Abstract
Short open reading frames (sORFs) are a newly identified family of genes, and the functions of most sORF genes and their encoded peptides (SEPs) are still unknown. In this study, two ATP synthase subunits were identified in kuruma shrimp (Marsupenaeus japonicus) as SEPs, namely MjATP5I and MjATP5L. They were widely distributed in all of the tested tissues of shrimp and upregulated in hemocytes and intestines in response to WSSV challenge. The injection of recombinant proteins (rMjATP5I and rMjATP5L) increased the expression of Ie1 and Vp28, while the knockdown of MjATP5I and MjATP5L decreased the expression of Ie1 and Vp28. All of the results suggest that MjATP5I and MjATP5L were beneficial for WSSV replication. Further exploration found that MjATP5I and MjATP5L RNAi significantly improved the shrimp survival rates, reduced ATP production, and upregulated the expression of antimicrobial peptide genes post viral challenge, and the two ATPase subunits and Relish negatively regulated each other. These results reveal that MjATP5I and MjATP5L facilitated WSSV duplication by regulating the production of ATP contents and the expression of antimicrobial peptide genes in shrimp.
1. Introduction
Kuruma shrimp Marsupenaeus japonicus, as one of the important economic animals in aquaculture, are often threatened by diseases caused by viruses, bacteria, fungi, and parasites. It is reported that more than half of shrimp diseases are caused by viruses [1]. In particular, the viral disease in shrimp culture caused by white spot syndrome virus (WSSV) is quite serious and results in significant losses in shrimp aquaculture. However, there are few prophylactic and therapeutic measures to control viral diseases. Understanding the mechanism of shrimp responding to viral infections is essential for controlling these diseases.
Shrimp, as invertebrates, lack adaptive immunity and depend entirely on their innate immunity, which includes humoral immunity and cellular immunity. In humoral immunity, the expression of antimicrobial peptide (AMP) genes is regulated by the Toll pathway, the immune deficiency (IMD) pathway, and the Janus kinase/signal transducer and activator of transcription pathway [2]. Some studies have shown that the expression of AMP genes such as the anti-lipopolysaccharide factors Alf-b1, Alf-c1, Alf-c2, Alf-d2, and Alf-e1 is regulated by transcription factor Relish in the IMD signal pathway [2,3,4]. In shrimp, the AMPs (mainly Alfs) are proved to suppress the WSSV replication and show strong antiviral roles in shrimp immunity [5,6].
In recent years, a new family of small open reading frame genes and their encoded peptides (SEPs) was identified in animals, plants, fungi, and bacteria based on expanded data from next-generation sequencing technologies. They usually encode proteins of less than 100 amino acids in length and are called small open reading frames or short open reading frames (sORFs) [7,8,9]. The sORF family is essentially a hidden genome in the genomes of organisms due to the low molecular mass and the difficulties in screening and identification [7]. It includes thousands of genes, while most of their functions are still unknown [10,11]. So, it is possible to find novel proteins with interesting functions from the sORF family. With the development of related studies, more and more sORFs have been identified to function crucially in some biological processes, such as development, cancer, and innate immunity [8,12,13].
ATP synthase, one of the most important proteins in mitochondria, is the only protein to synthesize ATP in the electron transport chain [14]. In mammals, the ATP synthase is the F1F0-ATPase, which is located in the inner membrane of mitochondria and forms a hetero-oligomeric complex with a molecular mass of around 650 kDa [15]. The F1F0-ATP synthase complex fulfils the vital functions in ATP synthesis and produces most of the ATP for cells [16], and is composed of two major parts, the F0-ATPase portion and the F1-ATPase portion. F0-ATPase is fat-soluble, and contains a, b, c, d, e, F6, A6L, f, and g subunits [17]. Meanwhile, F1-ATPase is water-soluble, and consists of α, β, γ, δ, and ε subunits [18]. In addition, F1F0-ATP synthase also structurally determines the cristae morphology of mitochondria [16]. Deviations in the structure of ATP synthase and its subunits might lead to various diseases in animals and humans [19,20,21]. The beta subunit of the F1-ATP synthase in shrimp, detected only in the granular hemocytes, could bind to WSSV in the immune defense reaction [22,23].
The ATP5I gene encodes ATP synthase subunit e in humans, which is one subunit of the mitochondrial F0-ATPase complex. ATP5I contains an ATP synthase subunit e domain. Its main function is to interact with the ATP synthase subunit g to promote the ATP synthase forming dimers [24]. It is reported that the antisense strand of ATP5I can cause the inhibition of cell growth in human hepatocellular carcinoma [25]. In shrimp Penaeus vannamei, the ATP synthase subunit e was screened and identified as a growth-associated marker gene [26]. ATP5L gene encodes the ATP synthase subunit g in humans, which is located in the F0 portion of the ATPase complex. ATP5L contains an ATP synthase subunit g domain. In addition to its functions in the interaction with ATP synthase subunit e [24], the ATP5L encoded mRNA functions as a direct target of miR-570 to facilitate the ATP loss in human platelet storage [27]. It is reported that the ATP synthase subunit g from yeast mitochondrial F1F0-ATP synthase is required for the oxidase activity of cytochrome c [28]. In Pacific white shrimp (P. vannamei), ATP synthase subunit g was identified and characterized as a candidate gene related to ammonia tolerance [29]. However, the genes encoding for ATP synthase subunits have not been characterized in the innate immunity of invertebrates.
There have been no reports about the ATPase subunits in the innate immunity of shrimp. Therefore, the present study aimed to detect the involvement of ATPase subunits in kuruma shrimp M. japonicus. We identified two genes, called MjATP5I and MjATP5L, in kuruma shrimp, which encoded for ATP synthase subunit e and g, respectively. They both belong to the sORF family, with predicted protein lengths of less than 100 amino acids. The tissue distribution and time course expression profiles of MjATP5I and MjATP5L were detected at the transcriptional level via quantitative real-time polymerase chain reaction (qPCR) analysis. Recombinant protein injection and the knockdown assay were used to check the potential functions of MjATP5I and MjATP5L in vivo. Additionally, the possible mechanisms of action for MjATP5I and MjATP5L in the shrimp viral immune reaction were investigated.
2. Materials and Methods
2.1. Bioinformatics Analysis
The nucleotide sequences of MjATP5I and MjATP5L were obtained from transcriptome sequencing of kuruma shrimp M. japonicus (BGI, Shenzhen, China). The gene sequences of MjATP5I and MjATP5L were analyzed with BLASTx program in the National Center for Biotechnology Information server (https://blast.ncbi.nlm.nih.gov). The DNA sequences were translated into the amino acid sequences using the translate tool of ExPASy (https://web.expasy.org/translate/), while the isoelectric point and molecular weight were predicted by the compute pI/Mw tool in ExPASy (https://web.expasy.org/compute_pi/). The sequences were aligned by the ClustalW program to construct the neighbor-joining phylogenetic trees by MEGA 5.2 software [30]. Additionally, the functional domains of MjATP5I and MjATP5L were detected by the simple module architecture research tool (SMART, http://smart.embl-heidelberg.de/).
2.2. Tissue Distribution and Expression Profile Analysis
Healthy kuruma shrimp M. japonicus (around 10 g per shrimp) were obtained from a local shrimp farm in Qingdao, Shandong Province, China, and cultured in the recirculating aquaculture system filled with aerated natural seawater at 22 °C. Shrimp were cultured for more than two days until the sample collection and the immune challenge.
Tissues including the hemocytes, heart, hepatopancreas, gills, stomach, and intestines of at least three normal shrimp were collected for the total RNAs extraction with RNAiso Plus (Takara, Dalian, China). Then, they were reversely transcribed into the first strand cDNAs by PrimeScript™ II 1st Strand cDNA Synthesis Kit (Takara, Dalian, China), diluted 20-fold and used as the templates for the distribution analysis by qPCR. The primers MjATP5I-RTF with MjATP5I-RTR and MjATP5L-RTF with MjATP5L-RTR (Table 1) for qPCR analysis are listed in Table 1. β-actin with the primers actin-RTF and actin-RTR (Table 1) was used as the internal reference gene. The procedure of qPCR was as follows: 94 °C for 10 min; 40 cycles of 94 °C for 15 s, 60 °C for 60 s, and 76 °C for 2 s; and a melting curve analysis from 65 °C to 95 °C. The tissue distributions of MjATP5I and MjATP5L were computed using the 2−ΔΔCT method. GraphPad Prism 8.0 (GraphPad Software, San Diego, CA, USA) was used to analyze the data and construct the figures.

Table 1.
Primers used in this study.
For the immune infection, 50 μL (1 × 105 copies) of WSSV was injected into shrimp as the challenged group, while the same volume of sterile PBS (137 mM NaCl, 10 mM Na2HPO4, 2.7 mM KCl, 1.8 mM KH2PO4, pH 7.4) was injected into the other shrimp as the control group. Total RNAs were obtained from hemocytes and intestines at 0, 6, 12, 24, 36, and 48 h after the immune challenge. Then, qPCR was carried out to check the expression patterns of MjATP5I and MjATP5L following the above procedure.
2.3. Recombinant Protein Expression and Purification
The full-length open reading frames (ORFs) of MjATP5I and MjATP5L were amplified with their corresponding expression primer ExF and ExR listed in Table 1. After purification and double restriction enzyme digestion, the DNA fragment for MjATP5I was ligated into the EcoR I and Xho I sites of pET32a(+) vector (Novagen, Madison, WI, USA), while the fragment for MjATP5L was ligated into the EcoR I and Not I sites of pET32a(+) vector using T4 DNA ligase (Thermo Fisher Scientific). Then, after cloning and sequencing to verify the sequence accuracy, the recombinant plasmids MjATP5I-pET32a and MjATP5L-pET32a were transformed into Escherichia coli Rosetta competent cells. Additionally, 0.5 mM isopropyl-1-thio-b-D-galactopyranosid (IPTG) was added to induce the recombinant protein expression at 37 °C for 5 h. The recombinant proteins (rMjATP5I and rMjATP5L) were all expressed in the inclusion bodies, which were washed with Buffer A (50 mM Tris-HCl, 5 mM EDTA, pH 8.0) and Buffer B (2 M Urea, 50 mM Tris-HCl, 5 mM EDTA, pH 8.0) twice, and then diluted in Buffer C (8 M Urea, 100 mM Tris-HCl, 10 mM DTT, pH 8.0) at 37 °C for 1 h. After centrifugation at 12,000× g for 10 min, the supernatants of the solutes were dialyzed against an at least 50-times greater volume of buffer (100 mM Tris-HCl, 5 mM EDTA, 5 mM cysteine, pH = 8.0) at 4 °C for over 12 h twice. Finally, the supernatants of the dialysates were purified by Ni-nitrilotriacetic acid (Ni-NTA) resin (GE Healthcare, Piscataway, NJ, USA) by means of affinity chromatography following the instructions. The purified proteins were dialyzed against 100 mM Tris-HCl (pH 8.0) and stored at −20 °C for use.
Thioredoxin (Trx) was recombinantly expressed and purified as the control tag protein by transforming the empty pET32a (+) vector into E. coli Rosetta competent cells and inducing with IPTG. As expressed in the supernatant, Trx was purified directly with Ni-NTA resin after the ultrasonication of bacteria. The purified Trx protein was dialyzed against 100 mM Tris-HCl (pH 8.0) and cryopreserved at −20 °C for use.
2.4. Protein Injection Assay
For the protein injection in vivo, the purified recombinant protein was injected into the shrimp body cavity in vivo. First of all, the recombinant protein (rMjATP5I or rMjATP5L) was mixed with WSSV by gently shaking at 4 °C for 1 h. Additionally, the mixture containing 10 μg of recombinant protein with 1 × 105 copies of WSSV was injected into shrimp. Meanwhile, the mixture of Trx (10 μg) with WSSV (1 × 105 copies) was injected into other shrimp simultaneously as the control group.
Additionally, 1 h later, hemocytes were collected for the immunocytochemistry assay to check whether the recombinant protein entered into hemocytes. Briefly, the hemolymph was collected by a 5-mL syringe preloading with the mixture of anticoagulant (450 mM NaCl, 10 mM KCl, 10 mM EDTA, 10 mM HEPES, pH 7.5) and 4% paraformaldehyde in equal proportion. The hemolymph was deposited onto the coated glass slides and sat for at least two hours at room temperature. Then, hemocytes were washed with PBS six times, and treated with 0.2% TritonX-100 at 37 °C for 5 min to enhance the permeability of the hemocyte membrane. After blocking with 3% BSA at 37 °C for half an hour, hemocytes were incubated with mouse anti-His antibody (1:1000 in 3% BSA; ZSGB-BIO, Beijing, China) at 4 °C for at least 12 h, and then hatched with FITC labeled goat anti-mouse IgG(H+L) (1:1000 in 3% BSA; ZSGB-BIO) at 37 °C for 3 h. The nuclei of hemocytes were dyed with 4′,6-diamidino-2-phenylindole (DAPI, 1:1000 diluted in PBS) for 10 min at room temperature. Finally, Olympus BX51 fluorescence microscope (Olympus Corporation, Tokyo, Japan) was used to observe hemocytes and take pictures.
At 48 h after WSSV challenge, total RNAs were extracted from shrimp hemocytes and intestines to reversely transcribe the first-strand cDNAs, which were diluted 20-fold and used as the templates for qPCR to analyze the viral duplication. The expression of WSSV Ie1 gene and Vp28 gene was detected by qPCR. The data were analyzed by the 2−ΔΔCT method and processed with GraphPad Prism 8.0 software.
2.5. RNA Interference
The primers MjATP5I-RNAiF with MjATP5I-RNAiR and MjATP5L-RNAiF with MjATP5L-RNAiR (Table 1) were used to amplify the DNA fragments for the synthesis of double-stranded RNAs (dsRNAs). T7 RNA polymerase (Thermo Fisher Scientific, Waltham, MA, USA) was used to synthesize the dsRNAs for MjATP5I and MjATP5L. After being purified by chloroform, dsRNAs were diluted to 1 μg/μL with DEPC-treated water. The primer Gfp-RNAiF and Gfp-RNAiR (Table 1) was used for the amplification of the green fluorescent protein (Gfp) nucleotide fragment, which was used to synthesize the Gfp dsRNA as the control in the RNA interference (RNAi) assay. Each shrimp was injected with 50 μg of dsRNA. Additionally, 24 h later, another 50 μg of dsRNA was injected into the shrimp. At 24 h after the second dsRNA injection, hemocytes and intestines were isolated for the total RNA extraction. The RNAi effects were detected by qPCR using the primers MjATP5I-RTF1 with MjATP5I-RTR1 and MjATP5L-RTF1 with MjATP5L-RTR1 (Table 1). Then, WSSV (1 × 105 copies) was injected into shrimp. Total RNAs were obtained from shrimp hemocytes and intestines at 48 h post WSSV infection. Additionally, the relative expression of WSSV immediate early gene (Ie1) and envelope protein 28 gene (Vp28) was analyzed by qPCR using the primers listed in Table 1.
The expression of Relish and some antimicrobial peptide genes, such as Alf-b1, Alf-c1, Alf-c2, Alf-d2, and Alf-e1 after RNAi of MjATP5I and MjATP5L, was detected by qPCR using the primers listed in Table 1.
2.6. Survival Rate Assay
For the survival rate assay, the shrimp were divided into three groups—the dsGfp-injected shrimp, the dsMjATP5I-injected shrimp, and the dsMjATP5L-injected shrimp. Each group contained at least 30 shrimp. At 24 h after the second dsRNA injection, WSSV (1 × 105 copies) was injected into the shrimp. Then, the number of dead shrimp was recorded every 24 h. The survival rates of each group were statistically analyzed, and the survival curves were constructed using GraphPad Prism 8.0.
2.7. ATP Content Detection Assay
After knockdown of MjATP5I and MjATP5L followed by the WSSV challenge, hemocytes were collected from at least three shrimp as mentioned above. Then, the ATP content in shrimp hemocytes was detected using the ATP content detection kit (BC0300, Solarbio, Beijing, China) following the manufacturers’ instruction. In brief, hemocytes were lysed with the extraction solution, sonicated for 1 min (ice bath, power 20% or 200 W, sonication 2 s and stop 1 s), and centrifuged at the speed of 10,000× g for 10 min at 4 °C to separate the pellet and the supernatant. The harvested supernatant was extracted with chloroform to retain the supernatant, which was mixed with reagent I as well as the pre-prepared working solution. Additionally, the absorbance value was measured at 10 s and 3 min 10 s at A340 using a spectrophotometer (M200pro, Tecan, Männedorf, Switzerland). The absorbance value of the ATP standard was also determined at the same time. The ATP content in the extracted hemocytes was calculated following the manufacturers’ instructions. The ATP content of the hemocytes from the Gfp RNAi shrimp were used as the control. Graphpad Prism 8.0 software was used to analyze results and construct figures.
2.8. The Expression Analysis of MjATP5I and MjATP5L after Relish Knockdown
For Relish RNAi in shrimp, the primers Relish-RNAiF and Relish-RNAiR (Table 1) were used to amplify DNA fragment for Relish dsRNA, which was synthesized and purified following the above method. After the dsRNA injection, the hemocytes were collected for total RNA extraction and cDNA synthesis. QPCR analysis was performed to check the knockdown effect in hemocytes with primers Relish-RTF and Relish-RTR (Table 1).
At 24 h after the second dsRNA injection, WSSV (1 × 105 copies) was injected into the shrimp body cavity. Total RNAs were obtained from hemocytes at 48 h post WSSV infection. Then, the relative expression of MjATP5I and MjATP5L was analyzed by qPCR using primers listed in Table 1. The Gfp RNAi shrimp were used as the control.
3. Results
3.1. Sequence Analysis of MjATP5I and MjATP5L
The ORF of MjATP5I contained 258 base pairs (bps) of nucleotides, encoding a putative protein of 85 amino acids (Figure 1A). The predicted molecular mass of MjATP5I was 9.46 kDa with a predicted isoelectric point of 9.57. MjATP5I contained an ATP synthase subunit e (ATP-synt E) function domain (Figure 1B). From the phylogenetic neighbor-joining (NJ) tree (Figure 1C), we could find that ATP5Is could be divided two groups—one group contained some arthropods, including insects and crustaceans, and the other group contained Eurytemora affinis, Caerostris darwini, and Parasteatoda tepidariorum. MjATP5I belonged to the crustacean subgroup. A high similarity (96.5%) could be found between MjATP5I and ATP5I from P. vannamei (Figure 1D).
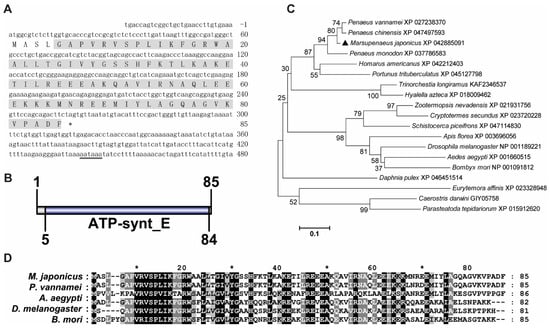
Figure 1.
Sequence analysis of MjATP5I. (A) The nucleotide and deduced amino acid sequence of MjATP5I. The letters with a grey background represent the ATP synthase subunit e domain. The double-underlined nucleotides show the polyadenylation signal region. (B) The domain schematics of MjATP5I. MjATP5I contained an ATP synthase E domain with 80 amino acids. (C) The phylogenetic tree analysis of ATP5Is from different animal species, including Aedes aegypti, Amphibalanus amphitrite, Apis florea, Bombyx mori, Caerostris darwini, Cryptotermes secundus, Daphnia pulex, Drosophila melanogaster, Eurytemora affinis, Homarus americanus, Hyalella azteca, Marsupenaeus japonicus, Parasteatoda tepidariorum, Penaeus chinensis, Penaeus monodon, Penaeus vannamei, Pollicipes pollicipes, Portunus trituberculatus, Schistocerca piceifrons, Trinorchestia longiramus, and Zootermopsis nevadensis. MjATP5I is marked by a solid black triangle. All of the amino acid sequences were obtained from GenBank. (D) Multiple sequence alignment of ATP5Is from M. japonicus, P. vannamei, A. aegypti, D. melanogaster, and B. mori. The conserved sites are labeled by white letters with a black background.
The ORF of MjATP5L had 300 bps, encoding a putative protein of 99 amino acids (Figure 2A). MjATP5L protein was predicted to have a theoretical molecular mass of 10.47 kDa and a theoretical pI value of 9.30. The function domain in MjATP5L protein was ATP synthase subunit g (ATP-synt G) domain (Figure 2B). The phylogenetic tree showed that these ATP5Ls could be divided into two groups, with one group including insects, and the other group containing crustaceans. MjATP5L with ATP5Ls from other shrimp were located in the same subgroup of the crustacean group (Figure 2C). The similarity between MjATP5L and ATP5L from P. vannamei was around 97.0% (Figure 2D).
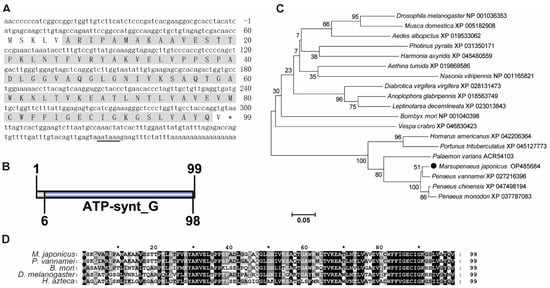
Figure 2.
Sequence analysis of MjATP5L. (A) The nucleotide and deduced amino acid sequence of MjATP5L. The letters with a grey background represent the ATP synthase subunit g domain. The double-underlined nucleotides show the polyadenylation signal region. (B) The domain schematics of MjATP5L. MjATP5L contained an ATP synthase G domain with 93 amino acids. (C) Phylogenetic tree analysis of ATP5Ls from Aedes albopictus, Aethina tumida, Anoplophora glabripennis, Bombyx mori, Diabrotica virgifera virgifera, Drosophila melanogaster, Harmonia axyridis, Homarus americanus, Leptinotarsa decemlineata, Marsupenaeus japonicus, Musca domestica, Nasonia vitripennis, Palaemon varians, Penaeus chinensis, Penaeus monodon, Penaeus vannamei, Photinus pyralis, Portunus trituberculatus, and Vespa crabro. ATP5L from M. japonicus is labeled by a black solid circle. All of the amino acid sequences were obtained from GenBank. (D) Multiple sequence alignment of ATP5L from M. japonicus, P. vannamei, B. mori, D. melanogaster, and Hyalella azteca. The conserved sites are labeled by white letters with a black background.
3.2. Tissue Distribution and Expression Profiles of MjATP5I and MjATP5L
To detect the tissue distribution and expression profiles of MjATP5I and MjATP5L, qPCR was performed. For the tissue distribution at the transcriptional level, MjATP5I could be detected in all six of the tested tissues including hemocytes, heart, hepatopancreas, gills, stomach, and intestines, with the highest expression in heart (Figure 3A). After WSSV challenge, the mRNA expression patterns of MjATP5I in the immune related tissues were checked by qPCR. The results show that the relative expression of MjATP5I could be upregulated at 36 h post viral immune challenge in hemocytes (Figure 3B) and intestines (Figure 3C).
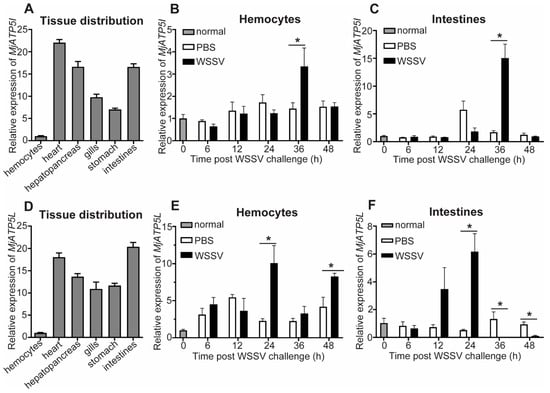
Figure 3.
Expression pattern analysis of MjATP5I and MjATP5L. (A) Tissue distribution of MjATP5I in tissues such as hemocytes, heart, hepatopancreas, gills, stomach, and intestines. (B,C) Expression profiles of MjATP5I in hemocytes (B) and intestines (C) post WSSV challenge. (D) Tissue distribution of MjATP5L in different tissues. (E,F) Expression profiles of MjATP5L in shrimp hemocytes (E) and intestines (F) after WSSV challenge. The PBS injected shrimp were used as the control. The results are expressed as the mean ± SD and significant differences were analyzed between the PBS-injected group and the WSSV-injected group using Student’s t-test. *, p < 0.05.
MjATP5L is also distributed in the six selected tissues with the highest expression level in the heart and intestines and the lowest expression level in the hemocytes (Figure 3D). The relative expression of MjATP5L in hemocytes could be upregulated significantly at 24 h and 48 h post WSSV infection (Figure 3E) and increased obviously in intestines at 24 h after the viral challenge (Figure 3F).
3.3. MjATP5I and MjATP5L Were Beneficial for WSSV Duplication
To check the involvement of MjATP5I and MjATP5L in WSSV duplication, qPCR was applied for the expression of viral genes after protein injection and RNAi. The predicted molecular masses of rMjATP5I or rMjATP5L were 27.44 kDa and 28.45 kDa, respectively, which were close to the molecular masses (around 30 kDa) in SDS-PAGE (Figure 4A,B). In the recombinant protein injection assay, the purified recombinant rMjATP5I or rMjATP5L was injected into the shrimp cavity after incubation with WSSV. The effects of recombinant protein injection were checked by immunocytochemistry assay. Considering the size of the tag protein, the purified Trx protein with a molecular mass of 20.40 kDa (Figure 4C), was used as the control to eliminate the effect of the tag protein. The results show that there were green fluorescence signals in the cytoplasm of the shrimp hemocytes post the injection of rMjATP5I or rMjATP5L (Figure 4D), while after the injection of Trx tag protein, the green fluorescence signals distributed in the whole hemocytes (Figure 4D), which was different from the distribution of rMjATP5I or rMjATP5L. It indicated that rMjATP5I and rMjATP5L could enter shrimp hemocytes and located in the cytoplasm to participate in viral immune response.
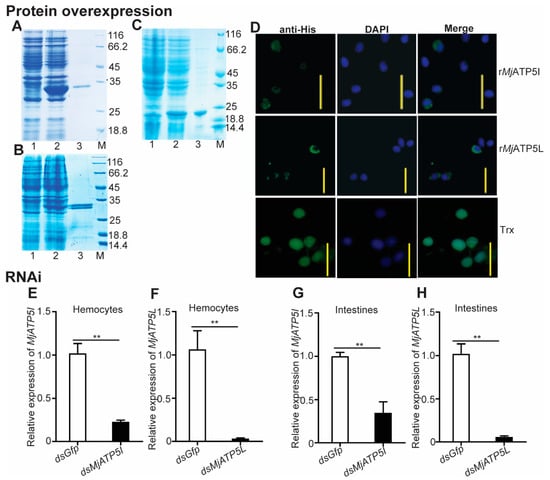
Figure 4.
Recombinant protein injection and RNAi effect detection for MjATP5I and MjATP5L in shrimp. The recombinant expression and protein purification of MjATP5I (A), MjATP5L (B), and Trx (C) in E. coli. Lane 1, total protein of E. coli with MjATP5I-pET32a, MjATP5L-pET32a, or empty pET-32a(+) vector without induction; lane 2, total protein of E. coli with MjATP5I-pET32a, MjATP5L-pET32a, or empty pET-32a(+) vector induced with 0.5 mM IPTG; lane 3, the purified rMjATP5I, rMjATP5L, or Trx; lane M, the standard protein marker. (D) Immunocytochemistry assay to detect the entrance of rMjATP5I, rMjATP5L, or Trx into shrimp hemocytes. The recombinant proteins in shrimp hemocytes were indicated by the green fluorescence signals, and the nuclei were stained by DPAI and shown by blue signals. Scale bar = 20 μm. (E,F) RNA interference effects of MjATP5I and MjATP5L were detected in hemocytes after RNAi. (G,H) Interference effects of MjATP5I and MjATP5L were detected in intestines after RNAi. The results are expressed as the mean ± SD and significant differences were analyzed between the dsMjATP5I-injected or dsMjATP5L-injected group and the dsGfp-injected group using Student’s t-test. **, p < 0.01.
Meanwhile, after knockdown of MjATP5I and MjATP5L in shrimp, WSSV was injected into shrimp. Firstly, the interference effect was checked by qPCR. The results show that the relative expression levels of MjATP5I and MjATP5L decreased obviously in both hemocytes (Figure 4E,F) and intestines (Figure 4G,H) compared to the control group. This demonstrates that MjATP5I and MjATP5L could be knocked down successfully in shrimp.
After the rMjATP5I injection, the expression of Ie1 and Vp28 increased obviously in hemocytes and intestines (Figure 5A–D), while after MjATP5I RNAi followed by viral injection, the relative expression of Ie1 and Vp28 decreased significantly in hemocytes and intestines (Figure 6A–D). Similar expression trends could be found in the rMjATP5L injection (Figure 5E–H) and dsMjATP5L injection assay (Figure 6E–H). All the results suggest that MjATP5I and MjATP5L are beneficial for the WSSV duplication in kuruma shrimp.
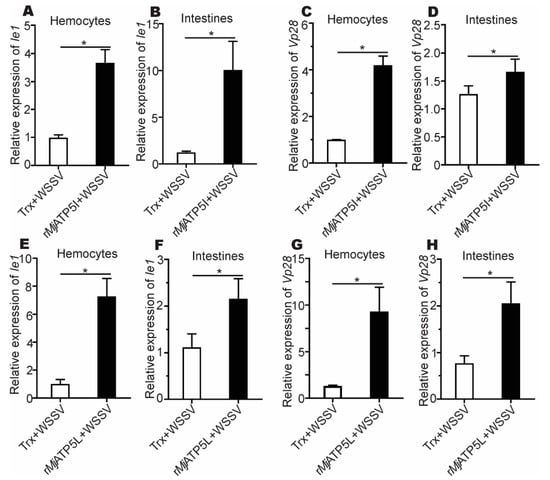
Figure 5.
The relative expression of Ie1 and Vp28 gene after recombinant protein injection. (A–D) The expression of Ie1 (A,B) and Vp28 (C,D) in hemocytes and intestines after rMjATP5I injection. (E–H) The expression of Ie1 (E,F) and Vp28 (G,H) in hemocytes and intestines after rMjATP5L injection. The results are expressed as the mean ± SD and significant differences were analyzed between rMjATP5I- or rMjATP5L-injected group and Trx-injected group using Student’s t-test. *, p < 0.05.
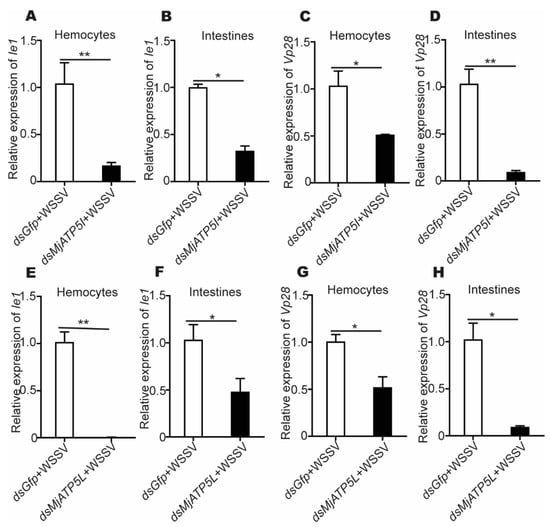
Figure 6.
The relative expression of Ie1 and Vp28 after RNAi. (A–D) The relative expression of Ie1 (A,B) and Vp28 (C,D) in hemocytes and intestines after MjATP5I knocking down. (E–H) The relative expression of Ie1 (E,F) and Vp28 (G,H) in hemocytes and intestines after MjATP5L knocking down. The results are expressed as the mean ± SD and significant differences were analyzed between the dsMjATP5I-injected or dsMjATP5L-injected group and the dsGfp-injected group using Student’s t-test. *, p < 0.05, **, p < 0.01.
3.4. MjATP5I and MjATP5L Influenced the Shrimp Survival Rate and the Production of ATP
To investigate the influence of the two ATP synthase subunits in shrimp survival, the survival rates were analyzed. The results showed that the survival rates of the dsMjATP5I-injected shrimp and the dsMjATP5L-injected shrimp were significantly higher than the control group post WSSV challenge (Figure 7A).
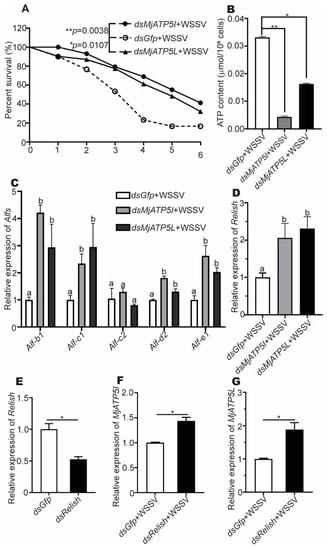
Figure 7.
The survival rate, ATP content detection, and the genes relative expression after RNAi and WSSV challenge. (A) Survival rates of the dsMjATP5I-injected and dsMjATP5L-injected shrimp compared with the dsGfp-injected shrimp after WSSV infection. The survival rate was calculated to present the survival curves as Kaplan–Meier plots. Differences between the MjATP5I RNAi group or MjATP5L RNAi group and the Gfp RNAi group were analyzed using the Log-rank test in the GraphPad Prism 8.0 program. (B) The content of ATP in hemocytes after MjATP5I and MjATP5L RNAi with WSSV challenge. The dsGfp-injected shrimp were used as the control. The results are expressed as the mean ± SD and significant differences were analyzed between the dsMjATP5I-injected or dsMjATP5L-injected group and the dsGfp-injected group using Student’s t-test. *, p < 0.05, **, p < 0.01. (C,D) Expression of Alfs (C) and Relish (D) in hemocytes after MjATP5I and MjATP5L knockdown. The columns are expressed as the mean ± SD, and the differences were calculated by the ordinary one-way ANOVA with multiple comparisons. The identical letters indicate there are no significant differences between the two groups for the same gene (p > 0.05), while different letters indicate significant differences between two groups of the same gene (p < 0.05). The dsGfp-injected shrimp were used as the control. (E) Expression of Relish in shrimp hemocytes after dsRelish injection. (F,G) The relative expression of MjATP5I (F) and MjATP5L (G) in hemocytes after Relish knockdown and WSSV infection. The results are expressed as the mean ± SD and significant differences were analyzed between the dsMjATP5I-injected or dsMjATP5L-injected group and the dsGfp-injected group using Student’s t-test. *, p < 0.05.
To explore the possible mechanism of the two ATP synthase subunits in the shrimp viral immune reaction, the ATP contents in shrimp hemocytes were measured after knockdown of MjATP5I and MjATP5L with WSSV challenge. We found that the content of ATP decreased dramatically after RNAi of MjATP5I and MjATP5L in hemocytes (Figure 7B). This revealed that MjATP5I and MjATP5L would affect the activity of ATP synthase and the subsequent ATP output of hemocytes in the shrimp defense reaction.
3.5. MjATP5I and MjATP5L Could Regulate the Expression of AMP Genes in Shrimp
To test whether MjATP5I and MjATP5L regulated the effector molecules of the immune signal pathway, the expression of AMP genes in the downstream of Relish was checked after RNAi and WSSV infection. Figure 7C shows that the expression of Alf-b1, Alf-c1, Alf-d2, and Alf-e1 was upregulated obviously in the MjATP5I and MjATP5L RNAi shrimp compared with the Gfp RNAi shrimp. However, there were no significant changes in the expression of Alf-c2. It was reported that both the Toll-Dorsal and the IMD-Relish pathway could regulate the expression of Alf-c2. So, in our assay, the expression of Alf-c2 might mainly be activated by the Toll-Dorsal pathway. The results demonstrate that some AMP genes under the control of Relish were regulated by MjATP5I and MjATP5L.
3.6. MjATP5I/MjATP5L and Relish Negatively Regulated Each Other
To investigate the relationship between MjATP5I/MjATP5L and Relish, the RNAi assay was performed. After the knockdown of MjATP5I and MjATP5L and WSSV infection, the relative expression of Relish increased significantly in hemocytes from the dsMjATP5I-injected or dsMjATP5L-injected shrimp compared with the dsGfp-injected shrimp (Figure 7D). The results imply that MjATP5I and MjATP5L could restrain the expression of Relish.
Meanwhile, after Relish RNAi (Figure 7E) and WSSV infection, the expression levels of MjATP5I and MjATP5L increased remarkably in hemocytes of the dsRelish-injected shrimp compared to the control (Figure 7F,G), which demonstrated that Relish repressed the expression of MjATP5I and MjATP5L.
Therefore, MjATP5I/MjATP5L and Relish negatively regulated each other’s expression in shrimp.
4. Discussion
In this study, we identified two ATP synthase subunits, ATP synthase subunit e and ATP synthase subunit g, from kuruma shrimp M. japonicus, and explored their possible functions in the viral immune reaction. To our knowledge, this is the first report about the function of the small ATP synthase subunits in shrimp viral immunity.
The two ATP synthase subunits were encoded by ATP5I and ATP5L, respectively, and defined as MjATP5I and MjATP5L in the present study. The amino acid sequences of MjATP5I and MjATP5L contained 85 and 99 residues, respectively. They were both newcomers to the sORF family. Recent studies showed that sORF functioned importantly in innate immunity as antimicrobial peptides, in the defense reaction against bacterial infection, in the development process, in the mitochondrial processes, and in the activity regulation of canonical proteins and so on [12,31,32]. Therefore, our identification and exploration of MjATP5I and MjATP5L in the viral immune reaction could expand the understanding for the novel functions of the sORF family.
The sequence analysis showed that ATP5I and ATP5L shared high similarities between kuruma shrimp and the other animals, mainly arthropods. It demonstrated that mitochondrial ATP synthase subunits were conservative in evolution. Additionally, from the results of the phylogenetic tree and the multiple sequence alignment, we found that MjATP5I and MjATP5L had close relationships with those from P. vannamei, which showed a close evolutionary relationship between kuruma shrimp and Pacific white shrimp.
In M. japonicus, MjATP5I and MjATP5L were widely distributed in all six of the examined tissues at the mRNA level. The relative expression of MjATP5I and MjATP5L could be induced in hemocytes and intestines after WSSV challenge. This indicated that they might participate in the viral immune response of kuruma shrimp. To detect whether shrimp ATP synthase subunits were involved in the viral immunity, we performed recombinant protein injection and RNAi assay, and analyzed the expression patterns of the viral genes after WSSV infection. The results show that the expression of Ie1 and Vp28 increased significantly after the injection of recombinant proteins (rMjATP5I and rMjATP5L), while the expression of Ie1 and Vp28 was downregulated after knockdown of MjATP5I and MjATP5L. These results indicate that MjATP5I and MjATP5L are beneficial for WSSV replication in shrimp.
ATP synthase is one important protein in the mitochondria. The function and mechanism of mitochondrial ATP synthase have already been studied very clearly. In addition to producing ATP for cellular survival, ATP synthase is also related to apoptosis. Pathogenetic variants of the ATP synthase subunit genes can cause fatal human diseases, such as neurodegenerative diseases and motor neuron diseases [33]. It is reported that angiostatin can bind and inhibit F1F0-ATP synthase to prevent tumor growth and metastasis [34]. Interestingly, besides locating in the inner membrane of the mitochondria, the subunits of ATP synthase were also found to locate in the eukaryotic cell surface and function as the receptor of the cell membrane [35]. Some larger ATP synthase subunits could interact with viruses. For example, the ATP synthase subunit beta was located on the cell surface of hemocytes and gill cells in Pacific white shrimp, P. vannamei [22]. It functioned importantly in the antiviral immune reaction to infection with yellow head virus and WSSV in shrimp [36,37,38,39]. In freshwater crayfish, Pacifastacus leniusculus, the ATP synthase subunit beta was identified on the plasma membrane of the hematopoietic tissue cells and served as the receptor for the cytokine astakine [35]. In addition to the large subunits, there are some small subunits in the ATP synthase. Most studies have shown that ATP5I and ATP5L are related to human diseases [21,25,27], and only a few studies reported their involvements in the growth or ammonia tolerance of invertebrates [26,29]. Their exact roles in the viral immunity of shrimp were explored in this study.
After knockdown of MjATP5I and MjATP5L in shrimp, the shrimp survival rates increased and the content of ATP in hemocytes decreased significantly compared to the control group. It was reported that WSSV would require and consume the host ATP for its replication and infection process [40]. Combined with the above results, we speculated that MjATP5I and MjATP5L were related to the ATP synthase activity, provided energy for WSSV replication and infection, and were used for the WSSV duplication and infection in shrimp. Therefore, the two small subunits of ATP synthase were beneficial for the WSSV duplication in shrimp. Knockdown of MjATP5I and MjATP5L significantly upregulated the expression of Relish and some antimicrobial peptides (Alf-b1, Alf-c1, Alf-d2, and Alf-e1). When the shrimp were challenged by WSSV, the expression of MjATP5I and MjATP5L was upregulated, the expression of Relish was inhibited, and then the expression of some downstream AMP genes decreased subsequently. Finally, the WSSV duplication was facilitated in shrimp. In a word, the ATP synthase subunits would be helpful for the WSSV duplication. The previous study showed that ATPase inhibitor factor 1 was beneficial for WSSV replication in kuruma shrimp M. japonicus through inducing the ROS production to activate the NF-κB signal pathway and regulating the expression of Dorsal [41]. Therefore, the mechanism of ATPase inhibitor factor 1 and the small subunits of ATP synthase in the viral immune reaction might be different.
In summary, two novel SEPs, MjATP5I and MjATP5L, were identified from kuruma shrimp M. japonicus. MjATP5I and MjATP5L can affect the activity of ATP synthase and the expression of the transcriptional factor Relish with the downstream AMP genes, and then facilitate the WSSV replication in shrimp.
Author Contributions
Conceptualization, X.-Z.S.; investigation, L.-J.H. and P.-Y.L.; analysis, L.-J.H., P.-Y.L. and X.-Z.S.; writing—original draft preparation, L.-J.H., P.-Y.L. and X.-Z.S.; writing—review and editing, X.-Z.S. and D.-X.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China, grant number 31930112, and the National Key Research and Development Program of China, grant number 2018YFD0900502.
Institutional Review Board Statement
The animal study protocol was approved by the Ethics Committee for Animal Research at Shandong University School of Life Sciences (protocol code SYDWLL-2022-054 and 22 March 2022).
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Flegel, T.W. Historic emergence, impact and current status of shrimp pathogens in Asia. J. Invertebr. Pathol. 2012, 110, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Hong, P.P.; Yang, M.C.; Zhao, X.F.; Wang, J.X. FOXO regulates the expression of antimicrobial peptides and promotes phagocytosis of hemocytes in shrimp antibacterial immunity. PLoS Pathog. 2021, 17, e1009479. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Wang, X.W.; Sun, J.J.; Wang, L.; Zhang, H.W.; Zhao, X.F.; Wang, J.X. Akirin interacts with Bap60 and 14-3-3 proteins to regulate the expression of antimicrobial peptides in the kuruma shrimp (Marsupenaeus japonicus). Dev. Comp. Immunol. 2016, 55, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.J.; Xu, S.; He, Z.H.; Shi, X.Z.; Zhao, X.F.; Wang, J.X. Activation of Toll pathway is different between kuruma shrimp and Drosophila. Front. Immunol. 2017, 8, 1151. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.S.; Lv, L.X.; Wang, J.X. Anti-lipopolysaccharide factor D from kuruma shrimp exhibits antiviral activity. Mar. Life Sci. Tech. 2022, 4, 52–61. [Google Scholar] [CrossRef]
- Li, S.; Guo, S.; Li, F.; Xiang, J. Functional diversity of anti-lipopolysaccharide factor isoforms in shrimp and their characters related to antiviral activity. Mar. Drugs 2015, 13, 2602–2616. [Google Scholar] [CrossRef]
- Aspden, J.L.; Eyre-Walker, Y.C.; Phillips, R.J.; Amin, U.; Mumtaz, M.A.; Brocard, M.; Couso, J.P. Extensive translation of small Open Reading Frames revealed by Poly-Ribo-Seq. Elife 2014, 3, e03528. [Google Scholar] [CrossRef]
- Saghatelian, A.; Couso, J.P. Discovery and characterization of smORF-encoded bioactive polypeptides. Nat. Chem. Biol. 2015, 11, 909–916. [Google Scholar] [CrossRef]
- Albuquerque, J.P.; Tobias-Santos, V.; Rodrigues, A.C.; Mury, F.B.; da Fonseca, R.N. small ORFs: A new class of essential genes for development. Genet. Mol. Biol. 2015, 38, 278–283. [Google Scholar] [CrossRef][Green Version]
- Couso, J.P. Finding smORFs: Getting closer. Genome Biol. 2015, 16, 189. [Google Scholar] [CrossRef]
- Srinivas, V.; Kumar, M.; Noronha, S.; Patankar, S. ORFpred: A Machine Learning Program to Identify Translatable Small Open Reading Frames in Intergenic Regions of the Plasmodium falciparum Genome. Curr. Bioinform. 2016, 11, 259–268. [Google Scholar] [CrossRef]
- Couso, J.P.; Patraquim, P. Classification and function of small open reading frames. Nat. Rev. Mol. Cell Biol. 2017, 18, 575–589. [Google Scholar] [CrossRef] [PubMed]
- Malekos, E.; Carpenter, S. Short open reading frame genes in innate immunity: From discovery to characterization. Trends Immunol. 2022, 43, 741–756. [Google Scholar] [CrossRef] [PubMed]
- Formentini, L.; Pereira, M.P.; Sanchez-Cenizo, L.; Santacatterina, F.; Lucas, J.J.; Navarro, C.; Martinez-Serrano, A.; Cuezva, J.M. In vivo inhibition of the mitochondrial H+-ATP synthase in neurons promotes metabolic preconditioning. EMBO J. 2014, 33, 762–778. [Google Scholar] [CrossRef]
- Havlickova, V.; Kaplanova, V.; Nuskova, H.; Drahota, Z.; Houstek, J. Knockdown of F1 epsilon subunit decreases mitochondrial content of ATP synthase and leads to accumulation of subunit c. Biochim. Biophys. Acta 2010, 1797, 1124–1129. [Google Scholar] [CrossRef]
- Bornhovd, C.; Vogel, F.; Neupert, W.; Reichert, A.S. Mitochondrial membrane potential is dependent on the oligomeric state of F1F0-ATP synthase supracomplexes. J. Biol. Chem. 2006, 281, 13990–13998. [Google Scholar] [CrossRef]
- Collinson, I.R.; Runswick, M.J.; Buchanan, S.K.; Fearnley, I.M.; Skehel, J.M.; van Raaij, M.J.; Griffiths, D.E.; Walker, J.E. Fo membrane domain of ATP synthase from bovine heart mitochondria: Purification, subunit composition, and reconstitution with F1-ATPase. Biochemistry 1994, 33, 7971–7978. [Google Scholar] [CrossRef]
- Leyva, J.A.; Bianchet, M.A.; Amzel, L.M. Understanding ATP synthesis: Structure and mechanism of the F1-ATPase (Review). Mol. Membr. Biol. 2009, 20, 27–33. [Google Scholar] [CrossRef]
- Cha, M.Y.; Cho, H.J.; Kim, C.; Jung, Y.O.; Kang, M.J.; Murray, M.E.; Hong, H.S.; Choi, Y.J.; Choi, H.; Kim, D.K.; et al. Mitochondrial ATP synthase activity is impaired by suppressed O-GlcNAcylation in Alzheimer’s disease. Hum. Mol. Genet. 2015, 24, 6492–6504. [Google Scholar] [CrossRef]
- Sardin, E.; Donadello, S.; di Rago, J.P.; Tetaud, E. Biochemical investigation of a human pathogenic mutation in the nuclear ATP5E gene using yeast as a model. Front. Genet. 2015, 6, 159. [Google Scholar] [CrossRef][Green Version]
- Mayr, J.A.; Havlickova, V.; Zimmermann, F.; Magler, I.; Kaplanova, V.; Jesina, P.; Pecinova, A.; Nuskova, H.; Koch, J.; Sperl, W.; et al. Mitochondrial ATP synthase deficiency due to a mutation in the ATP5E gene for the F1 epsilon subunit. Hum. Mol. Genet. 2010, 19, 3430–3439. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Cheng, J.J.; Yang, B.; Huang, J. The role of F1 ATP synthase beta subunit in WSSV infection in the shrimp, Litopenaeus vannamei. Virol. J. 2010, 7, 144. [Google Scholar] [CrossRef] [PubMed]
- Cui, C.; Zhu, L.; Tang, X.; Xing, J.; Sheng, X.; Chi, H.; Zhan, W. Differential white spot syndrome virus-binding proteins in two hemocyte subpopulations of Chinese shrimp (Fenneropenaeus chinensis). Dev. Comp. Immunol. 2021, 125, 104215. [Google Scholar] [CrossRef]
- Bustos, D.M.; Velours, J. The modification of the conserved GXXXG motif of the membrane-spanning segment of subunit g destabilizes the supramolecular species of yeast ATP synthase. J. Biol. Chem. 2005, 280, 29004–29010. [Google Scholar] [CrossRef] [PubMed]
- Ying, H.; Yu, Y.; Xu, Y. Antisense of ATP Synthase Subunit e Inhibits the Growth of Human Hepatocellular Carcinoma Cells. Oncol. Res. 2001, 12, 6. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Liu, Q.; Huang, Z.; Chen, X.; Yang, C.; Zhang, Y.; Zhao, Y.; Wang, F. ATP synthase subunit e is a shrimp growth-associated breeding marker. Genomics 2022, 114, 110410. [Google Scholar] [CrossRef]
- Dahiya, N.; Sarachana, T.; Kulkarni, S.; Wood, W.H., III; Zhang, Y.; Becker, K.G.; Wang, B.D.; Atreya, C.D. miR-570 interacts with mitochondrial ATPase subunit g (ATP5L) encoding mRNA in stored platelets. Platelets 2017, 28, 74–81. [Google Scholar] [CrossRef]
- Boyle, G.M.; Roucou, X.; Nagley, P.; Devenish, R.J.; Prescott, M. Identification of subunit g of yeast mitochondrial F1F0-ATP synthase, a protein required for maximal activity of cytochrome c oxidase. Eur. J. Biochem. 1999, 262, 315–323. [Google Scholar] [CrossRef]
- Zeng, D.; Yang, C.; Li, Q.; Zhu, W.; Chen, X.; Peng, M.; Chen, X.; Lin, Y.; Wang, H.; Liu, H.; et al. Identification of a quantitative trait loci (QTL) associated with ammonia tolerance in the Pacific white shrimp (Litopenaeus vannamei). BMC Genom. 2020, 21, 857. [Google Scholar] [CrossRef]
- Kumar, S.; Nei, M.; Dudley, J.; Tamura, K. MEGA: A biologist-centric software for evolutionary analysis of DNA and protein sequences. Brief. Bioinform. 2008, 9, 299–306. [Google Scholar] [CrossRef]
- Lu, Y.; Zhuang, Y.; Liu, J. Mining antimicrobial peptides from small open reading frames in Ciona intestinalis. J. Pept. Sci. 2014, 20, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Zanet, J.; Chanut-Delalande, H.; Plaza, S.; Payre, F. Small Peptides as Newcomers in the Control of Drosophila Development. Curr. Top. Dev. Biol. 2016, 117, 199–219. [Google Scholar] [PubMed]
- Garone, C.; Pietra, A.; Nesci, S. From the Structural and (Dys)Function of ATP Synthase to Deficiency in Age-Related Diseases. Life 2022, 12, 401. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Liang, H.; Liao, X.; Pan, J.; Chen, J.; Zhao, S.; Xu, Y.; Wu, Y.; Ni, J. A humanized chimeric antibody Hai178 targeted to the beta subunit of F1F0 ATP synthase. Tumour Biol. 2016, 37, 15903–15912. [Google Scholar] [CrossRef]
- Lin, X.; Kim, Y.A.; Lee, B.L.; Soderhall, K.; Soderhall, I. Identification and properties of a receptor for the invertebrate cytokine astakine, involved in hematopoiesis. Exp. Cell Res. 2009, 315, 1171–1180. [Google Scholar] [CrossRef]
- He, N.; Qin, Q.; Xu, X. Differential profile of genes expressed in hemocytes of White Spot Syndrome Virus-resistant shrimp (Penaeus japonicus) by combining suppression subtractive hybridization and differential hybridization. Antivir. Res. 2005, 66, 39–45. [Google Scholar] [CrossRef]
- Rosa, R.D.; Barracco, M.A. Shrimp interferon is rather a portion of the mitochondrial F0-ATP synthase than a true alpha-interferon. Mol. Immunol. 2008, 45, 3490–3493. [Google Scholar] [CrossRef]
- Wang, H.C.; Wang, H.C.; Leu, J.H.; Kou, G.H.; Wang, A.H.; Lo, C.F. Protein expression profiling of the shrimp cellular response to white spot syndrome virus infection. Dev. Comp. Immunol. 2007, 31, 672–686. [Google Scholar] [CrossRef]
- Bourchookarn, A.; Havanapan, P.O.; Thongboonkerd, V.; Krittanai, C. Proteomic analysis of altered proteins in lymphoid organ of yellow head virus infected Penaeus monodon. Biochim. Biophys. Acta 2008, 1784, 504–511. [Google Scholar] [CrossRef]
- Liang, Y.; Xu, M.L.; Wang, X.W.; Gao, X.X.; Cheng, J.J.; Li, C.; Huang, J. ATP synthesis is active on the cell surface of the shrimp Litopenaeus vannamei and is suppressed by WSSV infection. Virol. J. 2015, 12, 49. [Google Scholar] [CrossRef][Green Version]
- Huo, L.J.; Yang, M.C.; Wang, J.X.; Shi, X.Z. Mitochondrial ATPase inhibitor factor 1, MjATPIF1, is beneficial for WSSV replication in kuruma shrimp (Marsupenaeus japonicus). Fish Shellfish Immunol. 2020, 98, 245–254. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).