.; Lücker, E.; Stier, N.; Wibbelt, G.; Riehn, K. Alaria alata mesocercariae in raccoons (Procyon lotor) in Germany. Parasitol. Res. 2013, 112, 3595–3600. [Google Scholar] [CrossRef]
- Fischer, M.L.; Sullivan, M.J.P.; Greiser, G.; Guerrero-Casado, J.; Heddergott, M.; Hohmann, U.; Keuling, O.; Lang, J.; Martin, I.; Michler, F.-U.; et al. Assessing and predicting the spread of non-native raccoons in Germany using hunting bag data and dispersal weighted models. Biol. Invasions 2016, 18, 57–71. [Google Scholar] [CrossRef]
- Bharti, A.R.; Nally, J.E.; Ricaldi, J.N.; Matthias, M.A.; Diaz, M.M.; Lovett, M.A.; Levett, P.N.; Gilman, R.H.; Willig, M.R.; Gotuzzo, E. Leptospirosis: A zoonotic disease of global importance. Lancet Infect. Dis. 2003, 3, 757–771. [Google Scholar] [CrossRef]
- Beltrán-Beck, B.; García, F.J.; Gortázar, C. Raccoons in Europe: Disease hazards due to the establishment of an invasive species. Eur. J. Wildl. Res. 2012, 58, 5–15. [Google Scholar] [CrossRef]
- Michelitsch, A.; Wernike, K.; Ulrich, L.; Mettenleiter, T.C.; Beer, M. SARS-CoV-2 in animals: From potential hosts to animal models. Adv Virus Res 2021, 110, 59–102. [Google Scholar]
- Holmes, E.C.; Goldstein, S.A.; Rasmussen, A.L.; Robertson, D.L.; Crits-Christoph, A.; Wertheim, J.O.; Anthony, S.J.; Barclay, W.S.; Boni, M.F.; Doherty, P.C.; et al. The origins of SARS-CoV-2: A critical review. Cell 2021, 184, 4848–4856. [Google Scholar] [CrossRef] [PubMed]
- Rutherford, C.; Kafle, P.; Soos, C.; Epp, T.; Bradford, L.; Jenkins, E. Investigating SARS-CoV-2 Susceptibility in Animal Species: A Scoping Review. Environ. Health Insights 2022, 16, 11786302221107786. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Wen, Z.; Zhong, G.; Yang, H.; Wang, C.; Huang, B.; Liu, R.; He, X.; Shuai, L.; Sun, Z.; et al. Susceptibility of ferrets, cats, dogs, and other domesticated animals to SARS-coronavirus 2. Science 2020, 368, 1016–1020. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.-R.; Cao, Q.-D.; Hong, Z.-S.; Tan, Y.-Y.; Chen, S.-D.; Jin, H.-J.; Tan, K.-S.; Wang, D.-Y.; Yan, Y. The origin, transmission and clinical therapies on coronavirus disease 2019 (COVID-19) outbreak–an update on the status. Mil. Med. Res. 2020, 7, 11. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Weekly Epidemiological Update on COVID-19, 80th ed.; World Health Organization: New York, NJ, USA, 2022. [Google Scholar]
- Aguirre, A.A.; Catherina, R.; Frye, H.; Shelley, L. Illicit wildlife trade, wet markets, and COVID-19: Preventing future pandemics. World Med. Health Policy 2020, 12, 256–265. [Google Scholar] [CrossRef]
- Freuling, C.M.; Breithaupt, A.; Müller, T.; Sehl, J.; Balkema-Buschmann, A.; Rissmann, M.; Klein, A.; Wylezich, C.; Hoper, D.; Wernike, K.; et al. Susceptibility of raccoon dogs for experimental SARS-CoV-2 infection. Emerg. Infect. Dis. 2020, 26, 2982–2985. [Google Scholar] [CrossRef] [PubMed]
- Chueca, L.J.; Kochmann, J.; Schell, T.; Greve, C.; Janke, A.; Pfenninger, M.; Klimpel, S. De novo Genome Assembly of the Raccoon Dog (Nyctereutes procyonoides). Front. Genet. 2021, 12, 658256. [Google Scholar] [CrossRef]
- Francisco, R.; Hernandez, S.M.; Mead, D.G.; Adcock, K.G.; Burke, S.C.; Nemeth, N.M.; Yabsley, M.J. Experimental susceptibility of North American raccoons (Procyon lotor) and striped skunks (Mephitis mephitis) to SARS-CoV-2. Front. Vet. Sci. 2021, 8, 715307. [Google Scholar] [CrossRef] [PubMed]
- Corman, V.M.; Landt, O.; Kaiser, M.; Molenkamp, R.; Meijer, A.; Chu, D.K.W.; Bleicker, T.; Brunink, S.; Schneider, J.; Schmidt, M.L.; et al. Detection of 2019 novel coronavirus (2019-nCoV) by real-time RT-PCR. Euro. Surveill. 2020, 25, 23–30. [Google Scholar] [CrossRef]
- Santos, P.D.; Michel, F.; Wylezich, C.; Hoper, D.; Keller, M.; Holicki, C.M.; Szentiks, C.A.; Eiden, M.; Muluneh, A.; Neubauer-Juric, A.; et al. Co-infections: Simultaneous detections of West Nile virus and Usutu virus in birds from Germany. Transbound. Emerg. Dis. 2021, 69, 776–792. [Google Scholar] [CrossRef]
- Ziegler, U.; Santos, P.D.; Groschup, M.H.; Hattendorf, C.; Eiden, M.; Hoper, D.; Eisermann, P.; Keller, M.; Michel, F.; Klopfleisch, R.; et al. West Nile Virus Epidemic in Germany Triggered by Epizootic Emergence, 2019. Viruses 2020, 12, 448. [Google Scholar] [CrossRef]
- Bakonyi, T.; Haussig, J.M. West Nile virus keeps on moving up in Europe. Eurosurveillance 2020, 25, 2001938. [Google Scholar] [CrossRef]
- Klaus, C.; Horugel, U.; Hoffmann, B.; Beer, M. Tick-borne encephalitis virus (TBEV) infection in horses: Clinical and laboratory findings and epidemiological investigations. Vet. Microbiol. 2013, 163, 368–372. [Google Scholar] [CrossRef]
- Pulkkinen, L.I.A.; Butcher, S.J.; Anastasina, M. Tick-Borne Encephalitis Virus: A Structural View. Viruses 2018, 10, 350. [Google Scholar] [CrossRef] [PubMed]
- Lindquist, L.; Vapalahti, O. Tick-borne encephalitis. Lancet 2008, 371, 1861–1871. [Google Scholar] [CrossRef]
- Sinnecker, H. Zeckenencephalitis in Deutschland. Zbl. Bact. Orig. 1960, 180, 12–18. [Google Scholar]
- Ziegler, U.; Lühken, R.; Markus, K.; Cadar, D.; van der Grinten, E.; Michel, F.; Albrecht, K.; Eiden, M.; Rinder, M.; Lachmann, L.; et al. West Nile virus epizootic in Germany, 2018. Elsevier 2018, 162, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Michel, F.; Sieg, M.; Fischer, D.; Keller, M.; Eiden, M.; Reuschel, M.; Schmidt, V.; Schwehn, R.; Rinder, M.; Urbaniak, S.; et al. Evidence for West Nile Virus and Usutu Virus Infections in Wild and Resident Birds in Germany, 2017 and 2018. Viruses 2019, 11, 674. [Google Scholar] [CrossRef] [PubMed]
- Jost, H.; Bialonski, A.; Maus, D.; Sambri, V.; Eiden, M.; Groschup, M.H.; Gunther, S.; Becker, N.; Schmidt-Chanasit, J. Isolation of usutu virus in Germany. Am. J. Trop. Med. Hyg. 2011, 85, 551–553. [Google Scholar] [CrossRef] [PubMed]
- Becker, N.; Jost, H.; Ziegler, U.; Eiden, M.; Hoper, D.; Emmerich, P.; Fichet-Calvet, E.; Ehichioya, D.U.; Czajka, C.; Gabriel, M.; et al. Epizootic emergence of Usutu virus in wild and captive birds in Germany. PLoS ONE 2012, 7, e32604. [Google Scholar] [CrossRef]
- Ziegler, U.; Jost, H.; Muller, K.; Fischer, D.; Rinder, M.; Tietze, D.T.; Danner, K.J.; Becker, N.; Skuballa, J.; Hamann, H.P.; et al. Epidemic Spread of Usutu Virus in Southwest Germany in 2011 to 2013 and Monitoring of Wild Birds for Usutu and West Nile Viruses. Vector-Borne Zoonotic Dis. 2015, 15, 481–488. [Google Scholar] [CrossRef]
- Cadar, D.; Bosch, S.; Jost, H.; Borstler, J.; Garigliany, M.M.; Becker, N.; Schmidt-Chanasit, J. Putative Lineage of Novel African Usutu Virus, Central Europe. Emerg. Infect. Dis. 2015, 21, 1647–1650. [Google Scholar] [CrossRef]
- Ashraf, U.; Ye, J.; Ruan, X.; Wan, S.; Zhu, B.; Cao, S. Usutu virus: An emerging flavivirus in Europe. Viruses 2015, 7, 219–238. [Google Scholar] [CrossRef]
- Heinz, F.X.; Stiasny, K. Flaviviruses and their antigenic structure. J. Clin. Virol. 2012, 55, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Beck, C.; Lowenski, S.; Durand, B.; Bahuon, C.; Zientara, S.; Lecollinet, S. Improved reliability of serological tools for the diagnosis of West Nile fever in horses within Europe. PLoS Negl. Trop. Dis. 2017, 11, e0005936. [Google Scholar] [CrossRef] [PubMed]
- Eiden, M.; Vina-Rodriguez, A.; Hoffmann, B.; Ziegler, U.; Groschup, M.H. Two new real-time quantitative reverse transcription polymerase chain reaction assays with unique target sites for the specific and sensitive detection of lineages 1 and 2 West Nile virus strains. J. Vet. Diagn. Investig. 2010, 22, 748–753. [Google Scholar] [CrossRef]
- Seidowski, D.; Ziegler, U.; von Ronn, J.A.; Muller, K.; Huppop, K.; Muller, T.; Freuling, C.; Muhle, R.U.; Nowotny, N.; Ulrich, R.G.; et al. West Nile virus monitoring of migratory and resident birds in Germany. Vector-Borne Zoonotic Dis. 2010, 10, 639–647. [Google Scholar] [CrossRef] [PubMed]
- Mayr, A.; Bachmann, P.A.; Bibrack, B.; Wittmann, G. Neutralisationstest. In Virologische Arbeitsmethoden, Band II (Serologie); Mayr, A., Bachmann, P.A., Bibrack, B., Wittmann, G., Eds.; Gustav Fischer Verlag: Jena, Germany, 1977; pp. 457–534. [Google Scholar]
- Yeh, J.Y.; Lee, J.H.; Park, J.Y.; Seo, H.J.; Moon, J.S.; Cho, I.S.; Kim, H.P.; Yang, Y.J.; Ahn, K.M.; Kyung, S.G.; et al. A diagnostic algorithm to serologically differentiate West Nile virus from Japanese encephalitis virus infections and its validation in field surveillance of poultry and horses. Vector-Borne Zoonotic Dis. 2012, 12, 372–379. [Google Scholar] [CrossRef] [PubMed]
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R.; et al. A Novel Coronavirus from Patients with Pneumonia in China, 2019. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef]
- Chan, J.F.-W.; Yuan, S.; Kok, K.-H.; To, K.K.-W.; Chu, H.; Yang, J.; Xing, F.; Liu, J.; Yip, C.C.-Y.; Poon, R.W.-S. A familial cluster of pneumonia associated with the 2019 novel coronavirus indicating person-to-person transmission: A study of a family cluster. Lancet 2020, 395, 514–523. [Google Scholar] [CrossRef]
- Boehm, E.; Kronig, I.; Neher, R.A.; Eckerle, I.; Vetter, P.; Kaiser, L.; Geneva Center for Emerging Viral Diseases. Novel SARS-CoV-2 variants: The pandemics within the pandemic. Clin. Microbiol. Infect. 2021, 27, 1109–1117. [Google Scholar] [CrossRef]
- World Health Organization, Coronavirus Disease (COVID-19) Weekly Epidemiological Update and Weekly Operational Update. Available online: https://www.who.int/emergencies/diseases/novel-coronavirus-2019/situation-reports (accessed on 9 August 2022).
- Guan, Y.; Zheng, B.J.; He, Y.Q.; Liu, X.L.; Zhuang, Z.X.; Cheung, C.L.; Luo, S.W.; Li, P.H.; Zhang, L.J.; Guan, Y.J.; et al. Isolation and Characterization of Viruses Related to the SARS Coronavirus from Animals in Southern China. Science 2003, 302, 276–278. [Google Scholar] [CrossRef]
- Ziegler, U.; Angenvoort, J.; Klaus, C.; Nagel-Kohl, U.; Sauerwald, C.; Thalheim, S.; Horner, S.; Braun, B.; Kenklies, S.; Tyczka, J.; et al. Use of competition ELISA for monitoring of West Nile virus infections in horses in Germany. Int. J. Environ. Res. Public Health 2013, 10, 3112–3120. [Google Scholar] [CrossRef]
- Ziegler, U.; Bergmann, F.; Fischer, D.; Müller, K.; Holicki, C.M.; Sadeghi, B.; Sieg, M.; Keller, M.; Schwehn, R.; Reuschel, M.; et al. Spread of West Nile Virus and Usutu Virus in the German Bird Population, 2019–2020. Microorganisms 2022, 10, 807. [Google Scholar] [CrossRef]
- Root, J.J.; Bentler, K.T.; Nemeth, N.M.; Gidlewski, T.; Spraker, T.R.; Franklin, A.B. Experimental infection of raccoons (Procyon lotor) with West Nile virus. Am. J. Trop. Med. Hyg. 2010, 83, 803–807. [Google Scholar] [CrossRef] [PubMed]
- Hagag, I.T.; Langner, T.; Groschup, M.H.; Keller, M. Molecular surveillance revealed no SARS-CoV-2 spillovers to raccoons (Procyon lotor) in four German federal states. Eur. J. Wildl. Res. 2022, 68, 54. [Google Scholar] [CrossRef] [PubMed]
- Ohno, Y.; Sato, H.; Suzuki, K.; Yokoyama, M.; Uni, S.; Shibasaki, T.; Sashika, M.; Inokuma, H.; Kai, K.; Maeda, K. Detection of antibodies against Japanese encephalitis virus in raccoons, raccoon dogs and wild boars in Japan. J. Vet. Med. Sci. 2009, 71, 1035–1039. [Google Scholar] [CrossRef]
- Blitvich, B.J.; Juarez, L.I.; Tucker, B.J.; Rowley, W.A.; Platt, K.B. Antibodies to West Nile virus in raccoons and other wild peridomestic mammals in Iowa. J. Wildl. Dis. 2009, 45, 1163–1168. [Google Scholar] [CrossRef]
- Bentler, K.T.; Hall, J.S.; Root, J.J.; Klenk, K.; Schmit, B.; Blackwell, B.F.; Ramey, P.C.; Clark, L. Serologic evidence of West Nile virus exposure in North American mesopredators. Am. J. Trop. Med. Hyg. 2007, 76, 173–179. [Google Scholar] [CrossRef]
- Ganzenberg, S.; Sieg, M.; Ziegler, U.; Pfeffer, M.; Vahlenkamp, T.W.; Horugel, U.; Groschup, M.H.; Lohmann, K.L. Seroprevalence and Risk Factors for Equine West Nile Virus Infections in Eastern Germany, 2020. Viruses 2022, 14, 1191. [Google Scholar] [CrossRef] [PubMed]
- Friedrich-Loeffler-Institut; Federal Research Institute for Animal Health. TSIS-TierSeuchenInformationsSystem. Available online: https://tsis.fli.de/Reports/Info_SO.aspx?ts=416&guid=ff9aa17d-b468-4fad-926a-ce9a1745a1d0 (accessed on 9 November 2022).
- Friedrich-Loeffler-Institut; Federal Research Institute for Animal Health. West-Nil-Virus–Aktueller Stand: Bisher 38 Bestätigte Infektionen bei Pferden und Vögeln in Deutschland. Available online: https://www.fli.de/de/aktuelles/kurznachrichten/neues-einzelansicht/aktueller-stand-bisher-38-bestaetigte-west-nil-virus-infektionen-bei-pferden-und-voegeln-in-deutschland/ (accessed on 9 November 2022).
- OIE, World Organisation for Animal Health. Chapter 8.20.-West Nile Fever. In Terrestrial Animal Health Code; Eloit, M., Ed.; OIE, World Organisation for Animal Health: Paris, France, 2022; Volume 30. [Google Scholar]



{kind=link}
{kind=link}
| German Federal State | Raccoon (n = 229) | Raccoon Dog (n = 11) | ||
|---|---|---|---|---|
| n | % | n | % | |
| Baden-Wurttemberg | 4 | 1.75 | ||
| Bavaria | 10 | 4.37 | - | - |
| Brandenburg | 4 | 1.75 | - | - |
| Hesse | 104 | 45.41 | - | - |
| Mecklenburg-Western Pomerania | 14 | 6.11 | 2 | 18.18 |
| Lower Saxony | 30 | 13.10 | - | - |
| North-Rhine Westphalia | 3 | 1.31 | - | - |
| Saxony | 6 | 2.62 | - | - |
| Saxony-Anhalt | 54 | 23.58 | 9 | 81.82 |
| Sample Number | Postal Code | Location | Date | Sex | Age | Land Cover | Positive for |
|---|---|---|---|---|---|---|---|
| Raccoon-B-023 | 63500 | Seligenstadt (Hesse) | 23 February 2021 | male | adult | forest | n.d. |
| Raccoon-B-033 | 31600 | Uchta (Lower Saxony) | 5 March 2021 | female | adult | farmland | USUV |
| Raccoon-B-034 | 34131 | Kassel (Hesse) | 28 February 2021 | male | adult | urban | n.d. |
| Raccoon-B-120 | 35510 | Butzbach (Hesse) | 17 September 2021 | female | adult | urban | n.d. |
| Raccoon-B-125 | 06458 | Hausneindorf (Saxony-Anhalt) | 3 September 2021 | female | juvenil | forest | n.d. |
| Raccoon-B-142 | 06466 | Gatersleben (Saxony-Anhalt) | 8 October 2021 | female | adult | urban | WNV |
| Raccoon-B-153 | 61194 | Assenheim (Hesse) | 7 October 2021 | male | adult | urban | USUV |
| Raccoon-B-155 | 06466 | Gatersleben (Saxony-Anhalt) | 14 October 2021 | male | juvenil | urban | WNV |
| Raccoon-B-171 | 06449 | Neu-Königsaue (Saxony-Anhalt) | 18 November 2021 | female | juvenil | forest | WNV |
| Raccoon-B-174 | 06449 | Neu-Königsaue (Saxony-Anhalt) | 6 November 2021 | male | adult | forest | WNV |
| Raccoon-B-178 | 06449 | Neu-Königsaue (Saxony-Anhalt) | 9 January 2022 | male | adult | forest | USUV |
| Raccoon-B-179 | 06458 | Hausneindorf (Saxony-Anhalt) | 9 January 2022 | female | adult | forest | n.d. |
| Raccoon-B-180 | 27389 | Stemmen (Lower Saxony) | 14 January 2022 | male | adult | farmland | WNV |
| Raccoon-B-185 | 63683 | Schwickartshausen (Hesse) | 30 January 2022 | male | adult | forest | n.d. |
| Raccoon-B-190 | 06449 | Neu-Königsaue (Saxony-Anhalt) | 16 February 2022 | male | adult | forest | WNV |
| Raccoon-B-201 | 06449 | Neu-Königsaue (Saxony-Anhalt) | 19 March 2022 | female | adult | forest | WNV |
| Raccoon Dog-B-002 | 39264 | Gommern (Saxony-Anhalt) | 20 February 2021 | male | adult | forest | n.d |
| Raccoon Dog-B-004 | 06406 | Bernburg (Saxony-Anhalt) | 27 February 2021 | female | adult | forest | TBEV |
| Raccoon Dog-B-006 | 06458 | Hausneindorf (Saxony-Anhalt) | 19 March 2022 | male | juvenil | farmland | USUV |
| WNV | USUV | TBEV | |
|---|---|---|---|
| Raccoons | 7/195 (3.6%) | 3/195 (1.5%) | 0/195 |
| Raccoon dogs | 0/9 | 1/9 (11.1%) | 1/9 (11.1%) |
| Blocking ELISA | Differentiation by Virus Neutralization Tests | ||||
|---|---|---|---|---|---|
| FLI ID-Number | Inhibition Percentage (IP%) | Result | WNV Antibody Titer (ND50) | USUV Antibody Titer (ND50) | TBEV Antibody Titer (ND50) |
| 1-W-B-142 | 93.02 | POS | 1920 | 160 | <10 |
| 1-W-B-155 | 94.64 | POS | 2560 | 60 | <10 |
| 1-W-B-171 | 91.56 | POS | 120 | <10 | 15 |
| 1-W-B-174 | 94.33 | POS | 640 | 30 | <10 |
| 1-W-B-180 | 94.22 | POS | 2560 | 480 | <10 |
| 1-W-B-190 | 93.15 | POS | 40 | 10 | <10 |
| 1-W-B-201 | 94.71 | POS | 960 | 20 | <10 |
| 1-M-B-006 | 86.44 | POS | <10 | 80 | <10 |
| 1-W-B-033 | 85.71 | POS | <10 | 40 | <10 |
| 1-W-B-153 | 34.87 | DOUBT | <10 | 80 | <10 |
| 1-W-B-178 | 44.12 | POS | 10 | 80 | <10 |
| 1-M-B-004 | 89.74 | POS | 30 | 15 | 120 |
| Blocking ELISA | Differentiation by Virus Neutralization Tests | ||||
|---|---|---|---|---|---|
| FLI ID-Number | Inhibition Percentage (IP%) | Result | WNV Antibody Titer (ND50) | USUV Antibody Titer (ND50) | TBEV Antibody Titer (ND50) |
| 1-W-B-125 | 47.27 | POS | 480 | 80 | 160 |
| 1-M-B-002 | 43.17 | POS | <10 | 30 | 10 |
| 1-W-B-023 | 32.61 | DOUBT | <10 | <10 | <10 |
| 1-W-B-034 | 32.04 | DOUBT | <10 | 20 | 10 |
| 1-W-B-120 | 33.55 | DOUBT | 30 | 60 | <10 |
| 1-W-B-179 | 88.90 | POS | 80 | 30 | <10 |
| 1-W-B-185 | 40.24 | POS | 40 | 30 | <10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Keller, M.; Peter, N.; Holicki, C.M.; Schantz, A.V.; Ziegler, U.; Eiden, M.; Dörge, D.D.; Vilcinskas, A.; Groschup, M.H.; Klimpel, S. SARS-CoV-2 and West Nile Virus Prevalence Studies in Raccoons and Raccoon Dogs from Germany. Viruses 2022, 14, 2559. https://doi.org/10.3390/v14112559
Keller M, Peter N, Holicki CM, Schantz AV, Ziegler U, Eiden M, Dörge DD, Vilcinskas A, Groschup MH, Klimpel S. SARS-CoV-2 and West Nile Virus Prevalence Studies in Raccoons and Raccoon Dogs from Germany. Viruses. 2022; 14(11):2559. https://doi.org/10.3390/v14112559
Chicago/Turabian StyleKeller, Markus, Norbert Peter, Cora M. Holicki, Anna V. Schantz, Ute Ziegler, Martin Eiden, Dorian D. Dörge, Andreas Vilcinskas, Martin H. Groschup, and Sven Klimpel. 2022. "SARS-CoV-2 and West Nile Virus Prevalence Studies in Raccoons and Raccoon Dogs from Germany" Viruses 14, no. 11: 2559. https://doi.org/10.3390/v14112559
APA StyleKeller, M., Peter, N., Holicki, C. M., Schantz, A. V., Ziegler, U., Eiden, M., Dörge, D. D., Vilcinskas, A., Groschup, M. H., & Klimpel, S. (2022). SARS-CoV-2 and West Nile Virus Prevalence Studies in Raccoons and Raccoon Dogs from Germany. Viruses, 14(11), 2559. https://doi.org/10.3390/v14112559